
补体蛋白调控靶点在糖尿病视网膜病变中的作用及临床研究进展
2024-03-05 10:25:16王绪瑞孙洪岩杨明明
国际眼科杂志 2024年3期
王绪瑞,孙洪岩,2,刘 丹,杨明明,2
0 引言
糖尿病是一组以高血糖为特征的代谢性疾病,近年来,世界范围内糖尿病患病率不断增加,造成严重的社会负担。糖尿病可引起大血管、微血管的多种并发症,如代谢性白内障、动眼神经损伤、眼部炎症、屈光不正等,其中,糖尿病视网膜病变(diabetic retinopathy,DR)被认为是最常见和损害性最严重的眼部并发症,且影响人群广泛,治疗预后较差,是目前眼科领域研究的重点及热点话题。DR的发病机制错综复杂,目前认为,长期暴露于高血糖和其他致病危险因素(如高血压等)启动了眼内的一系列生理和生化变化包括氧化应激、蛋白激酶C(PKC)的活化、炎症反应、多元醇代谢通路的异常等,并最终导致微血管损伤和视网膜功能障碍[1-2]。近年来有越来越多的证据表明,DR中的视网膜微血管损伤与补体系统激活密切相关[3]。视网膜是一种免疫豁免组织,有其独特的免疫系统,包括血-视网膜屏障(blood-retinal barrier,BRB)、先天免疫细胞(如小胶质细胞群等)和补体系统[4]。在衰老时,天然免疫机制(特别是补体系统)低水平激活,从而维持正常的眼稳态和视网膜完整性[5-6]。然而,如果补体持续激活的时间较长,就会导致不可逆转的功能丧失,从而产生有害的影响。因此,本文从多种补体蛋白探讨补体系统在DR发生发展中的作用。
1 补体系统
补体系统广泛存在于人体血清、组织液和细胞膜表面,被熟知的组分包括30余种,是一个具有精密调控机制的、具有免疫效应放大的先天蛋白质反应系统。通常,多数补体成分在血浆中是无活性的前体物质(酶原),只有在被激活物激活后才具有其特殊的生物学功能。许多内源性或外源性物质(包括抗原-抗体复合物、多种微生物成分等)可通过三条既独立又交叉的途径,通过一系列级联酶解反应,进而激活补体系统,三条途径即经典途径、旁路途径和凝集素途径(MBL途径)(图1)。所有这三条途径所形成的活化产物具有调理吞噬异物、调节免疫应答、介导炎症反应、清除免疫复合物和溶解细胞等功能。

图1 补体系统激活的三条通路。
1.1经典途径激活物首先与C1q结合,顺序活化C1r、C1s、C4、C2、C3等多种补体蛋白,形成C3转化酶(C4b2a)与C5转化酶(C4b2a3b)的级联酶促反应过程。C5可被裂解为C5b,在后续的级联反应中与C6、C7、C8、C9相继结合,最终形成攻膜复合物(membrane attack complex,MAC)。经典途径的补体激活在多种眼科疾病中已被证实发挥重要作用,包括青光眼、年龄相关性黄斑变性(age-related macular degeneration,ARMD)、自身免疫性葡萄膜炎等,特别是本文着重论述的DR[7-9]。
1.2旁路途径旁路途径又称替代激活途径,一般是由微生物或外源异物直接激活,而不依赖于机体抗体的形成,在B因子(CFB)、D因子(CFD)和备解素P因子(CFP)等多种重要因子的参与下,多种补体蛋白形成旁路途径的C3、C5转化酶,进而启动级联酶促反应过程。旁路途径虽然被发现较晚,但其在补体系统中具有重要地位,是目前研究的热点话题。我们团队的多项研究发现其与前葡萄膜炎、DR等息息相关[10-14]。
1.3凝集素途径凝集素途径又称MBL途径(MBL pathway),与经典/旁路途径不同的是,病原体表面糖结构被血浆中的甘露糖结合凝集素(mannose-binding lectin,MBL)、纤维胶原素(ficolin,FCN)等直接识别,进而活化MBL相关丝氨酸蛋白酶(MBL-associated serine protease,MASP),随后依次活化C4、C2、C3、C5,最后进入补体激活的末端通路。MBL途径常被研究者忽略,但其仍拥有与众不同的作用,它的激活物质非常广泛,并且对经典途径和旁路途径的活化具有交叉促进作用[15]。
综上所述,以上三种途径均可通过融合并刺激C3/C5转化酶的形成,随后共同进入末端通路,最终形成MAC。在此基础上形成的MAC可溶解细胞或导致细胞死亡[16],因此,常常认为补体系统的激活是把双刃剑,不适当的补体活化会导致机体损伤,从而引发多种疾病。
2 多种补体蛋白在DR中的作用
2.1全身和局部补体激活参与DR的发生人体适当的免疫反应对于消除外源性和内源性因素引起的组织损伤是必不可少的,人眼也不例外。在正常人眼中,先天免疫系统通过精密的调节,严格控制免疫反应以防止不可逆的组织损伤,从而维持视网膜的稳态。免疫和炎症机制在DR发病机制中的作用一直得到许多业内人士的重视,补体系统作为先天免疫的重要组成成分,目前多项研究证明补体系统在糖尿病眼中发挥重要作用[3-4]。Gerl等[17]报道了DR患眼的Bruch膜、脉络膜毛细血管中广泛补体激活的证据,通过免疫组织化学检测发现补体相关蛋白的密集染色,提示DR患眼中存在补体激活。Pauly等[18]通过对92 000个小鼠视网膜细胞进行单细胞RNA测序,发现了小鼠视网膜细胞类型的独特补体表达信号,表明视网膜微环境中局部补体表达的调节。这提示了一个有趣的概念,即局部视网膜补体激活可能独立于通常由肝脏产生的补体成分。此外,Mandava等[19]研究证明这种局部激活是持续的,并且与系统性补体活性分离。因此,补体系统在DR中的作用可能包括全身系统性的作用,以及局部的激活与调节。
2.2经典途径相关调控靶点与DR C1、C4和C2是经典途径前段重要的补体蛋白。在经典途径中,激活物一般是抗原-抗体复合物,其首先与C1q结合,顺序活化C1r、C1s、C4、C2、C3,启动下一步的级联反应。相关研究表明,C1、C4和C2与DR密切相关,Huang等[20]研究发现缺乏IgG外泌体的糖尿病小鼠视网膜血管损伤相对减少,在此基础上,通过进一步C1激活实验推测血浆中载有IgG的外泌体可能通过结合并水解激活C1复合物,进而激活经典补体途径,最终损伤下游视网膜血管,而其中的分子机制和通路可能是未来需要研究的方向。García-Ramírez等[21]研究增殖性糖尿病视网膜病变(PDR)患者的蛋白质谱,通过采用荧光差异凝胶电泳(DIGE)进行蛋白质组学分析,发现C4b、C3等相关补体蛋白在PDR患者的玻璃体中明显升高,该研究进一步进行蛋白免疫印迹实验证实C3的这种差异表达,且与视网膜中检测到的mRNA水平差异一致,说明经典途径补体激活参与DR的发病,且可能是参与疾病的后期阶段。我们团队在前期研究中发现,DR患者血清中C4呈高表达,C4是活化的C1s的第一个底物,因此DR病理进程中可能有经典途径补体系统的激活和补体所诱发的炎症反应[22]。有趣的是,在另一个炎症性疾病视神经炎中,有学者发现血清补体C3、C4水平下降[23]。此外,Pauly等[18]研究发现Müller细胞是补体激活因子C1s、C3、C4和CFB的主要贡献者,衰老增强C1s、CFB、CFP和I因子(CFI)表达,H因子(CFH)表达降低,短暂性视网膜缺血增加小胶质细胞、Müller细胞和视网膜色素上皮(RPE)细胞中补体的表达。
2.3旁路途经相关调控靶点与DR CFB、CFH是补体旁路途径的重要调节蛋白。CFB裂解形成的Bb可与C3b结合形成C3bBb,此为旁路途径C3转化酶,启动下一步级联反应。CFH能够抑制旁路途径的C3/C5转化酶(C3bBb/C3bBb3b),发挥负性调节作用。CFI是一种丝氨酸蛋白酶,在辅助因子存在的情况下能够抑制C4b2a、C4b2a3b、C3bBb、C3bBb3b的形成与活性,抑制补体级联反应。近年,CFB和CFH的基因多态性与DR的关系越来越成为眼科医生的研究热点。García-Ramírez等[21]通过检测PDR患者玻璃体液中的CFB,证实了旁路途径的参与。Toni等[24]发现rs1410996(CFH)多态性可能与1型糖尿病患者的PDR相关。我们团队在一项研究中分析DR和糖尿病对照组CFB和CFH基因多态性的关系,结果显示,与糖尿病对照组相比,DR患者的rs1048709(CFB)A等位基因和AA基因型频率显著升高;而rs800292(CFH)A等位基因和AA基因型频率显著降低,且rs800292/AA基因型与DR的延迟进展有关[14],因此认为CFH可能是DR的一个保护性因素。无独有偶,另一项研究中rs800292(CFH)的AG基因型被认为可能是ARMD的保护因素[25]。关于眼内CFH的来源,有学者认为,DR中CFH来源于激活的小胶质细胞,DR患者眼内液中检测到CFH水平的升高可能是作为抑制补体过度激活的反馈机制[26],基于此,深入靶向小胶质细胞介导的补体激活的研究可能为靶向治疗DR提供新的思路。尽管CFI与DR发病之间的联系尚未完全明确,但编码CFI的基因突变与患ARMD的风险增加之间的关系已得到证实[27]。另有研究表明CFI参与急性前葡萄膜炎的发病过程[10]。因此,CFI与DR之间的关系值得进行更深入的研究。
2.4MBL途径相关靶点与DR 血浆中的MBL可特异性识别表面糖结构是甘露糖、甘露糖胺等为末端糖基的多种病原体,然后活化MASP,启动下一步级联反应。一般情况下,MBL不与机体自身组织结合,但在糖尿病或组织缺血等情况下,细胞表面糖基化模式的改变会显著增加MBL的自身反应性,从而导致糖尿病患者的免疫激活增加,局部和全身炎症增加,并伴随糖尿病晚期并发症的风险增加。补体系统与糖尿病血管并发症之间的联系的潜在机制之一可能是糖尿病患者动脉壁晚期糖基化终末产物(advanced glycation end products,AGEs)的形成,这些产物为MBL的结合产生了新的结合位点[28]。在许多临床前沿研究中,MBL途径与糖尿病微血管和大血管改变之间的联系已被证实。有研究分别检测1型和2型糖尿病患者血浆MBL水平,发现其高于健康人[29-30]。另有横断面研究发现,DR患者MBL水平与DR严重程度存在正相关性,提示MBL途径可能在DR的发病机制中发挥作用[31]。Holt等[32]随访研究18 a,首次发现血清中高水平MAp44(MBL途径蛋白)与DR的发展之间存在显著关联,但MAp44被认为是MBL途径的抑制剂,因此MAp44的升高可能是代偿性的上调亦或是其他作用,这需要进一步探索研究。另外,该研究还发现高水平的H-ficolin与单纯视网膜病变的发生相关,提示MBL途径补体激活在DR的早期阶段可能起着更为突出的作用。遗憾的是,目前关于DR患者中MBL基因和MASP基因多态性的研究仍较为空白,这值得进一步研究。
2.5核心补体蛋白C3和C5与DR C3和C5是补体活化的共同组分,C3可被C3转化酶裂解为C3a、C3b,C3b还可以进一步裂解为C3c、C3dg、C3d等小片段,C5可被C5转化酶裂解为C5a、C5b。C3a和C5a都游离于液相,是重要的炎症介质,C3d可参与适应性免疫应答。已有相关研究表明,C3和C5在DR患者体内表达增加,Zhang等[33]发现DR患者血清中C3a和C5a水平高于正常人。Gao等[34]发现PDR患者玻璃体中C3等补体蛋白表达增加。另外,Muramatsu等[35]研究报告PDR患者玻璃体中检测到C5a表达增加,并发现C5a浓度与血管内皮生长因子(vascular endothelial growth factor,VEGF)、单核细胞趋化蛋白-1(monocyte chemotactic protein-1,MCP-1)等炎症细胞因子水平显著相关,提示C5a可能与炎症细胞因子等协同参与病理性的血管新生,在PDR的发病机制中发挥重要作用。此外,Cheng等[36]发现,视网膜Müller细胞高表达C5aR,C5aR可与C5a结合导致白细胞介素(IL)-6和VEGF的释放,参与DR的病理过程[36]。也有学者发现C5基因多态性与DR发生相关。我们团队通过分析多个补体通路基因与DR发病的关联性,首次发现rs17611(C5)与2型糖尿病患者视网膜病变之间存在弱关联,揭示DR与C5多态性之间的关联性,表明补体途径与DR发病机制有关[37]。Xu等[38]发现C5基因rs2269067(C5)与2型糖尿病患者发生PDR的风险相关,且该基因型还与IL-6产生增加相关。因此,C3和C5及其相关成分可能还通过炎症机制参与病理性的血管新生。C3、C5是补体级联反应中的核心步骤,未来研究可进一步探讨C3、C5和炎症在DR血管新生中的具体分子机制,以及如何通过药物或基因治疗干预DR的发展。
2.6补体抑制剂CD55和CD59与DR CD55蛋白是补体通路中的一种调控蛋白,也被称为补体衰变加速因子(DAF),其通过糖基磷脂酰肌醇(GPI)连接固定在细胞膜上,广泛分布在不同的组织细胞表面和细胞分泌物中,CD55蛋白的主要作用是加速C3转化酶和C5转化酶中C2a和Bb因子的衰变解离进而抑制补体系统激活。CD59也是一种GPI锚定蛋白,大多数有核细胞的膜上都存在这种特殊蛋白,CD59能够在补体级联反应的末期通过竞争性结合C8上的新生表位,抑制C9与C5b678的结合,从而阻止MAC的形成。MAC可直接引起细胞裂解并刺激细胞释放炎性细胞因子。已有多项研究表明,MAC在糖尿病患者的视网膜中沉积增加[17],MAC参与DR的发病机制肯可能是MAC沉积直接损伤视网膜细胞如周细胞[39],细胞损伤也可以促进炎症的发生。CD55和CD59
都是补体系统的抑制剂,既往研究发现糖尿病供者的视网膜CD55和CD59水平显著降低,表明调节机制的丧失可能是糖尿病患者视网膜补体激活增加的原因,且病程较长的糖尿病患者(9±3 a)视网膜MAC沉积增加[40]。另有研究发现一种被称为可溶性CD59(sCD59)的MAC抑制剂能够减轻糖尿病小鼠视网膜的损伤,该研究采用基因治疗方法将腺相关病毒载体(AAV2/8-sCD59)递送到小鼠眼部,结果发现sCD59能够减弱视网膜血管渗漏到玻璃体,抑制视网膜血管无灌注区域的产生、视网膜神经节细胞凋亡和MAC沉积,从而发挥对视网膜的保护作用[41]。此外,该研究团队还报道了sCD59对ARMD和自身免疫性葡萄膜炎(EAU)的作用[42-43],希望未来能应用于临床。
3 靶向补体抑制剂的临床应用
补体系统参与的眼科疾病众多,针对补体系统的靶向补体药物研究目前也在临床中取得了重大进展,并且拥有广阔的前景。补体C5抑制剂Eculizumab曾被研究用于ARMD的治疗[44],尽管其疗效不高,但其在视神经脊髓炎谱系疾病(NMOSD)的治疗中是一种有效且耐受性良好的治疗方法[45]。C3抑制剂Pegcetacoplan被证明可以延缓ARMD中地图样萎缩(GA)的进展,并且该药的3期临床试验正在进行中[46-47]。此外,该药还被应用于夜间阵发性血红蛋白尿(PNH)的治疗。另一种C5抑制剂Avacincaptad pegol(Zimura)也在ARMD的治疗中取得了良好的疗效[48]。此外,一款具有抑制VEGF和补体激活(C3b/C4b)双重作用的药物Efdamrofusp alfa被证明在体外具有良好的安全性和抗血管生成作用,并且目前也已完成用于新生血管性年龄相关性黄斑变性(neovascular age-related macular degeneration,nARMD)治疗的1b期临床试验[49-50]。针对CFH、CFB、CFI的药物也正在临床研究中[51-53],我们期待有更好的成果服务眼科疾病。在其他眼科疾病中,自身免疫性葡萄膜视网膜炎[54]和青光眼[55]也在近年的临床研究中取得了良好的效果,并且有望被列入补体靶向药物治疗的候选名单。总之,补体靶向药物具有广阔的研究前景,尽管目前的药物在眼科的应用仍着重于ARMD的治疗,但随着补体系统在各个眼科疾病的作用机制逐渐被发掘,补体靶向药物有望应用于包括DR在内的众多眼科疾病的治疗(表1[56])。
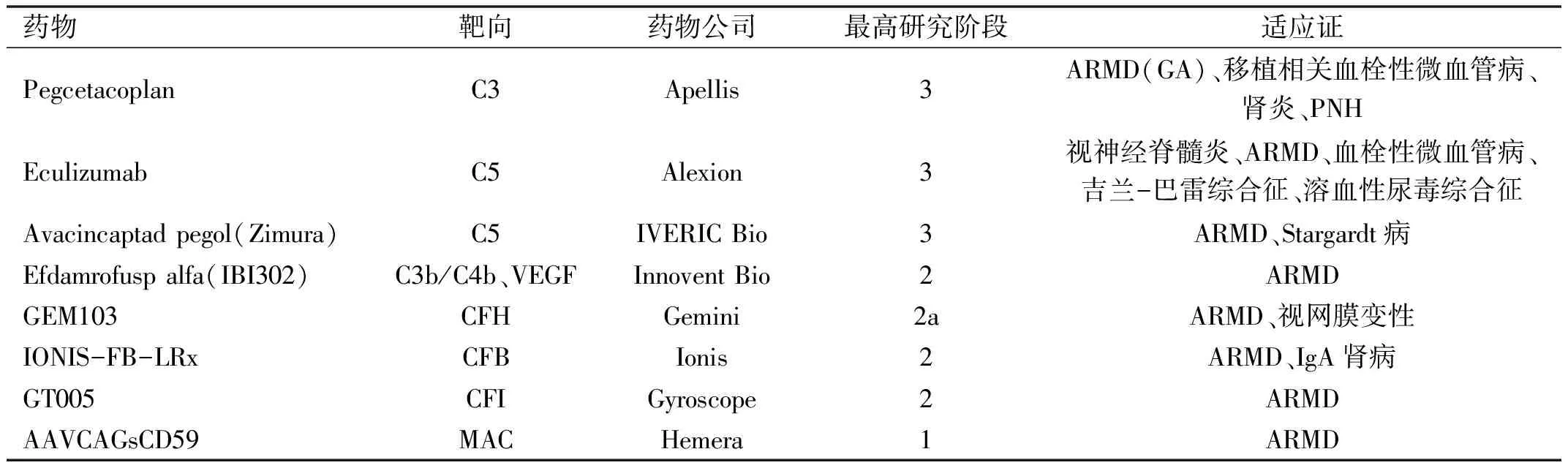
表1 常见的应用于眼科的靶向补体药物
4 小结与展望
DR是一种常见的严重危害眼健康的全球性疾病,了解其发病机制对于疾病的诊断和治疗有重大意义。综上所述,补体系统与DR的发生发展密切相关,但补体蛋白对DR的具体机制仍尚未完全阐明,期待未来相关研究能着重于各个补体蛋白在DR中的具体通路、详细机制等,从而在此基础上寻找更有效的治疗方案。截至目前,DR的治疗方案较少,包括抗VEGF药物、激光光凝等,且有希望的新疗法仍在研发中[57-58]。考虑到DR是全身疾病糖尿病进展的一部分,控制原发病应该是DR预防和治疗的重要前提。在新的治疗策略中,已有学者指出,相关补体成分可靶向作为包括DR在内的视网膜疾病的合适的预测因子[59],不仅如此,针对补体系统的靶向补体药物已在眼科其他相关疾病的治疗中取得了一定成效[44-45,49-50,56],这些研究和成果也将为DR的防治提供重要的参考和借鉴。
猜你喜欢
高中数理化(2025年3期)2025-02-15 00:00:00
保健医苑(2022年5期)2022-06-10 07:46:38
建材发展导向(2021年16期)2021-10-12 05:39:24
中国临床医学影像杂志(2021年6期)2021-08-14 02:21:56
昆明医科大学学报(2021年8期)2021-08-13 09:00:02
昆明医科大学学报(2021年2期)2021-03-29 07:42:24
肝博士(2020年5期)2021-01-18 02:50:18
中国医药指南(2017年3期)2017-11-13 02:55:32
安徽医科大学学报(2016年12期)2017-01-15 14:22:02
江西通信科技(2015年2期)2015-12-17 06:30:34
