
苏云金芽孢杆菌Cry 毒素共性结构短肽串联表达及功能分析
2024-03-09 08:40金嘉凤徐重新刘贤金
河南农业科学 2024年1期
金嘉凤,沈 成,孟 萌,陈 蔚,徐重新,刘 媛,刘贤金
(1.江苏大学食品与生物工程学院,江苏 镇江 212013;2.江苏省农业科学院农产品质量安全与营养研究所∕省部共建国家重点实验室培育基地—江苏省食品质量安全重点实验室,江苏 南京 210014;3.南京农业大学植物保护学院,江苏 南京 210023;4.江苏大学生命科学学院,江苏 镇江 212013)
Bt Cry 毒素是由苏云金芽孢杆菌(Bacillus thuringiensis,Bt)代谢产生的一类对鳞翅目[1]、鞘翅目[2]等常见农业害虫具有特异性高致死活性的伴孢晶体蛋白;目前涉及Bt Cry 毒素对靶标害虫作用的机制研究,无论是“穿孔形成假说”还是“信号转导假说”,都共同指向Bt Cry毒素极有可能是通过抑制靶标害虫中肠受体如钙黏蛋白、腺苷三磷酸结合盒转运蛋白、氨肽酶、碱性磷酸酶等特殊功能蛋白或酶的活性,从而诱发一系列生理生化级联反应,造成中肠细胞受损,最终导致虫体死亡[3]。根据国际Bt 数据库权威网站(http:∕∕www.lifesci.sussex.ac.uk∕home∕Neil_Crickmore∕Bt∕)发布信息,截至2023年,已发现并命名的Bt Cry 毒素家族基因共有78 个(Cry1—Cry78),基因亚型共计818 个。Bt Cry 毒素是当今世界研究最深入、应用规模最大的生物抗虫蛋白,其制剂以及转基因抗虫作物在全球农业病虫害绿色防治上起到了引领作用,并带来了重要的经济价值与社会效益。根据国际农业生物技术应用服务组织(International Service for the Acquisition of Agri-biotech Applications,ISAAA)2019 年统计数据,目前全球推广种植的转Bt Cry毒素基因抗虫农作物年均超过1 亿hm2,农产品的价值增至约190 亿美元[4]。然而,随着Bt Cry 毒素制剂及转Bt Cry毒素基因抗虫作物长期大范围推广应用,由其诱发产生的靶标害虫非正常抗药性进化频率加剧以及严重威胁非靶标生物安全等问题日益凸显[5-6]。因此,探寻其抗虫功能效应物以及对其筛查与追踪检测用的创新技术意义重大,市场应用前景广阔。
在探寻Bt Cry 毒素功能效应物方面,早期较为传统的方式是以Bt Cry 毒素为模板,采用不同亚型的结构域[7]或功能片段[8]融合替换,实现具有更强抗虫活性或更广抗虫谱系的新型Bt Cry 毒素人工创制[9];而近年来,依托抗体“网络免疫学说”中Ab2β抗独特型抗体具有模拟抗原部分结构和生物功能的特性为理论基础[10],以Bt Cry 毒素抗体为靶标,从噬菌体展示抗体库中靶向筛选具备模拟Bt Cry毒素抗虫功能的Ab2β 抗独特型抗体[11],已成为创制Bt Cry 毒素功能效应物的全新路径,但在模拟功效上仍然未达到理想效果。在Bt Cry毒素筛查与追踪检测方面,基于抗体的免疫学检测方法是公认的最为简单快捷的方式[12]。目前,抗体创制研究从传统的多克隆抗体、单克隆抗体发展到了全新的基因工程抗体创制与应用阶段[13],极大地推进了Bt Cry 毒素检测方法的创新发展,但还需进一步改进和提升基因工程抗体的稳定性。
笔者团队前期在Bt Cry毒素共性结构和功能分析研究中,通过比较7 种Bt Cry 毒素(Cry1Aa、Cry1Ab、Cry1Ac、Cry1B、Cry1C、Cry1E、Cry1F)的氨基酸序列信息,获得了3 个初步具备模拟Bt Cry 毒素部分结构和功能的共性结构短肽[14],氨基酸序列分别为:CSQRYRVRIRYAS、CSFREWEADPTNPALREEMRI 和CSGTVDSLDEIPPQNNNV;但因这3 个短肽分子质量太小,既难以通过常规方式进行克隆表达与纯化制备,也无法将其作为免疫原直接用于免疫动物制备Bt Cry 毒素广谱抗体。基于此,拟借鉴成熟的商品化人源噬菌体展示单链抗体库重-轻链拼接技术[15],通过设计特异性Linker 将这3 个共性结构短肽串联拼接成为具备免疫原功能的大分子多肽,以期得到更具研究意义和应用价值的可模拟Bt Cry毒素抗虫功能以及替代其作为免疫原用于免疫制备Bt Cry毒素广谱抗体的新型多肽材料。
1 材料和方法
1.1 材料
供试Bt Cry 毒素共性结构串联多肽(命名为Bt Cry-GXJG-11)氨基酸序列如图1 所示,其对应核苷酸序列委托通用生物(安徽)股份有限公司合成;pSUMO-Mut 表达载体、E.coliBL21 感受态细胞购于北京全式金生物技术有限公司;Bt Cry 毒素(Cry1Ab、Cry1Ac、Cry1Ah、Cry1B、Cry1C、Cry1F)购于美延(北京)农业科技有限责任公司;供试靶标害虫棉铃虫3 龄幼虫以及棉铃虫钙黏蛋白受体(HaCad-TBR)[16]由省部共建国家重点实验室—江苏省食品质量安全重点实验室饲养和制备提供;供试Balb∕c模式试验小白鼠由扬州大学比较医学中心提供,动物免疫流程符合省部共建国家重点实验室—江苏省食品质量安全重点实验室理论委员会要求。试验试剂:EcoRⅤ∕XhoⅠ核酸内切酶、T4 DNA连接酶、PCR 纯化试剂盒、质粒提取试剂盒、卡那霉素(Kana)、IPTG 诱导剂购于日本Takara 公司;Anti His-Tag(HRP)鼠源单克隆抗体、Goat anti-mouse IgG-HRP 单克隆抗体、Biosharp BL741A 预染蛋白Marker 购于北京擎科生物科技有限公司;TMB 底物显色液、ECL 显色液、PVDF 膜购于北京普利莱基因技术有限公司;其他试剂均为分析纯。

图1 Bt Cry-GXJG-11基因对应的氨基酸序列Fig.1 The amino acid sequence of Bt Cry-GXJG-11 gene
1.2 方法
1.2.1 Bt Cry-GXJG-11 同源建模、模型评价及分子对接 参照GROCHULSKI等[17]和ZHAO等[18]报道方法,将供试Bt Cry-GXJG-11氨基酸序列作为目标蛋白,以同源性较高的Cry1Ac毒素晶体结构作为同源建模模板,利用Swiss-Model(https:∕∕swissmodel.expasy.org∕)预测该共性结构串联多肽的三维模型。进而采用在线网站https:∕∕saves.mbi.ucla.edu∕中的ERRAT、Verify3D 以及WHATCHECK 模块对其进行综合评估,分析其合理性。将Bt Cry-GXJG-11 和Cry1Ac 毒素与靶标害虫棉铃虫HaCad-TBR 受体蛋白的pdb 文件利用ZDOCK 在线网站(https:∕∕zdock.umassmed.edu∕)进行对接,得到10个对接复合物,进一步利用DockScore 获得综合打分最高的复合物。通过DrugscorePPI在线网站提交综合排序最高的对接复合物pdb 文件,计算突变前后结合自由能的变化值。
1.2.2 Bt Cry-GXJG-11 基因克隆 以合成的Bt Cry-GXJG-11 基因为模板,设计特异性上游引物序列“TGAGGCTCACCGCGAACAGATTGGAGGCTGCAGTCAGCGCTATCGTGTTCGTATTCGTTATGCAA”和特异性下游引物序列“TTTGTTAGCAGCCGGATCTCAGGTGCTCGAGTTAAACATTATTGTTCTGCGGCGGGATTTCATCC”对其目的基因片段进行扩增。PCR 扩增条件:首先94 ℃预变性10 min,接着94 ℃变性1 min、56 ℃退火1 min、72 ℃延伸1 min,共30个循环反应,最后72 ℃延伸10 min。以琼脂糖凝胶电泳分析PCR 产物的目的条带状况,最后借助纯化试剂盒纯化目的片段。
取纯化后的Bt Cry-GXJG-11 基因片段和pSUMO-Mut 载体分别以EcoRⅤ和XhoⅠ核酸内切酶进行双酶切,借助PCR 产物纯化试剂盒纯化回收相应酶切产物,并且酶连过夜。次日以42 ℃热激90 s 将酶连体系转化到E.coliBL21 感受态细胞中,接着涂布于含50µg∕mL Kana 的LB 固体培养基上,37 ℃过夜培养。从培养板中随机挑取长出的单克隆菌落,接种到含有Kana的LB液体培养基中,Kana终质量浓度为50µg∕mL,在37 ℃摇床中以220 r∕min振荡培养过夜。次日取培养菌液分别采用T7 通用引物对目的基因进行PCR 扩增、重组质粒双酶切以及目的片段测序等方式进行综合鉴定。
1.2.3 Bt Cry-GXJG-11 蛋白原核表达与纯化 挑取经鉴定为Bt Cry-GXJG-11 基因克隆阳性的单克隆菌液接种到含Kana 抗生素的LB 液体培养基中,在37 ℃摇床中以220 r∕min 振荡培养过夜。次日将培养菌液按1∶1 000 转接到含Kana 的LB 液体培养基中,在37 ℃摇床中以220 r∕min 振荡培养2.5 h 至对数生长期,接着向体系中加入终浓度为0.2 mmol∕L IPTG诱导剂,随后将温度调至15 ℃,诱导表达16 h。提前预冷离心机至4 ℃,取出诱导培养体系以8 000×g离心10 min,弃上清液后,沉淀细胞以30 mL PBS 重悬。取出重悬体系,借助超声波细胞破碎仪破碎细胞30 min,频率为每工作1 s、间隙2.5 s。事先将离心机预冷到4 ℃,裂解液以8 000×g离心20 min,收集上清液(即含Bt Cry-GXJG-11 目的蛋白),借助12%SDS-PAGE 电泳分离后,采用考马斯亮蓝法对目的蛋白进行染色分析。
然后利用His-Trap(HP)亲和纯化柱对上清液中的目的蛋白进行梯度洗脱。先用含有10 mmol∕L咪唑溶液25 mL 平衡Ni 柱,使之带有电荷,再封住Ni 柱底端。将上清溶液上样至Ni-IDA Binding-Buffer 预平衡的亲和层析柱,反复上样3 次。用5 倍柱体积的20 mmol∕L 咪唑冲洗,洗去未与柱子结合的蛋白质。再用含有250 mmol∕L 咪唑的Elution-Buffer 洗涤柱子,收集洗脱液加入透析袋中,使用PBS进行透析过夜,收集不含有咪唑的蛋白质溶液。最终将获得的目的蛋白采用Western-Blot 法进行鉴定和分析。
1.2.4 Bt Cry-GXJG-11 蛋白竞争ELISA 和分子互作分析 竞争ELISA分析:将10µg∕mL的Cry1Ac毒素蛋白按100 µL∕孔包被到96 孔酶标板中,置于4 ℃静置包被过夜。次日取出包被板,用200µL∕孔PBST 缓冲液(PBS 缓冲液中含0.05% Tween 20)洗板3 次,接着用200µL∕孔MPBS 封闭液(含4%脱脂奶粉的PBS)在37 ℃恒温培养箱中封闭2 h,然后再以PBST 缓冲液洗涤。随即按梯度顺序,每孔加入不同体积10 µg∕mL HaCad-TBR 受体蛋白溶液(0、100、150、175、190、195、199、200µL)与不同体积10µg∕mL 的Bt Cry-GXJG-11 蛋白溶液(200、100、50、25、10、5、1、0µL)预混复合液,置于37 ℃恒温培养箱中孵育1 h,取出孵育板,以PBST 缓冲液洗涤后,每孔加入以PBS 溶液按1∶5 000 稀释的商品化Anti His-Tag(HRP)鼠源单克隆抗体100 µL,并置于37 ℃恒温培养箱内再次进行1 h孵育。取出孵育板,以PBST 缓冲液洗涤后,每孔加入100 µL 显色液,在暗光条件下反应15 min 至显色,最后每孔加入50 µL 终止液(2 mol∕L H2SO4),在酶标板中测定各孔的OD450值,绘制竞争抑制率曲线。
分子互作分析:将纯化出的蛋白质样品与HaCad-TBR 进行分子互作,通过传感器芯片实时检测整个过程的相互作用。选用APS 探针,在PBS 中浸泡10 min。分别配制0.1%和10%的BSA溶液,将PBS 溶液、BSA 溶液、HaCad 受体和蛋白质样品按每孔220 µL 包被96 孔板(包被顺序为PBS、HaCad、0.1% BSA、Bt Cry-GXJG-11 蛋白∕Cry1Ac、0.1%BSA)。Qctet 运行软件选择参数Baseline、Loading、Baseline 2、Association、Disassociation,运行后通过分析软件获取生物分子结合、解离常数等,验证是否结合。
1.2.5 Bt Cry-GXJG-11 蛋白抗虫活性测定 取1 mL 质量浓度为10µg∕mL 的Bt Cry-GXJG-11 蛋白溶液、1 mL PBS(阴性对照)、1 mL 质量浓度为10µg∕mL 的Cry1Ac 蛋白溶液(阳性对照)均匀涂布到含有固体饲料的培养皿上,置于室温晾干后,每个培养皿挑入20 头处于3 龄幼虫阶段的棉铃虫,在室温条件下进行室内抗虫活性测定试验。每24 h记录1 次培养皿中供试虫的生长发育状态,一共记录120 h,每处理一组试验反复进行3 次。借鉴仲建锋等[19]报道的方法,对本试验的检定虫死率和误差进行分析。
1.2.6 Bt Cry-GXJG-11 蛋白免疫效果测定 首先,将100 µL 质量浓度为100 µg∕mL 的Bt Cry-GXJG-11蛋白溶液与150µL完全弗氏佐剂混匀并乳化,采用腹部注射的方式,将其注射到6 周龄的Balb∕c 模式试验小白鼠体内。饲养2 周后进行第2 次免疫,将100 µL 质量浓度为100 µg∕mL 的Bt Cry-GXJG-11 蛋白溶液与150µL 不完全弗氏佐剂混匀并进行充分乳化后,同样采用腹部注射方式注射到供试小鼠体内,2周后从供试小鼠尾部抽取血液(约50µL)用ELISA 法测定免疫效果;同时按第2 次免疫方式进行第3 次免疫,且在2 周后抽取供试小鼠尾部血液用ELISA法测定免疫效果。
参照上述竞争ELISA 法描述步骤,分别将质量浓度为10 µg∕mL 的Cry1Ab、Cry1Ac、Cry1Ah、Cry1B、Cry1C、Cry1F 毒素蛋白按100 µL∕孔包被到96 孔酶标板中,经过“洗涤—封闭—洗涤”步骤,将采集获得的供试小鼠血清梯度稀释液(以PBS 溶液按10 倍稀释法进行稀释)以100 µL∕孔加入包被板中,经过“孵育—洗涤”过程,每孔加入100 µL 以PBS 溶液按1∶5 000 稀释的商品化Goat anti-mouse IgG-HRP 单克隆抗体,再经过“孵育—洗涤—显色—终止”过程,测定各孔的OD450值,绘制ELISA 反应曲线。
2 结果与分析
2.1 Bt Cry-GXJG-11同源建模、模型评价及分子对接结果
采用Swiss-Model 预测Bt Cry-GXJG-11 的三维结构(图2),其主体构象为α 螺旋;对其进行模型评价(图3)发现,拉氏图允许区参数达到96.9%,无氨基酸落在不合理区,说明构建的模型可信度较高。以Cry1Ac 毒素为对照标准,分别将Bt Cry-GXJG-11和Cry1Ac毒素与靶标害虫棉铃虫HaCad-TBR 受体进行对接,它们在DockScore 软件上获得的综合打分最高的复合物构象分别如图4 和图5 所示。通过DrugscorePPI在线网站提交这2 种对接复合物pdb文件,计算出相应复合物结合自由能的变化值:其中Cry 1Ac 毒素与HaCad-TBR 互作关键位点存在3个氨基酸残基的结合自由能变化值>2 kJ∕mol(图6),而Bt Cry-GXJG-11 与HaCad-TBR 互作关键位点(244TYR)也存在1 个氨基酸残基的结合自由能的变化值>2 kJ∕mol(图7),符合蛋白质互作高结合活性的特征。通过对比Cry1Ac 毒素(图6)和Bt Cry-GXJG-11(图7)与HaCad-TBR 的结合自由能参数信息,发现供试Bt Cry-GXJG-11在三维结构上具备模拟Bt Cry 毒素某些特征性构象,从而与Bt Cry毒素靶标害虫棉铃虫HaCad-TBR 关键受体产生了类似Cry1Ac毒素的互作反应。

图2 Bt Cry-GXJG-11三维结构模型Fig.2 Three-dimensional structure of Bt Cry-GXJG-11

图3 Bt Cry-GXJG-11三维结构模型的拉氏图分析Fig.3 Ramachandran plot of Bt Cry-GXJG-11 threedimensional structure

图4 Bt Cry-GXJG-11与HaCad-TBR分子对接复合物构象Fig.4 Conformation of Bt Cry-GXJG-11 docking complex with HaCad-TBR molecule

图5 Cry 1Ac毒素与HaCad-TBR分子对接复合物构象Fig.5 Conformation of Cry 1Ac toxin docking complex with HaCad-TBR molecule

图6 Cry1Ac中主要参与结合的各残基结合自由能变化Fig.6 Change of binding free energy of each residue in Cry1Ac
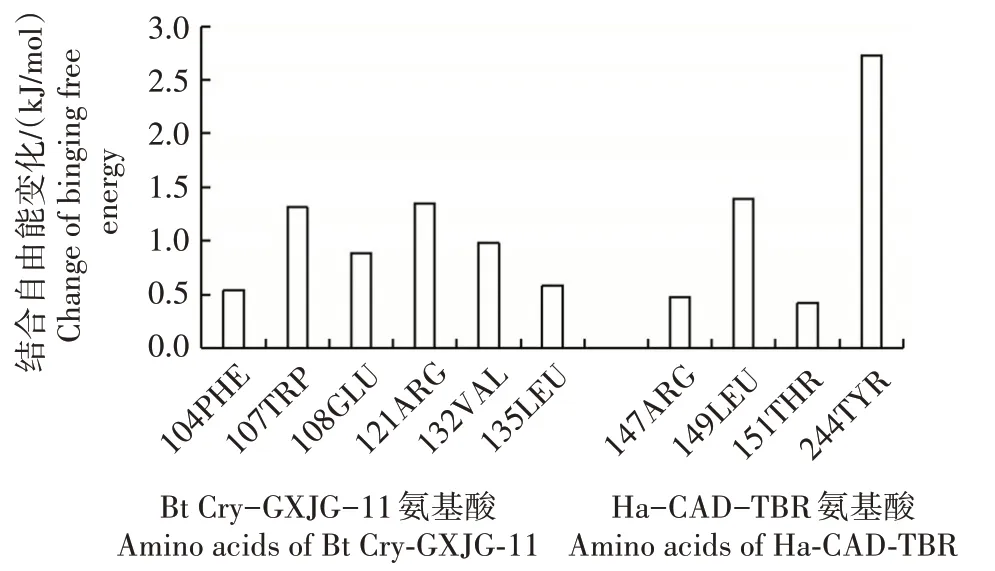
图7 Bt Cry-GXJG-11中主要参与结合的各残基结合自由能变化Fig.7 Change of binding free energy of each residue in Bt Cry-GXJG-11
2.2 Bt Cry-GXJG-11基因克隆结果
Bt Cry-GXJG-11 基因在特异性引物PCR 扩增后,产物经琼脂糖凝胶电泳,在450 bp 左右出现了与预期目的DNA 分子质量大小相当的条带(图8),说明PCR 扩增成功。经PCR 产物回收试剂盒纯化、酶切、酶连等步骤,将构建的pSUMO-Mut-Bt Cry-GXJG-11 重组质粒载体转化到E.coliBL21 感受态细胞中,在抗性筛选培养基上涂布长出的阳性单菌落,经T7 通用引物PCR 扩增,其在琼脂糖凝胶上出现了与预期目的DNA 分子质量大小相当的DNA 条带(图9);同时以阳性克隆菌液提取的质粒经EcoRⅤ-XhoⅠ核酸内切酶特异性双酶切鉴定,释放出了与预期分子质量大小相当的DNA 条带(图10)。结合氨基酸序列结果(图1),充分证实pSUMO-Mut-Bt Cry-GXJG-11 重组质粒构建成功,可用于后期目的蛋白诱导表达。

图8 Bt Cry-GXJG-11基因PCR产物Fig.8 PCR product of Bt Cry-GXJG-11

图9 pSUMO-Mut-Bt Cry-GXJG-11重组质粒转化E.coli BL21感受态细胞后的菌落PCR产物Fig.9 Colony PCR products of pSUMO-Mut-Bt Cry-GXJG-11 recombinant plasmids in E.coli BL21

图10 pSUMO-Mut-Bt Cry-GXJG-11重组质粒双酶切验证Fig.10 Double digestion of pSUMO-Mut-Bt Cry-GXJG-11 recombinant plasmids
2.3 Bt Cry-GXJG-11蛋白原核表达与纯化结果
将经鉴定为阳性的pSUMO-Mut-Bt Cry-GXJG-11重组质粒菌液接种到含Kana的LB液体培养基中,在15 ℃和220 r∕min 培养条件下,以终浓度为0.2 mmol∕L IPTG 诱导表达16 h,该诱导表达上清液经His-Trap(HP)亲和纯化柱梯度洗脱和纯化回收,并用PBS 进行透析过夜,结合SDS-PAGE 电泳(图11)分析结果,确定制备获得了Bt Cry-GXJG-11蛋白,其在洗脱缓冲液中的质量浓度为0.5 mg∕mL。

图11 Bt Cry-GXJG-11蛋白表达及纯化Fig.11 Expression and purification of Bt Cry-GXJG-11 protein
2.4 Bt Cry-GXJG-11蛋白竞争ELISA和分子互作分析结果
设计将Cry1Ac毒素作为包被原,然后以制备的系列梯度浓度Bt Cry-GXJG-11 蛋白作为竞争抑制物,抑制靶标害虫棉铃虫HaCad-TBR 受体蛋白与Cry1Ac 毒素结合,建立竞争抑制ELISA。结果如图12 所示,Bt Cry-GXJG-11 蛋白能与Cry1Ac 毒素靶标害虫棉铃虫HaCad-TBR 受体蛋白产生特异性结合反应,从而抑制其与Cry1Ac 毒素结合,说明Bt Cry-GXJG-11蛋白在空间结构上与Bt Cry毒素具有某些相似之处。与此同时,借助Octet蛋白质互作分析仪,通过生物膜干涉法测定了Bt Cry-GXJG-11蛋白与靶标害虫棉铃虫HaCad-TBR 受体的结合能力以及互作的动力学过程(图13)。结果发现,Bt Cry-GXJG-11 蛋白与棉铃虫HaCad-TBR 受体蛋白具有良好结合能力,其亲和力常数(KD)值达到2.151×10-7mol∕L,这也进一步证实Bt Cry-GXJG-11 蛋白具备模拟Bt Cry毒素某些结构的功能。

图12 Bt Cry-GXJG-11蛋白与棉铃虫HaCad-TBR受体的竞争抑制ELISA曲线Fig.12 Competitive inhibition ELISA curve of Bt Cry-GXJG-11 for HaCad-TBR
2.5 Bt Cry-GXJG-11蛋白抗虫活性测定结果和免疫效果
以质量浓度为10 µg∕mL 的Bt Cry-GXJG-11 蛋白溶液作为测试药剂,供试棉铃虫幼虫在取食浸有该药剂的饲料后,结果如表1 所示,120 h 后校正死亡率达到27.5%,而同样质量浓度Cry1Ac 毒素阳性对照的校正死亡率为91.2%,Bt Cry-GXJG-11 蛋白对棉铃虫致死率接近Cry1Ac 毒素的30%。而以Bt Cry-GXJG-11 蛋白溶液为免疫原,经3 次免疫过后,采集的供试小鼠血清对Cry1Ab、Cry1Ac、Cry1Ah、Cry1B、Cry1C 和Cry1F 等6 种供试Bt Cry 毒素具有较强的广谱识别能力,其中经第3 次免疫后对Cry1Ah 的结合能力最强,效价达到1.5×103,而对Cry1B 结合能力相对较弱(图14、15)。由此充分证实,设计的Bt Cry-GXJG-11在一定程度上具有模拟Bt Cry毒素结构和抗虫活性的功能。

表1 Bt Cry-GXJG-11蛋白对棉铃虫室内杀虫活性Tab.1 The insecticidal activity of Bt Cry-GXJG-11 protein against Helicoverpa armigera indoor

图14 Bt Cry-GXJG-11蛋白第2次免疫小鼠血清对供试Bt Cry毒素的ELISA曲线Fig.14 The Second time immune effect of Bt Cry-GXJG-11 using mice analyzed by ELISA with Bt cry toxins

图15 Bt Cry-GXJG-11蛋白第3次免疫小鼠血清对供试Bt Cry 毒素的ELISA曲线Fig.15 The third time immune effect of Bt Cry-GXJG-11using mice analyzed by ELISA with Bt cry toxins
3 结论与讨论
尽管Bt Cry 毒素亚型很多,但它们在进化上高度保守,分子质量接近,结构较为类似,抗虫谱系也大同小异[20],这为Bt Cry 毒素共性结构设计提供了可能。本研究基于团队前期依托7 种较为典型的Bt Cry 毒素氨基酸序列特征获得的分别对应Bt Cry毒素Domian I(CSQRYRVRIRYAS)、Domian Ⅱ(CSFREWEADPTNPALREEMRI) 和 Domian Ⅲ(CSGTVDSLDEIPPQNNNV)的3段共性结构短肽[14],通过设计特异性Linker将它们拼接复合成为具有免疫原性的长链多肽(Bt Cry-GXJG-11),其结构与上述任何一个短肽共性结构相比,在理论上拥有更大程度Bt Cry 毒素特征的共性结构,因此在模拟Bt Cry 毒素结构和功能上具备潜在的更大可能性和优势,这在其与Bt Cry 毒素靶标害虫棉铃虫HaCad-TBR 受体蛋白同源建模和分子对接预测的信息中也得到了初步印证。本研究依托pSUMO-Mut 质粒载体及其配套的E.coliBL21表达菌,从表达上清液中制备获得了质量浓度为0.5 mg∕mL 的Bt Cry-GXJG-11 蛋白,换算到最初的原始培养体系其质量浓度约为1.5 mg∕L,这个表达水平与MORICOLI等[21]、QIU 等[22]和RAHBARNIA 等[23]报道的分子质量与其相当的目的蛋白在各自构建的原核表达体系中分别能达到35、40、133 mg∕L 的结果相比还有较大差距,因此,在后期研究中有必要进一步优化包括表达载体及其配套宿主菌以及培养温度、诱导剂浓度、培养时间等条件来提升Bt Cry-GXJG-11蛋白表达水平和制备效率。目前,纯化制备的Bt Cry-GXJG-11 蛋白对靶标害虫棉铃虫的致死率为27.5%,抗虫活性只达到Cry1Ac毒素的30%,显然这个水平远达不到实际应用的要求,但在同源建模与分子对接预测中初步锁定了Cry-GXJG-11 与靶标害虫棉铃虫HaCad-TBR 受体互作的关键位点,这为后期对其活性改造指明了方向。值得注意的是,以同样纯化制备的Bt Cry-GXJG-11蛋白为免疫抗原,经过3 次免疫刺激,供试小鼠血清就能同时识别供试Cry1Ab、Cry1Ac、Cry1Ah、Cry1B、Cry1C 和Cry1F等6 种Bt Cry 毒素,说明该多肽材料较为完整地保留了Bt Cry 毒素特征的共性结构,其在作为免疫原制备Bt Cry毒素检测用广谱抗体上较前期研究中须将各短肽共性结构分别偶联载体蛋白才能作为免疫原[14]的策略更为方便和高效,也有望直接作为包被抗原从噬菌体展示基因工程抗体库中淘筛获得Bt Cry 毒素广谱检测用基因工程抗体,这显然比XU等[24]设计的采用不同Bt Cry 毒素作为交替包被抗原的烦琐过程才能从供试噬菌体抗体库中捕获靶标基因工程抗体的策略更为便捷高效。
综上表明,本研究创制的具备模拟Bt Cry 毒素共性结构和功能的多肽已经展示出了作为免疫原用于高效制备Bt Cry毒素检测用广谱抗体的良好应用价值,同时在替代Bt Cry 毒素抗虫蛋白用于靶标农业害虫绿色防治上也具有进一步深入研究和挖掘应用的潜力。
猜你喜欢
中国棉花(2023年4期)2023-08-08
军民两用技术与产品(2021年10期)2021-03-16
林业科技(2020年3期)2021-01-21
世界农药(2019年3期)2019-09-10
中国蔬菜(2016年8期)2017-01-15
光学精密工程(2016年4期)2016-11-07
新疆农垦科技(2016年2期)2016-08-21
现代农业(2016年5期)2016-02-28
现代农业(2016年4期)2016-02-28
肿瘤影像学(2015年3期)2015-12-09
