
甘南高寒草甸退化对植物功能群物种多样性与物种多度分布的影响
2024-03-08 12:33:00刘旻霞杨春亮
西北植物学报 2024年1期
刘旻霞,刘 成,杨春亮
(西北师范大学 地理与环境科学学院,兰州 730070)
物种多度分布是群落生态学的重要研究内容,研究群落物种多度分布有助于揭示群落特征,反映群落构建过程[1]。物种-多度关系最早由Motomura[2]提出。物种多度分布模型可以分为统计模型、生态位模型和中性理论模型:统计模型基于数学统计,生态学效应较弱,多数学者更关注基于生态位理论的生态位模型和基于中性理论的中性模型。生态位理论认为物种生态位的分化是物种共存的依据,而中性理论认为随机效应和扩散限制因素是控制物种分布的主导因素,这一理论也为传统生态位观点的不足提供了较为完善的补充说明[3]。物种多度分布的变化与时间动态密切相关,随时间行进,曾经占据主导作用的生态过程可能不再重要,群落物种多度分布发生变化。不同时间阶段的群落具有不同的物种多度分布特征,了解群落不同阶段物种组成、群落结构和物种多度分布特征,对于理解维持物种共存的生态学过程具有重要意义[4]。生态学家将植物功能群定义为对相似环境条件发生类似响应的植物类群[5]。植物功能群忽略了物种之间的差异,更加强调植物的功能属性,例如生长形态、生命周期、光合作用方式等,从而更加聚焦于生态系统的功能和生态过程。通过对群落进行功能群分类,可以简化研究系统的复杂程度,这在研究生态系统的功能、结构以及动态变化时是非常必要的[6]。青藏高原是物种形成和分化的中心,是重要的生态屏障,也是对全球气候变化最敏感的地区之一[7]。
近年来,由于人类活动和气候变化的影响,青藏高原出现了大面积的高寒草甸退化现象,高寒草甸的退化导致了青藏高原生态系统功能的恶化,严重威胁到青藏高原及其下游地区的生态安全和可持续发展[8]。
为了探究退化高寒草甸的生态质量,研究草甸退化机理和退化后的治理措施,多年来不少学者从不同层面进行了大量研究:随着退化持续发生,高寒草甸生产力不断下降,植被群落的盖度、地上和地下生物量下降,群落结构的稳定性和群落物种的多样性发生变化,组分以禾本科为主向以杂类草类为主转变[9]。在此背景下,以退化过程中的高寒草甸植物群落为研究对象,通过对群落物种多度分布进行模型拟合,回答以下问题:植物功能群多样性随退化程度的加深是如何变化的? 群落全部物种多度分布格局随退化程度的加深是如何变化的? 最优拟合模型是什么? 不同功能群植物物种多度分布格局有何不同? 对以上问题进行探究,可以丰富不同退化程度下植物群落物种多度分布格局的相关研究,也有助于揭示功能群多样性的分布规律以及形成机制,展现该地区群落形成中的资源分配模式,从而为青藏高原高寒草甸的植被恢复和生物多样性的保护提供技术支撑和理论依据。
1 材料和方法
1.1 研究区概况
研究区位于甘肃省甘南藏族自治州玛曲县的高寒草甸与湿地生态系统科学观测研究站(33°06′—36°10′N,100°46′—104°44′E)。海拔平均在3 000 m 以上,气候类型为寒冷湿润的高原气候,冬季寒冷漫长,夏季温暖短促,平均气温在2.8~13.6℃之间。年平均降水量在500~750 mm 之间,降水集中于6—8月,雨热同期,日照充足,最高日照时间可达2 600 h,平均无霜期可达48 d。植被组成以多年生草本植物为主,主要优势种有:矮生嵩草(Carex alatauensis)、垂穗披碱草(Clinelymus nutans)、早熟禾(Poa annua)和鹅绒委陵菜(Potentilla anserina)等。
1.2 样地建立与群落调查
样地调查为2021年7月。草甸退化是一个长时序的过程,采用空间序列来取代时间序列。参考Wen等[9]建立的草地退化指数(grassland degradation index,GDI),将草甸分为无退化草甸(non-degraded meadow,ND)、轻度退化草甸(lightly degraded meadow,LD)、中度退化草甸(moderately degraded meadow,MD)和重度退化草甸(severely degraded meadow,SD)。其中,GDI计算公式为:
式中:C为植被盖度;P为产草量比例;E为可食用牧草比例;H为可食用牧草高度。当GDI指数小于13.9时,表示该样地为未退化草甸;GDI指数在13.9~33.5之间时,表示该样地为轻度退化草甸;GDI指数为33.5~52.4时,该样地为中度退化草甸;GDI指数在52.4~68.4之间时,该样地为重度退化草甸。
不同退化程度的山坡要求坡度相近,坡向相同,按照4个退化程度设置样地,每个退化程度随机设置5个10 m×10 m 的大样方,每个大样方内按五点法设置5个1 m×1 m 的小样方,总计100个小样方。记录样地的坡度、坡向和经纬度等数据,并测量地表裸露程度,实测并记录植物物种及其数量,植被高度采用样方内植物自然高度的平均值(使用直尺进行10次测量,而后计算其平均值),用针刺法测量植被盖度。
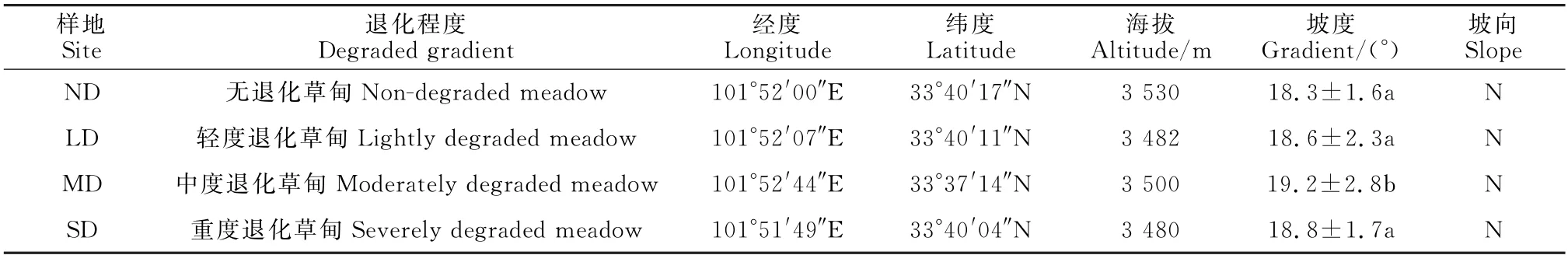
表1 样地基本信息Table 1 Basic information of sample plot
1.3 植物功能群对划分
研究区植物种类丰富,基于不同植物的生活型特征和功能特点,将研究区内的草本植物群落划分为3个主要功能群,依次是:禾草植物功能群、豆科植物功能群和杂类草功能群。其中,禾草功能群主要包含:密花早熟禾(Poa annua)、波伐早熟禾(P.poophagorum)和矮生嵩草(C.alatauensis)等禾本科和莎草科植物,其特点是对土壤养分和水分的利用率较高;豆科植物功能群主要包括:紫花苜蓿(Medicago sativa)和甘肃棘豆(Oxytropis kansuensis)等豆科植物,其特点是固氮能力强;杂类草功能群主要包括老鹳草(Geranium wilfordii)和唐松草(Thalictrum aquilegiifolium)等植物。通过将植物群落划分为不同的功能群,可以更好地理解植物群落的组成和生态功能,并对其进行更准确的描述和分析。
1.4 物种-多度拟合模型
针对各退化群落全部物种以及不同的植物功能群,采用三类(6种)模型拟合了其物种-多度分布关系。
1.4.1 统计模型
对数级数模型(log-series model,LS)。不考虑个体数为0的物种,负二项分布在k=0时的特殊分布模式即为对数级数模型,由本模型可知,群落中多度为n的物种数目S可表示为[10]:
式中:α为物种多样性指数,α始终大于0且与样本量没有关系,而X(0≤X≤1)作为常数受到样本大小的影响。
对数正态模型(log-normal model,NORM)。Preston最早提出将该模型应用于物种多度分布的研究,该模型认为群落中物种的个体数(即物种多度)符合正态分布,对于物种多度分布来说,变量为群落中每个物种的个体数,因子即影响群落组成的各种过程[11]。群落中第i个物种的多度A i表示为:
式中:μ为正态分布的均值;δ为方差;φ为正态偏差。
1.4.2 生态位模型
几何级数模型(geometric series model,GEO)。几何级数模型首先假设在1个生态系统中,所有物种共享相同的资源池,并且每个物种在资源池中都能占据一定的份额,每个物种以相同的时间间隔进入生境。将群落中物种数设定为M,第1个物种占领总生态位的a份,第2个物种占据剩余的a×(a-1)份,第3个物种占用剩下的a×(a-1)2份[12]。如果模型中占据最多资源的物种的多度用N1进行表示,那么第i个物种的多度Ni用公式表示为:
断棍模型(broken stick model,BRO)。该模型假定在群落中每个物种在资源利用上是相互竞争的,并且没有物种优势。在这种情况下,每个物种在资源利用上所获得的份额是随机的,符合均匀分布。因此,可以将总资源量等分为若干份,每个物种所获得的份额等于其占据的生态位的大小。然后,可以将所有份额从小到大排列,就形成了断棍模型[13]。若用S表示总物种多度,J表示群落中物种的个体总数,则第i个物种的多度A i表示为:
1.4.3 中性理论模型
复合群落零和多项式模型(metacommunity zero-sum multinomial distribution model,MZSM)。复合群落零和多项式模型的基本假设是:在1个封闭的群落中,如果1个物种的多度增加,另1个物种的多度会随之减少,双方变化的总和为0[14]。该模型需要取样点物种个体数J和多样性指数θ来支持,群落当中任意取样点内多度为n的物种数量S为:
式中:θ为基本多样性指数;J为群落总的物种多度;y为某个物种的多度;Γ(n)为Gamma分布,表示S个物种出现时所需的多度n的分布;t为某个物种含有n个个体的时间。
局域群落的VOLKOV 模型(volkov model,VOLKOV)。VOLKOV 基于对种群动态变化的微分方程描述,包括竞争种群的生长、死亡和迁移过程。在VOLKOV 模型中,竞争系数描述了不同物种之间的竞争强度,其中竞争强度越高,物种之间的竞争就越激烈。竞争系数的大小是通过测量物种间相对生长速率的比例来确定的。该模型的一大优点是可以预测在不同环境条件下物种丰富度和群落结构的变化。该模型还可以通过对竞争系数和其他参数的调整来模拟不同物种之间的相互作用,从而研究群落动态的稳定性和变异性[15]。局域群落中多度为n的物种数量S可表示为:
式中:n为某物种的多度;m为迁移率;J为群落样本大小;Γ(z)为z的函数;γ为迁移到局域群落的个体数。
1.5 模型拟合效果检验
K-S检验:K-S检验采用Kolmogorov-Smirnov(K-S)检验上述模型拟和效果与实际观测分布的差异。K-S检验是通过计算2个样本的经验分布函数之间的距离(统计量D)来判断二者是否存在显著差异,P<0.05,则表示该模型被拒绝。
AIC准则:采用AIC(akaike information criterion)评估模型拟合的优劣性。在模型选择时,当含有的参数个数越小,概率密度分布就与真实的数据越接近,AIC 值也就越小,模型拟合效果在此时达到最优。一般认为,模型之间AIC 值差异大于2时,2个模型有显著差异,当AIC值差异小于2时,认为2个模型的拟合效果相似。
1.6 数据处理与图像绘制
多度数据由不同退化程度各个样方中物种多度的平均值数据组成;使用Excel 2020 进行数据整理;使用PC-Ord软件5.0完成Bray-Curtis相异性分析,并使用双向聚类分析对所有物种进行分类;物种多度模型的拟合利用R(4.1.3)软件中的sads拓展包完成,物种多样性指数计算使用vegan拓展包完成;群落中植物的高度和盖度及功能群物种多样性指数使用SPSS计算平均值和标准误,并采用单因素方差分析(one-way ANOVO)来检验不同处理间的差异性。K-S 检验使用R(4.1.3)软件中的Matching拓展包完成;功能群多样性指数参照马克明[16]的计算,物种重要性值参照王育松等[17]的计算。使用Origin 2021完成相关图形的绘制。
2 结果与分析
2.1 不同退化程度草甸植物群落物种组成及其变化
随着高寒草甸退化程度增加,群落的物种组成发生了明显变化(表2),不同退化程度高寒草甸的盖度、高度显著降低(P<0.05)。无退化草甸植被类型为早熟禾(Poa annua)+披碱草(Elymus dahuricus);轻度退化草甸的植被类型为:矮嵩草(Carex alatauensis)+早熟禾(Poa annua);中度退化草甸的植被类型为细叶亚菊(Ajania tenuifolia)+早熟禾(Poa annua);重度退化草甸的植被类型为:西伯利亚蓼(Knorringia sibirica)+条叶银莲花(Anemone obtusiloba)。随着高寒草甸退化程度的加深,植被优势种从无退化草甸的以禾本科和莎草科植物为主,逐渐变为杂类草植物为主。图1表明,不同退化程度的高寒草甸样地可以分为4类,不同退化程度各为一类,无退化草甸与轻度退化草甸植被组成较为接近,中度退化草甸与重度退化草甸植被组成较为接近。研究区内的66种植物可分成6类。

图中黑色方框代表物种在样方内存在,白色方框代表物种不存在,ND为无退化草甸的样方,LD为轻度退化草甸的样方,MD为中度退化草甸的样方,SD为重度退化草甸样方。图1 不同退化程度草甸样地与植物分布Black represents the presence of species.White represents the absence of species.ND is a sample of non-degraded meadow.LD is a sample of lightly degraded meadow.MD is a sample of moderately degraded meadow.SD is a sample of severely degraded meadow.Fig.1 Distribution of sample plots and plants with different degrees of degradation
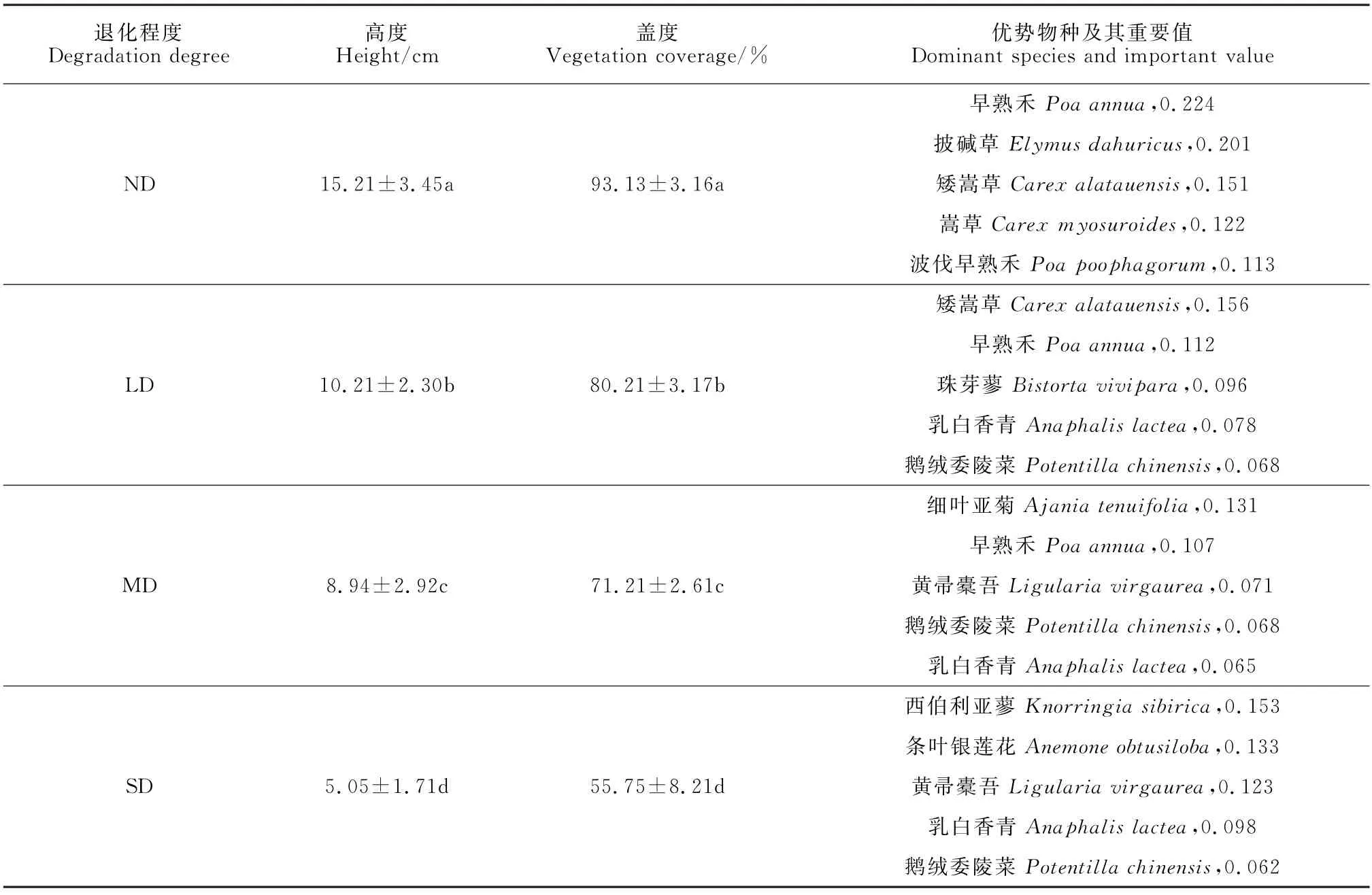
表2 不同退化程度草甸高度、盖度及其优势物种变化Table 2 Changes in dominant species with different degraded gradient
2.2 不同退化程度草甸植物功能群多样性
对功能群多样性进行分析,结果(表3、图2)表明,同一退化梯度上各功能群的多样性指数变化趋势基本一致,杂类草的多样性指数最大,其次为禾草功能群,豆科功能群的多样性指数最小。

图2 高寒草甸植物功能群多样性指数随退化程度的变化趋势Fig.2 Variation trend of plant functional group diversity index with degradation degree in alpine meadow
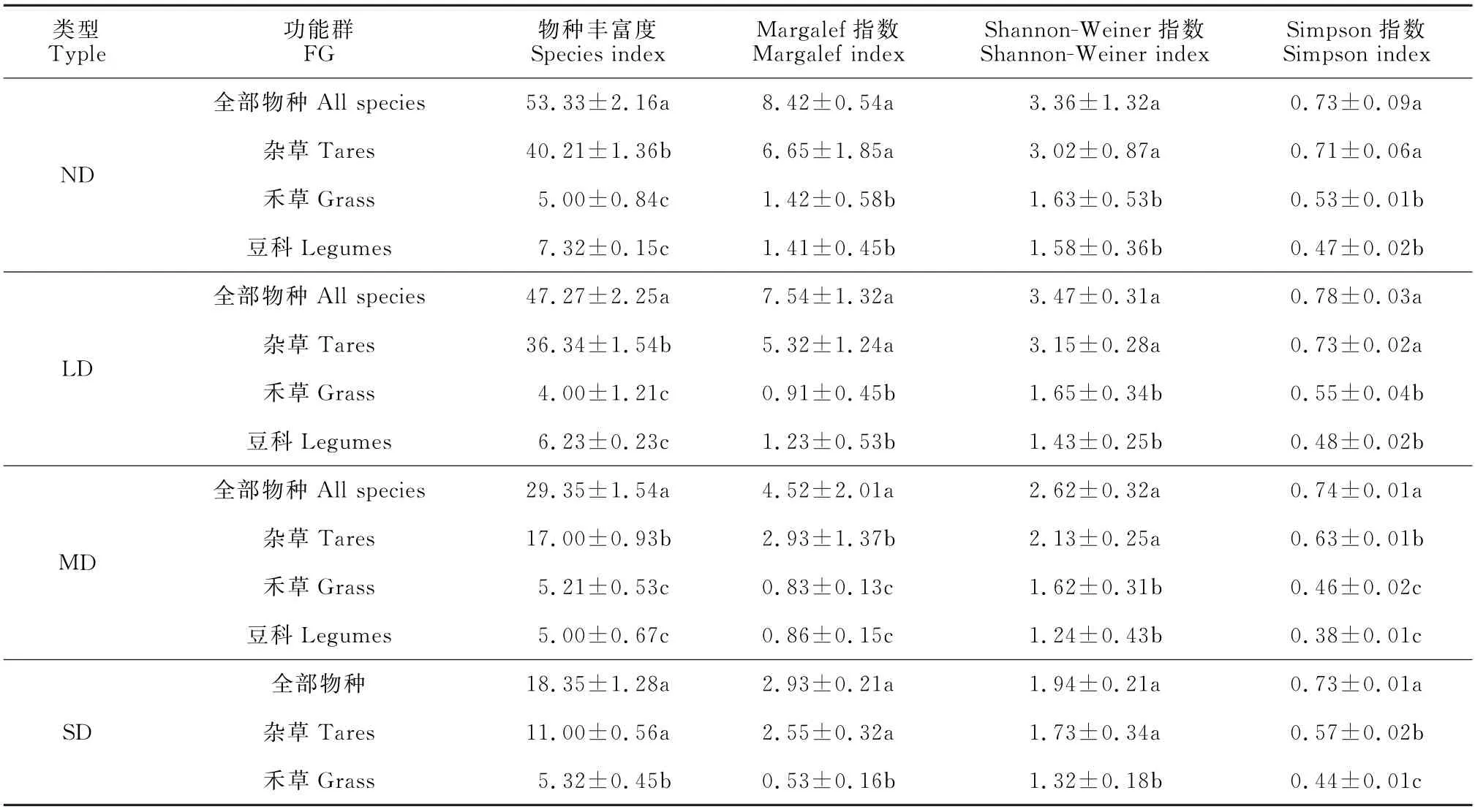
表3 不同退化程度草甸植物功能群多样性指数的变异Table 3 Variation of diversity index of plant functional groups of meadows with different degraded degrees
全部物种、杂类草功能群与禾草、豆科功能间多样性指数的差异性显著(P<0.05)。在不同退化梯度上,杂类草功能群的丰富度呈现逐渐减小的趋势,禾草功能群和豆科功能群丰富度变化较为平缓,杂类草功能群的变化趋势和全部物种的变化趋势相似,禾草功能群和豆科功能群的物种丰富度明显低于杂类草功能群;杂类草功能群、禾草功能群和豆科功能群的Margalef指数随退化程度加深而减小,其中杂类草功能群与全部物种的变化趋势相似,而禾草功能群和豆科功能群的变化趋势相对平缓;杂类草功能群的Shannon指数随着退化程度的加深,先增加后减少,与全部物种的Shannon指数变化趋势类似,呈现单峰变化,禾草功能群与豆科功能群的Shannon指数随着退化程度的加深而减小,且低于全部物种的Shannon指数;杂类草功能群和禾草功能群的Simpson指数均表现为先增加后减小,豆科功能群的Simpson指数随退化程度的加深而减小,显著低于全部物种和杂类草功能群的Simpson指数。
2.3 不同退化程度草甸全部物种多度分布的模型选择与比较
选择LS、NORM、BRO、GEO、VOLKOV 和MZSM 6种模型对不同退化程度草甸全部物种多度分布进行拟合与优势度检验。由K-S检验结果(表4)可以得出,LS和NORM 在拟合轻度退化草甸群落物种多度分布时,未能通过检验(P<0.05)。AIC检验表明:无退化草甸的最优拟合模型为VOLKOV,轻度退化草甸的最优拟合模型为GEO模型,中度退化草甸的最优拟合模型为BRO,重度退化草甸的最优拟合模型也为BRO,且与VOLKOV 模型的AIC值差<2,二者拟合效果相似。
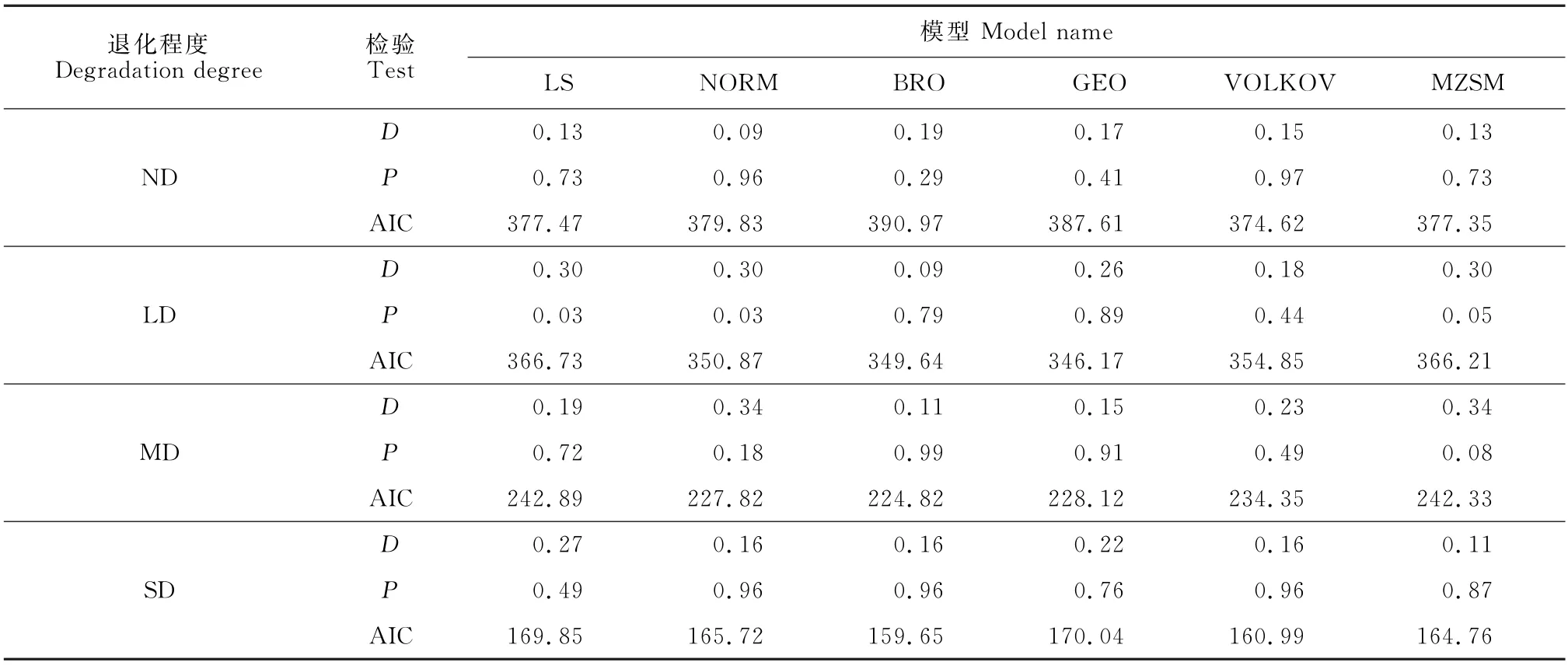
表4 不同退化程度草甸全部物种多度分布模型拟合优势度检验Table 4 Species abundance models fitted dominance text of meadows in different degradation degree
由实测值的多度分布曲线可知,退化程度越高,相对多度曲线斜率越小,无退化群落的相对多度曲线最为平缓。4 种不同退化程度草甸物种-多度关系的拟合值与实测值见图3。

图3 不同退化程度草甸全部物种的拟合值实测值对比Fig.3 Comparison of observed and fitted values of meadows in different degradation degree
2.4 不同功能群多度分布的模型选择与比较
2.4.1 禾草功能群
K-S检验结果(表5)显示,所有模型均通过检验(P>0.05)。AIC 检验结果及禾草功能群物种多度拟合值实测值对比(图4)表明:对于禾草功能群,在无退化草甸和轻度退化草甸,最优拟合模型主要是BRO,在中度退化草甸和重度退化草甸,最优拟合模型既有BRO 和GEO,也有NORM 模型,表明其资源分配方式由固定分配和随机分配共同主导。
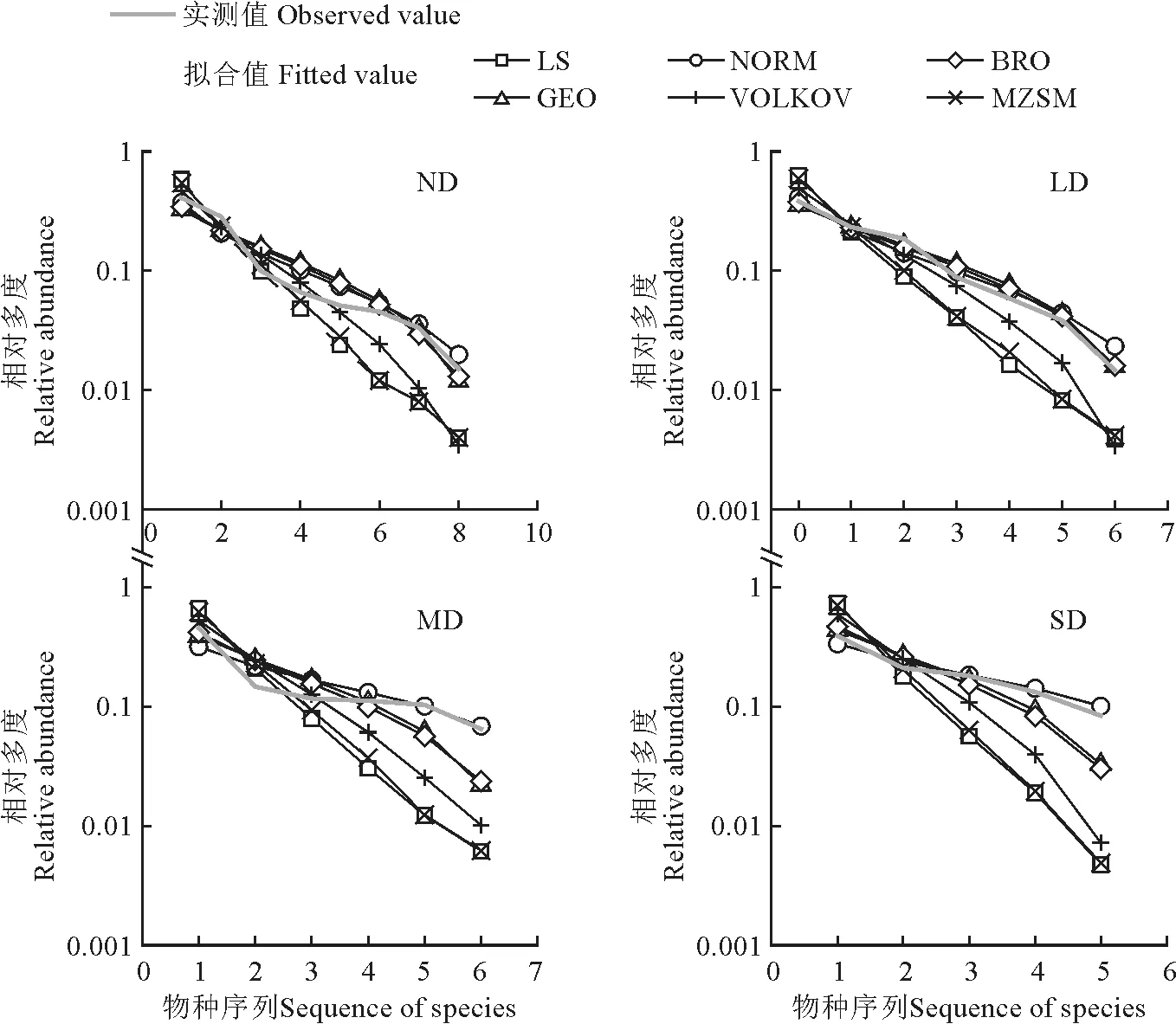
图4 不同退化程度草甸禾草功能群物种多度拟合值实测值对比Fig.4 The species abundance model fitting of grasses functional groups

表5 不同退化程度草甸禾草功能群物种多度分布模型拟合优势度检验Table 5 Testing the dominance of model fit for species abundance distribution of grasses functional groups
2.4.2 豆科功能群
对不同退化程度草甸豆科功能群多度分布的拟合与优势度检验表明,所有模型均通过了K-S检验(表6)。AIC检验及豆科功能群物种多度拟合值实测值对比(图5)表明最优拟合模型均为BRO,资源分配方式以固定分配为主。
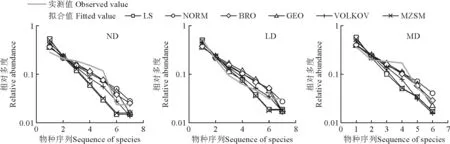
图5 不同退化程度草甸豆科功能群物种多度拟合值实测值对比Fig.5 The species abundance model fitting of legumes functional groups
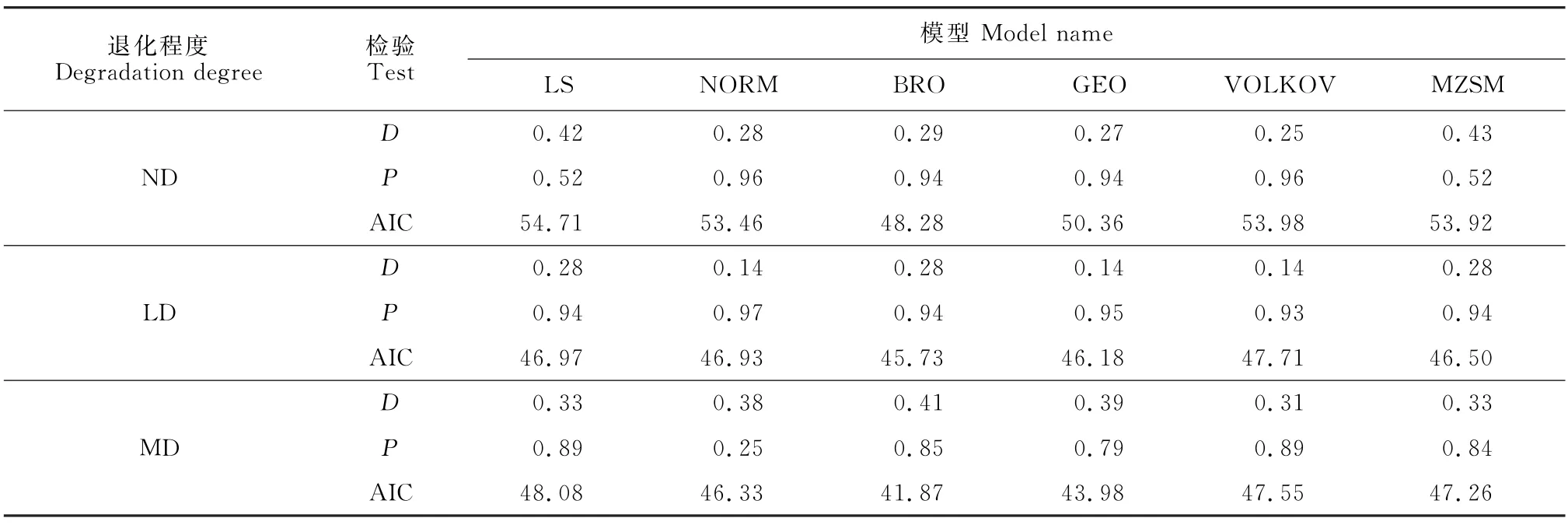
表6 不同退化程度草甸豆科功能群物种多度分布模型拟合优势度检验Table 6 Testing the dominance of model fit for species abundance distribution of legumes functional group
在重度退化草甸,豆科功能群退出群落。在轻度退化草甸,所有选取的模型都有较好的拟合效果,表明所有模型所代表的资源分配方式均有一定的指导意义。
2.4.3 杂类草功能群
对杂类草功能群而言,K-S检验结果(表7)表明LS与MZSM 在拟合轻度退化草甸物种多度分布时未能通过检验(P<0.05)。AIC检验及杂类草功能群物种多度拟合值实测值对比(图6)表明,无退化草甸的最优拟合模型为VOLKOV,轻度、中度和重度退化草甸为BRO,最优拟合模型从中性模型向生态位模型转变,其资源分配由随机分配向固定分配转变。
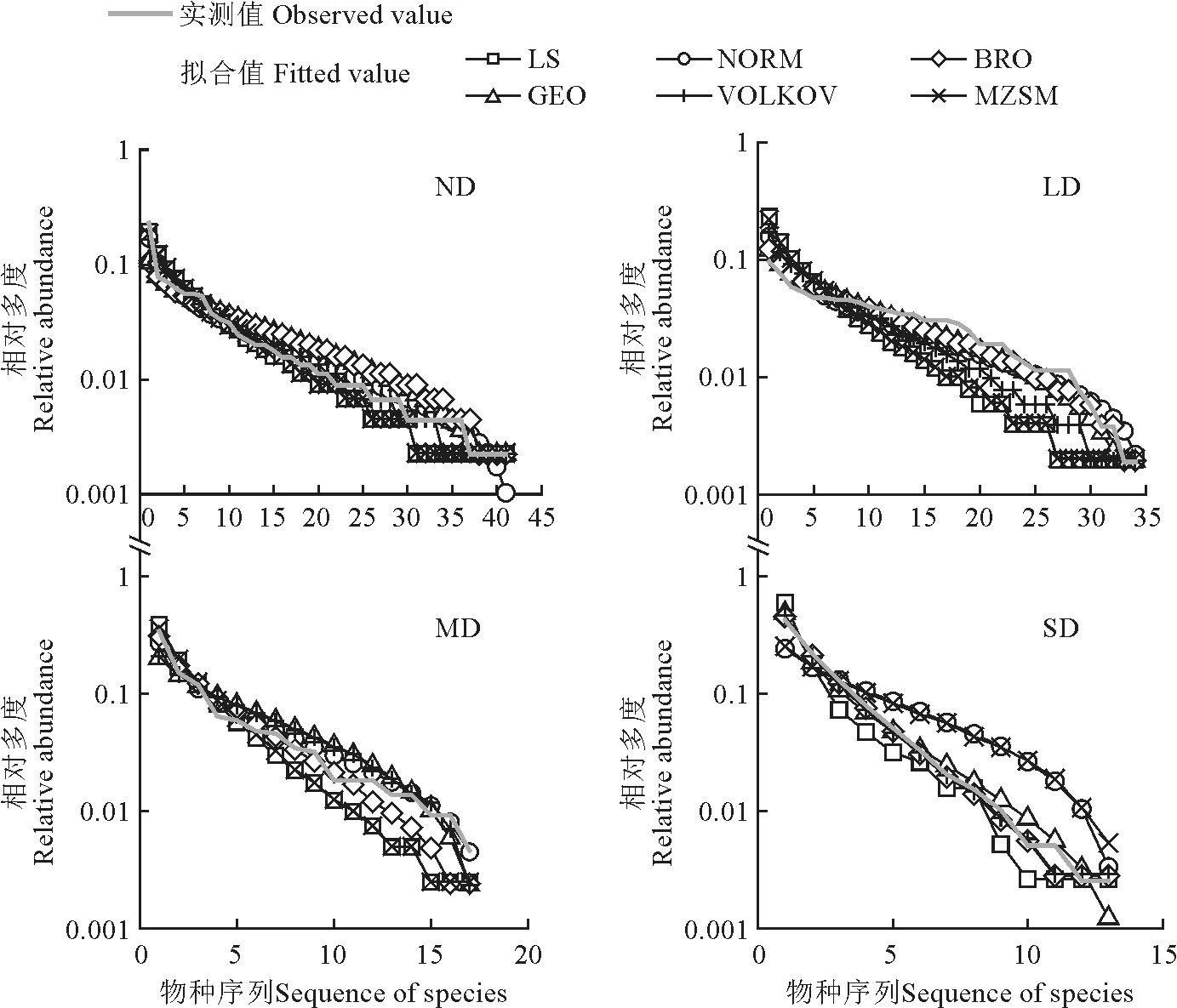
图6 不同退化程度草甸杂类草功能群物种多度拟合值实测值对比Fig.6 The species abundance model fitting of miscellaneous grass functional groups

表7 不同退化程度草甸杂类草功能群物种多度分布模型拟合优势度检验Table 7 Testing the dominance of model fit for species abundance distribution of miscellaneous grass functional groups
3 讨论
3.1 植物功能群多样性对退化的响应
植物功能群连接植物群落和生态系统过程,其结构可作为高寒草甸退化的标志,保持功能群多样性能够维持植物群落功能最大化[18]。本研究通过计算植物功能群的多样性指数,探究不同植物功能群对群落多样性的贡献,结果表明,同一退化梯度上各功能群的4个多样性指数的变化趋势基本一致(图1、图2)。随着退化程度的增加,杂类草功能群的丰富度和均匀度均显著下降,而豆科功能群和禾草功能群的丰富度和均匀度指数的下降趋势较为平缓,但杂类草功能群在草甸群落中的多样性和丰富度均保持着较高水平,其多样性指数与整体群落最为接近,这表明杂类草功能群在群落多样性分布中起着主导作用,对群落多样性的贡献更大,杂类草在植物群落资源分配中占据主导地位。
在甘南高寒草甸,无退化草甸植物群落优势种主要是禾草功能群植物,但是由于放牧等活动的干扰,导致禾本科和莎草科等适口性较好的禾草功能群植物被采食,为杂类草功能群提供了生态位,加剧了杂类草入侵,使得轻度退化草甸杂类草功能群的多样性最高,随着退化的加剧,高寒草甸向盐碱化和干旱化变化,这种变化导致大多数物种无法继续生存,只有能适宜恶劣生境的植物能够继续生长,最终使得所有功能群的丰富度指数和多样性指数大幅度下降[18];另一方面,随着草甸的退化,土壤含水率降低,容重增加,使得能够适应干旱、贫瘠、盐碱环境的杂类草功能群植物取代了湿生、根系较浅的禾草功能群与豆科功能群植物[19]。另外,随着退化程度加深,生境的水热条件发生改变,形成不同的微气候,群落以降低禾草和豆科功能群的多样性为代价增加了杂类草功能群的多样性,杂类草功能群对资源竞争的频率不断增加。面对环境变化和干扰,不同功能群的植物往往会采取比如生长适应策略、繁殖适应等策略来对退化进行响应[20],群落内不同功能群间的相互作用,有助于适应不断变化的环境条件并缓解草甸退化所带来的影响,相互作用的差异也会决定整个群落对于环境退化的适应能力。
3.2 高寒草甸植物群落物种多度分布对退化的响应
物种相对多度曲线分布图可以直观展示群落的均匀度和丰富度,物种多度分布曲线的越平缓,说明群落中物种多度存在的差异越小,群落物种的均匀度也就越大;反之,相对多度分布曲线越陡,说明群落物种分布就越均匀度越低[21]。本研究中,随着高寒草甸退化程度的加深,全部物种实测值的相对多度曲线斜率越来越小(图3),物种丰富度随之降低,无退化群落的相对多度曲线最为平缓。退化改变了群落中物种的丰富度与均匀度,与其他生境相比,无退化草甸群落物种分布最均匀,物种丰富度最高,多样性最大,而重度退化生境与之完全相反。这可能是由于无退化草甸,水分和养分的含量相对较高,加之人类活动的影响较小,但随着退化程度的加深,植被盖度降低,地表径流增大,土壤的养分和水分流失越发严重,草甸变得贫瘠,生境逐渐恶化,能容纳的物种减少,不足以支撑更多的物种进行生长发育等活动,最终影响植被的空间分布,因此在无退化草甸的物种多样性最高,丰富度和均匀度也最大。这也与周宸宇等[22]的研究结果一致,物种的丰富度会受到高寒草甸退化的显著影响,最终表现为优势种丧失,群落结构简单化,群落稳定性降低。
植物功能群与群落稳定性、生产力和退化恢复密切相关[8],功能群的引入,有助于更好地认识群落的生物多样性和生态系统结构,将不同退化程度的高寒草甸划分为禾草功能群、豆科功能群和杂类草功能群并对其多度分布进行模型拟合。全部物种在无退化草甸的最优拟合模型为中性模型,轻度、中度和重度退化草甸的最优拟合模型均为生态位模型,全部物种群落构建由资源的随机分配为主转向以资源的固定分配为主,这可能与无退化草甸资源丰富有关。对于禾草功能群,在无退化草甸和轻度退化草甸,最优拟合模型主要是BRO,而在中度退化草甸和重度退化草甸,最优拟合模型既有BRO 和GEO,也有代表随机分配的NORM 模型(表5、图4)。对于豆科功能群,其最优拟合模型始终为BRO(表6、图5)。在无退化草甸,受干扰较小,植被生长状况较好,各功能群稳定共存,相比于竞争作用,功能群内种间的资源共享作用更大[21],此时禾草功能群与豆科功能群多度分布主要受生态位划分的影响,资源以固定分配为主。尽管最优拟合模型相同,但群落的形成可能存在差异[23],随着退化程度的增加,草甸生境逐渐恶劣,加之禾草功能群和豆科功能群是牲畜主要采食的对象,由于家畜的反复采食,会使得适口性较好的禾草功能群与豆科功能群没有时间去发展进化,一定程度上增强了其种内竞争[24],使得部分物种被淘汰,未被淘汰的植物占据着稳定的生态位而共存,因此,BRO 很好地拟合了轻度退化草甸禾草功能群和豆科功能群的物种多度分布状况,其资源分配以固定分配为主。对于中度退化和重度退化草甸的禾草功能群,NORM的拟合效果与BRO 拟合效果相似,NORM 认为资源的分配是随机进行的[15],这可能是由于过度的干扰使得禾草功能群生存不稳定,使得其资源分配出现随机性。对于禾草功能群,其资源分配的模式从固定分配模式转向随机分配和固定分配共存的分配模式。豆科功能群的最优拟合模型始终为BRO 模型,其资源分配始终是以固定分配为主。
随着退化程度的加深,杂类草功能群的最优拟合模型由中性模型向生态位模型转变(表7、图6),资源分配由随机分配向固定分配转变。对于无退化草甸,优越的环境资源维持着杂类草功能群相对较高的群落物种多样性与多度,在物种多样性高的功能群中,中性过程是物种多度分布格局形成的主要驱动力[24]。草甸进入退化状态后,环境恶化,种内竞争和种间竞争加剧,在严酷生境中,以生态位为基础的环境筛选、生态位分化常常被认为是群落构建的主导过程[25],杂类草功能群抵抗各类因素的干扰能力较强,使得杂类草功能群可以占据更大的生存空间,生长状态比其他功能群的生长状态更稳定,资源分配方式以固定分配为主。
4 结论
(1)随着退化程度加深,植被优势种由禾本科和莎草科植物为主逐渐变为以杂类草功能群植物为主。在同一退化梯度,功能群多样性指数的变化趋势一致,而随着退化程度的增加,杂类草功能群在草甸群落中的多样性和丰富度均保持着较高水平,这表明杂类草功能群在群落多样性分布中起主导作用。
(2)全部物种多度分布随着退化程度加深发生变化,其中,无退化草甸的最优拟合模型是中性模型,轻度、中度和重度退化草甸的最优拟合模型是生态位模型,资源分配模式由随机分配转向固定分配的分配模式;禾草功能群的最优拟合模型以生态位模型为主,资源分配方式由固定分配和随机分配共同主导;豆科功能群的最优拟合模型是生态位模型,资源分配方式以固定分配为主;杂类草功能群的最优拟合模型由中性模型向生态位模型转变,与全部物种多度分布的最优模型基本一致,资源分配由随机分配向固定分配转变,可以认为杂类草功能群是影响群落物种多度分布的主要原因,但禾草和豆科功能群的贡献也不可忽视。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
草地学报(2021年7期)2021-08-03 13:37:10
科学中国人·上半月(2021年4期)2021-07-20 19:01:37
活力(2019年21期)2019-04-01 12:17:10
南方农业·下旬(2017年11期)2018-01-09 14:37:50
中国饲料(2017年15期)2017-08-22 04:38:29
农业环境科学学报(2017年2期)2017-03-20 14:57:30
杂草学报(2015年2期)2016-01-04 14:57:54
