
中药皂苷类化合物生物转化研究进展
2024-02-28 06:03:02李忆红梁雨璐解嘉琪黄建梅
中草药 2024年3期
李忆红,梁雨璐,解嘉琪,王 灿,黄建梅
北京中医药大学中药学院,北京 102488
皂苷是由皂苷元与糖基通过糖苷键连接而成的糖苷类化合物,在水溶液中可形成肥皂状泡沫,故得皂苷之名。皂苷类化合物广泛分布于陆生植物中,少量存在于海绵、海星、珊瑚等海洋生物中[1],皂苷按其苷元碳骨架结构可分为三萜和甾体皂苷。三萜皂苷在豆科(甘草、黄芪等)、五加科(人参、三七等)、桔梗科(桔梗等)、远志科(远志等)、伞形科(柴胡等)和石竹科等植物中分布较多;甾体皂苷大部分分布于百合科(知母、重楼等)、薯蓣科(薯蓣等)和玄参科等植物中。皂苷在中药中广泛存在,具有抗肿瘤、抗病毒、抗炎、抗菌、解热、镇静等生物活性[2]。与生物碱、黄酮等其他天然产物不同,皂苷原型成分口服后生物利用度相对较低,但通过生物转化可以获得生物利用度相对较高的次级皂苷和苷元,其生物学活性增强。研究发现人参皂苷药效活性与人参皂苷分子糖基数量有关,如稀有人参皂苷具有更高的药理活性,药动学性质优于原型人参皂苷[3-4],稀有人参皂苷与人参皂苷具有相同的母核结构,只是糖基的数量不同,说明修饰皂苷分子中的糖基部分,对提高其药理活性及生物利用度具有重要意义。
天然皂苷可以通过水解而获得次级皂苷及其衍生物、苷元,常见的方法有物理(加热)、化学(酸、碱水解)和生物转化等方法,其中生物转化包括肠道菌群转化、微生物转化和体外酶催化。酸水解反应条件剧烈,易使苷元发生脱水、双键移位和环合等结构变化[5-6],无法控制糖基的选择性水解。另外,利用酸碱水解皂苷的方法副反应产物较多,水解产物仍需进一步处理。因此,近年来皂苷成分转化生成次级皂苷及苷元的研究主要集中于生物转化领域,由于其反应条件温和,具有更强的位置选择性和立体选择性,被广泛应用于研究和生产中。生物转化主要反应类型包括糖基水解反应、氧化还原反应、羟基化及断裂重排反应等[7-8],其中以糖基水解反应应用最为广泛。本文结合国内外近年研究成果,对不同类型的皂苷化合物通过生物转化的方式水解糖基的文献进行整理,为后续皂苷定向修饰糖基的研发和利用的研究提供思路和参考。
1 三萜皂苷的生物转化
1.1 四环三萜
1.1.1 人参皂苷 人参皂苷根据其苷元骨架可分为达玛烷、奥克梯隆醇和齐墩果烷型,其中大多数人参皂苷母核结构属于达玛烷型[9],且是人参的主要活性成分,根据苷元羟基位置的不同进一步将达玛烷型人参皂苷分为原人参二醇(protopanaxadiol,PPD)和原人参三醇(protopanaxatriol,PPT)型,PPD 型人参皂苷的糖基位于达玛烷的C-3 和C-20位,PPT 型人参皂苷的糖基位于C-6 和C-20 位[10]。人参皂苷Rb1、Rb2、Rc、Rd、Rg1等皂苷侧链含有多个糖基,占人参总皂苷的80%以上,被认为是人参皂苷的主要成分[11-12]。相比之下,次级人参皂苷F2、CK、Rg3、Rh2占人参总皂苷的比例不到0.1%[13]。综合文献分析发现,人参皂苷的糖基水解反应主要发生在C-3、C-6 和C-20 位,水解脱去葡萄糖基、鼠李糖基、呋喃型阿拉伯糖基和吡喃型阿拉伯糖基。人参皂苷在体内主要的代谢途径为逐级脱糖基,将PPD 型人参皂苷Rd、F2、Rg3、CK、Rh2与人源肠道菌群离体孵育培养,发现人参皂苷Rd 的主要转化产物为人参皂苷F2、Rg3、Rh2、CK 和PPD;人参皂苷F2主要转化产物为人参皂苷CK 和PPD;人参皂苷Rg3主要转化产物为人参皂苷Rh2和PPD;而人参皂苷Rh2和人参皂苷CK 又可以生成PPD[14-15]。离体大鼠肠道菌群对人参皂苷Rb1的水解也是逐级脱糖基的过程[16]。人参总皂苷在体内肠道菌群中的代谢同离体肠道菌群一致,Guo 等[17]验证了肠道菌群转化三七皂苷的过程,结果发现在常规饲养大鼠血浆中检测到人参皂苷F1、Rh2、CK 3 个次级皂苷和PPT,而在无菌大鼠血浆中未检测到,因此推断人参皂苷体内的代谢基本规律为四糖皂苷→三糖皂苷→二糖皂苷→单糖皂苷→皂苷元[18]。
Chi 等[19]研究发现,多种微生物协同作用可以通过人参皂苷Rd→人参皂苷F1→人参皂苷CK的途径将人参皂苷Rb1转化为人参皂苷CK、Rh2;通过人参皂苷Re→人参皂苷Rg2、Rg2→人参皂苷Rh1的途径将人参皂苷Re 转化为人参皂苷Rh1。从韩国主食泡菜中提取的各种菌株,如LeuconostoccitreumLH1,可以很好地将PPD 型人参皂苷转化为人参皂苷CK[20]。Kim 等[21]筛选出12 种菌株,其中M.trithecenlyticum可将人参皂苷Rb1转化为人参皂苷Rh2。Zhou 等[22-23]研究表明,A.niger3.3883 和A.oryzae3.591 菌株的混合孢子液在7%海藻酸钠培养基中能够将人参皂苷Rb1转化为人参皂苷Rd、Rg3,利用Lactobacillusparacaseisubsp. tolerans,可以水解人参皂苷Rg3中C-3 位葡萄糖基,获得人参皂苷Rh2[24]。 Hu 等[25]利用来自Bifidobacterium adolescentisATCC15703 的β-葡萄糖苷酶研究人参皂苷的生物转化,以人参皂苷Rb1和Rd 为底物,人参皂苷Rb1的C-3 葡萄糖基部分水解生成绞股蓝皂苷-XVII(gypenoside-XVII,Gyp-ⅩⅤⅡ),人参皂苷Rd 脱去C-3 外侧葡萄糖基生成人参皂苷F2;人参皂苷Rb1脱去C-3 和C-20 外侧葡萄糖基水解生成Gyp-XVII 和人参皂苷Rd。
人参皂苷结构包括1~4 个糖苷键,常见的糖基有β-葡萄糖基、L-阿拉伯糖基和L-鼠李糖基。因此,通过使用特定糖苷酶(如α-L-呋喃型阿拉伯糖苷酶、α-L-吡喃型阿拉伯糖苷酶、β-葡萄糖苷酶或果胶酶)选择性水解不同位置的糖基,将人参皂苷水解制备次级人参皂苷是可行的[26]。Zhang 等[27]从枯草芽孢杆菌中克隆并优化了呋喃型阿拉伯糖苷酶基因,能特异性水解人参皂苷Rc 阿拉伯糖糖基,将其转化生成人参皂苷Rd,Kim 等[28]报道了一种重组酶α-L-吡喃型阿拉伯糖苷酶,通过选择性水解C-20 位的吡喃型阿拉伯糖基,可高效催化人参皂苷Rb2转化为人参皂苷Rd。α-鼠李糖苷酶能够水解PPT 型人参皂苷C-6 位的鼠李糖产生人参皂苷Rg1、Rh1[29],此外,Chang等[30]对来自AspergillusnigerKCCM 11239 的β-葡萄糖苷酶进行优化,该酶在人参皂苷Rb1的C-20 位水解葡萄糖基生成人参皂苷Rd 和Rg3,在C-3 位水解葡萄糖基生成人参皂苷F2,β-葡萄糖苷酶MT619 利用2 种转化途径将人参皂苷Rb1转化为次级人参皂苷 CK 和 PPD[31]:人参皂苷Rb1→Gyp-XVII→Gyp-LXXV→人参皂苷 CK→PPD;人参皂苷Rb1→Gyp-XVII→人参皂苷F2→人参皂苷CK→ PPD,果胶酶可将人参皂苷Rb1转化为人参皂苷Rd[32]。对人参皂苷转化途径及生物转化情况见图1。
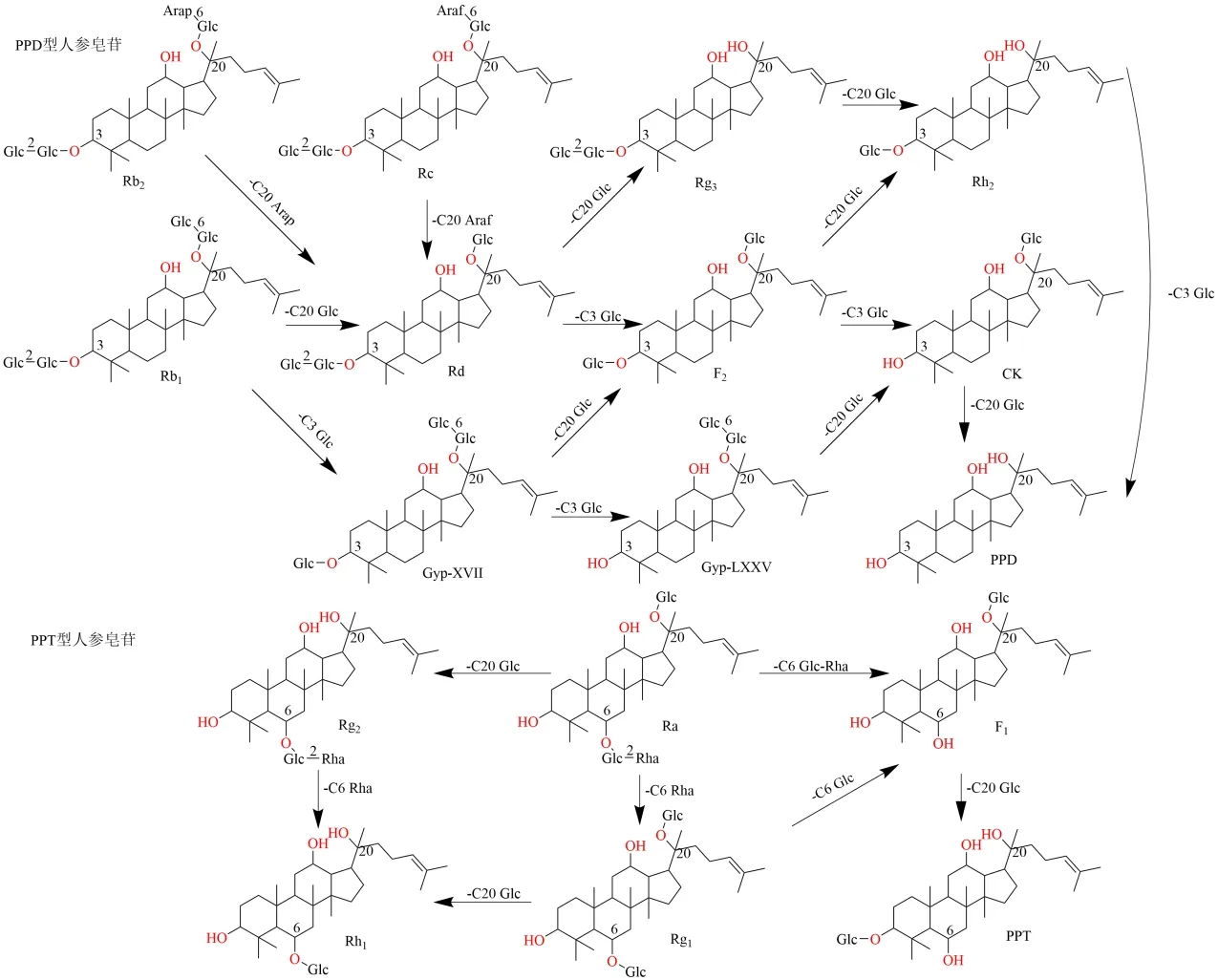
图1 人参皂苷的生物转化途径Fig.1 Biotransformation pathway of ginsenosides
1.1.2 Gyp 目前,Gyp 的生物转化研究部分和人参皂苷重合,如Gyp-XVII、LXXV 转化途径,Gyp生物转化途径见图2。Chen 等[33]研究发现给予大鼠Gyp-LVI 后,其在大鼠体内的主要转化代谢过程是脱糖基和脱水,5 个代谢物分别是Gyp-XLVI、Gyp-L、Gyp-LI、达木林B 和A。保加利亚乳杆菌对Gyp-XLVI 的主要转化物与大鼠胃肠道转化产物一致,分别为Gyp-L、Gyp-LI、达木林A 和B[34]。

图2 绞股蓝皂苷的生物转化途径Fig.2 Biotransformation pathway of gypenosides
利用高效液相色谱对比分析,发现Gyp 经微生物Aspergullusglaucus转化后,产生低极性的次级皂苷,陈良华等[35]利用微生物FFCDL-90 菌株得到的鼠李糖苷酶将Gyp-5 转化为人参皂苷Rd。以多糖基原型Gyp 为底物,筛选出高产Gyp 糖苷酶的菌株Absidasp. GYP4r,采用离子交换色谱法分离纯化得到Gyp 糖苷酶能够特异性水解Gyp,对pNP-α-D-Glc、pNP-β-D-Glc、pNP-α-D-Gal 均具有水解能力[36-38]。
通过使用FervidobateriumpennivoransDSM9078产生的重组酶将Gyp-XLIX 脱去C-21 位葡萄糖基水解转化生成gylongiposide I[39],Gyp-XLVI 通过柚皮苷酶转化生成Gyp-TN-1[40]。刘玥[41]利用NLZ-R和NLZ-X 特异性水解Gyp-NPLC0394 的鼠李糖糖基和木糖糖基,生成化合物A~C;利用蜗牛酶可以水解Gyp-LVI 和Gyp-XLVI 的葡萄糖基,生成2-羟基-3-O-β-D-吡喃葡萄糖基-(20S)-原人参二醇-20-O-[β-D-吡喃木糖基-(1→6)]-β-D-吡喃葡萄糖苷(I)、2-羟基-(20S)-原人参二醇-20-O-[β-D-吡喃木糖基-(1→6)]-β-D-吡喃葡萄糖苷(II)、2-羟基-3-O-β-D-吡喃葡萄糖-(20S)-原人参二醇-20-O-β-D-吡喃葡萄糖苷(III)、2-羟基-(20S)-原人参二醇-20-O-β-D-吡喃葡萄糖苷(IV)。
1.1.3 黄芪皂苷 黄芪皂苷是黄芪的主要药效成分,在黄芪研究中占有重要地位。目前关于黄芪皂苷生物转化的研究较少,但对其皂苷结构的修饰,如脱乙酰基、脱糖基途径的规律总结较充分。黄芪皂苷在肠道菌群中的主要转化途径包括脱糖基、脱乙酰基、脱氢等,共同转化产物为环黄芪醇(cycloastragenol,CA)。孟欣桐等[42]研究表明黄芪甲苷(astragaloside-IV,AS-IV)可经人源肠道菌群脱糖基后,转化成易吸收入血的CA,大鼠肠道菌也可将ASI 脱去C-3 和C-6 位的木糖基和葡萄糖基生成环黄芪醇-6-O-β-D-吡喃葡萄糖苷和CA[43]。AS-IV 与大鼠肠道菌群离体孵育4 h,脱去C-3 和C-6 位糖基,水解生成环黄芪醇-6-O-β-D-葡萄糖苷(brachyoside B,Bra B)、cyclogaleginoside B(Cyc B)、CA 和氧化型CA[44]。
刘晓会[45]从不同来源的霉菌中筛选到能够转化黄芪总皂苷生成 AS-IV 的菌种——Absidia corymbiferaAS2。采用微生物转化技术,以黄芪总皂苷作为底物,通过发酵转化提高AS-IV 含量,并在黄芪总皂苷中鉴定出4 个可以转化为AS-IV 的化合物,经结构鉴定分别为黄芪皂苷I(astragaloside I,ASI)、异ASI(isoASI)、ASII 和isoASII,进一步研究该菌株生物转化的机制发现,其所表达的乙酰酯酶可有效水解ASII 中C-3 位吡喃木糖基残基的乙酰基, 转化途径分别是 ASII→ASI 、isoASII→ASII→AS-IV、ASI→(ASII、isoASII)→ASIV、isoASI→ASII→AS-IV[46-47]。利用Penicillium canescens也可将黄芪总皂苷向AS-IV 转化,使AS-IV 含量显著升高,是未转化前的5.51 倍[48]。8种不同的菌株进行AS-IV 的生物转化研究,发现A.nigerM85 生物转化活性最强,转化后AS-IV 含量是未转化前的11.7 倍[49]。阮鸣[50]使用不同的药(食)用真菌发酵黄芪,发现均可以将ASI 转化成异AS-IV,推测其转化机制为菌株生长过程中所产生的木糖苷酶水解AS-IV C-3 位木糖基后将其转化成异AS-IV。Wang 等[51]利用Bacillussp. LG-502成功将ASI 转化为CA,经核磁和质谱分析,推测其对AS-IV 的转化途径与肠内菌转化相似,转化机制为Bacillussp. LG-502 分泌的葡萄糖苷酶和木糖苷酶,AS-IV 在活性更高的葡萄糖苷酶的作用下优先脱去C-6 位葡萄糖基,随后脱去C-3 位木糖基生成CA,黄芪皂苷类化合物的生物转化途径见图3。

图3 黄芪皂苷的生物转化途径Fig.3 Biotransformation pathway of astragalosides
Cheng 等[52]使用来自Trichodermareesei的2 种酶(β-木糖苷酶、β-葡萄糖苷酶)将AS-IV 转化生成CA。来源于Dictyoglomusthermophilum的β-葡萄糖苷酶和β-木糖苷酶也能将AS-IV 转化生成CA[53-54]。商品化的β-葡萄糖苷酶可以将黄芪甲苷C-3 位木糖基脱去生成Bra B[55]。从Absidasp. A3r、A84r、A9r、A8r、A38r、ARr 6 种株菌中筛选出了黄芪皂苷糖苷酶高产菌株,可以将多糖基的黄芪皂苷转化生成低糖基的次级皂苷[56-57],研究还发现,来自Absidiasp. A3r 的黄芪皂苷糖苷酶特异性强,底物为人参皂苷Re、ASI 和柴胡皂苷等均未发生脱糖基的水解反应,该酶只水解四环三萜环阿屯烷型的黄芪皂苷C-16 位的α-鼠李糖基[58]。
1.1.4 罗汉果皂苷类成分 Zhou 等[59]在ig 罗汉果皂苷V 于大鼠后,在血浆、尿液、胆汁和粪便样品中发现罗汉果皂苷V 的脱糖基代谢物,其转化是依次脱去不同数量和位置上的葡萄糖,李典鹏等[60]在大鼠体外代谢研究中得出相同结论。经人源肠道菌群孵育,罗汉果皂苷III 依次脱去C-3 位葡萄糖基和C-24 位龙胆二糖基水解生成次级皂苷罗汉果皂苷IIA1和皂苷元罗汉果醇[61]。
采用产高活性β-葡萄糖苷酶的青霉属真菌J-9对罗汉果总皂苷进行生物转化,转化生成次级皂苷产物的化学结构:罗汉果醇-24-O-β-D-吡喃葡萄糖基-(1→2)-β-D-吡喃葡萄糖苷(A)、B 为罗汉果皂苷IIIE(B)、11-酮-罗汉果醇-3-O-β-D-吡喃葡萄糖基-24-O-β-D-吡喃葡萄糖基-(1→2)-β-D-吡喃葡萄糖苷(C)[62-63]。罗汉果总皂苷经β-葡萄糖苷酶水解1 次的产物以三糖皂苷和四糖皂苷为主;水解2 次的产物经鉴定结构为C-3 或C-24 位连接1 个β-葡萄糖基的单糖皂苷及少量二糖皂苷为主[64],利用β-葡聚糖酶水解转化罗汉果皂苷V,生成连接不同数量糖基的罗汉果皂苷IIIE、III、IVE 和赛门苷I 等次级皂苷[65],生物转化途径见图4。
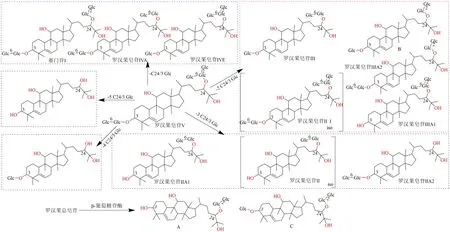
图4 罗汉果皂苷类成分的生物转化途径Fig.4 Biotransformation pathway of mogrosides
1.1.5 酸枣仁皂苷类成分 杜晨晖等[66]利用UPLC-Q-Orbitrap-MS 分析发现,酸枣仁皂苷类成分在正常人源肠道菌群中的主要转化途径为脱糖基反应,依次脱去皂苷元外侧糖基,脱糖基顺序不同会生成不同的转化产物。酸枣仁皂苷在大鼠肠道菌群中的转化途径与人源相似,均会产生及次生皂苷和酸枣仁皂苷元。酸枣仁皂苷A(jujuboside A,Ju A)被茯苓真菌中的葡萄糖苷酶水解脱去C-3 位葡萄糖基,生成Ju B[67],Ju A 在离体人源肠道菌群中也会转化生成为酸枣仁皂苷B[68-69],酸枣仁皂苷在肠道菌群和微生物中的转化途径见图5。Matsuda 等[70]通过酶解法将原酸枣仁皂苷A、B、B1 C-23 位的葡萄糖基脱去,转化生成为已知的酸枣仁皂苷。

图5 酸枣仁皂苷类成分的生物转化途径Fig.5 Biotransformation of jujubosides
1.2 五环三萜
1.2.1 甘草酸 多数甘草皂苷类成分均具有五环三萜结构母核,甘草酸是其中含量最高的成分,也是甘草皂苷类成分的代表性化合物。GL 为齐墩果烷型三萜皂苷,在C-3 位有2 分子葡萄糖醛酸基[71]。Akao 团队[72-74]阐明甘草酸的肠道菌群转化途径主要有2 种,见图6。(1)甘草酸1 次性水解脱去2个葡萄糖醛酸,直接生成甘草次酸;(2)先水解掉C-3 位末端的1 个葡萄糖醛酸基,生成中间体单葡萄糖醛酸甘草次酸( glycyrrhetinic acid monoglucuronide,GAMG),再水解脱去1 个葡萄糖醛酸生成甘草次酸。有研究发现无菌大鼠无法将甘草酸吸收入血且未发生脱糖基的转化反应,说明肠道菌的转化作用是将甘草皂苷类成分吸收入血的必要环节,随后研究团队从人源粪便中分离筛选到参与甘草酸转化的菌株,如Eubacteriumsp. GLH、Ruminococcussp. PO1-3 和Streptococcussp. LJ-22等,其中Streptococcussp. LJ-22 被证实可特异性地水解甘草酸C-3 位末端的1 个葡萄糖醛酸基生成GAMG[75]。Huang 等[76]将甘草提取物与人源肠道菌群体外孵育,研究结果表明甘草皂苷A3、E2、G2、22-β-乙酰氧基甘草酸等三萜皂苷的肠道菌群转化方式与甘草酸相同,均为脱糖基后生成次级皂苷及苷元。
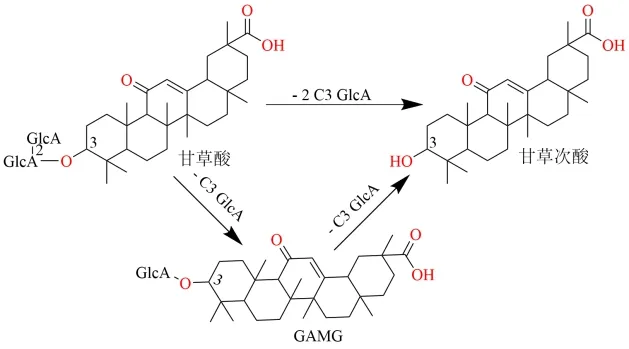
图6 甘草酸的生物转化途径Fig.6 Biotransformation pathway of glycyrrhizic acid
近些年,研究者对甘草酸的微生物转化的相关研究较多[77-79],发现了许多能成功转化甘草酸的菌种,但甘草酸的转化途径简单,与肠道菌群转化途径一致。用于微生物转化的菌株产β-D-葡萄糖醛酸苷酶,不同来源的酶对脱糖基的位置具有立体选择性, 从StreptococcusLJ-22[80]、Penicillium purpurogenumLi-3[81]和TalaromycespinophilusLi-93[82]中分离纯化的葡萄糖醛酸苷酶仅水解甘草酸C-3 位末端葡萄糖醛酸基生成GAMG,水解生成甘草次酸。来源于Aspergillusniger[83]和EubacteriumL-8[84]的葡萄糖醛酸苷酶可将甘草酸彻底水解生成甘草次酸。
1.2.2 柴胡皂苷类成分 Shimizu 等[85]研究表明柴胡皂苷A、B1、B2、D、G 等在大鼠肠道中主要发生脱糖基反应,首先脱去C-3 位末端的葡萄糖基生成次级苷原柴胡皂苷F、A、H、G、D,然后脱去C-3位岩藻糖生成皂苷元F、A、H、G、D,Kida 等[86]将柴胡皂苷类成分与人粪便共同培养后,也得到相同的结论,并从人源肠内菌株中筛选出Eubacteriumsp. A-44,研究该菌株参与生物转化的机制。研究发现从E. sp. A-44 中分离得到的酶(β-葡萄糖苷酶和β-岩藻糖苷酶)可逐级脱去柴胡皂苷C-3 位的岩藻糖和葡萄糖,最终生成柴胡皂苷元。Yu 等[87]发现柴胡皂苷C 在人源肠道菌群中的转化途径与柴胡皂苷A、B1、B2、D、G 类似,在体内均为逐步脱糖基的转化。国内学者研究发现柴胡皂苷A 经大鼠肠道菌群转化生成柴胡次皂苷F 和柴胡皂苷元F[88],进一步证明了柴胡皂苷在肠内菌的转化途径。
徐金丽和富瑶瑶发现sp.c42、Microbacteriumsp.GS514-saiko 和Rhodanobactersp.GS3054 所产的酶都能将柴胡皂苷转化为低糖基的次级皂苷[89-90]。重组酶ABQ 与柴胡皂苷A 反应使C-3 位的葡萄糖水解生成柴胡次皂苷F,继续水解C-3 位的岩藻糖生成柴胡皂苷元F[91],纤维素酶可以将柴胡皂苷B2脱糖基生成前柴胡皂苷D,蜗牛酶可以将柴胡皂苷B1、A、D 脱糖基分别生成前柴胡皂苷A、F、G[92],柴胡皂苷的生物转化途径见图7。

图7 柴胡皂苷类成分的生物转化途径Fig.7 Biotransformation pathway of saikosaponins
1.2.3 白头翁皂苷类成分 白头翁皂苷 B4(anemoside B4,B4)与不同肠段的肠道菌群孵育共鉴定得到10 个代谢物,其中9 个产物均是脱糖基生成,B4 母核为2,3-羟基白桦脂酸皂苷元,含有5个糖基,在肠道菌群中逐级脱糖基,生成含有2~4个不同数量糖基的次级皂苷及2,3-羟基白桦酸[93]。白头翁皂苷D、B7 和BD 在体外肠道菌群中也发生类似的脱糖基反应,得到次级皂苷及常春藤皂苷元[94-95]。
当前,对于白头翁皂苷糖苷酶的生物转化已有大量研究,并从中分离出多种能转化白头翁皂苷的糖苷酶[96-97],利用来自Absidiasp.P00r 的酶,将六糖基白头翁皂苷转化生成含有2 个糖基的次级皂苷常春藤皂苷元-3-O-L-吡喃鼠李糖基-(1→2)-L-吡喃阿拉伯糖苷[98]。利用菌株Absidiasp.P39r 生成的水解酶,水解白头翁皂苷H C-28 位末端鼠李糖基,得到白头翁皂苷PS-I;C-3 和C-28 位分别脱去葡萄糖基,得到白头翁皂苷PS-II;进一步水解脱去C-28位葡萄糖基,得到白头翁皂苷A,白头翁皂苷在肠内菌及其酶的转化见图8。

图8 白头翁皂苷类成分的生物转化途径Fig.8 Biotransformation pathway of anemosides
1.2.4 走马胎皂苷类成分 走马胎为紫金牛科植物,九节龙皂苷-I(ardipusilloside-I,ADS-I)和走马胎皂苷Ag3是该药材中2 个代表性的皂苷成分。体外模拟人肠道菌群对ADS-I 进行生物转化研究,初步推断出M1~M4 4 个代谢产物的结构[99]。其中代谢产物M1、M2、M4 与ADS-I 在大鼠肠内菌的转化产物相同[100],均为ADS-I 分别脱去C-3 位葡萄糖基(M1)、葡萄糖基和鼠李糖基(M2)及C-3 位全部糖基脱去后生成的苷元(M4)。脱去3 个糖基的M3 是人体肠道菌独有的转化产物,脱去C-3 位末端鼠李糖基的M5 是大鼠肠道菌独有的转化产物,提示人和动物菌群的组成不同,ADS-I 的生物转化产物也会发生变化。
张静[101]在内生菌和土壤菌种筛选得到3 株对走马胎皂苷Ag3具有转化能力的菌株,分别是SphingomonasyabuuchiaeGTC868T(AB071955)、BacilluslicheniformisATCC14580T(AE017333)和BacillusasahiiMA001T(AB109209)。该团队也发现果胶酶也可以转化走马胎皂苷Ag3生成次级皂苷A-1、A-2、A-3 及苷元。ADS-I 和走马胎皂苷Ag3生物转化途径见图9。
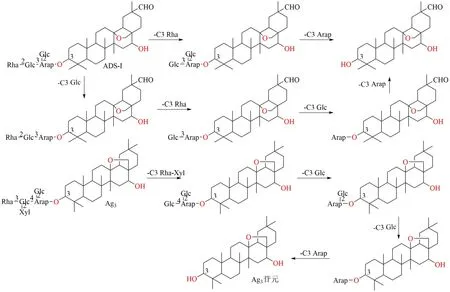
图9 ADS-I 和Ag3 生物转化途径Fig.9 Biotransformation pathway of ADS-I and Ag3
1.2.5 其他五环三萜类皂苷 桔梗皂苷肠内菌的转化途径主要是脱去C-3 位糖基[102]。经黑曲霉与黑根霉转化后桔梗总皂苷的色谱峰明显后移,说明皂苷糖基脱去生成次级皂苷或苷元[103]。Shin 等[104-105]发现耐酸性果胶酶Cytolase PCL5 可以水解桔梗皂苷E C-3 位葡萄糖基和C-28 位的木糖基和芹糖基,具有底物特异性,Cytolase PCL5 是一种混合酶,因此可能对多种糖基具有特异性。同样,来自黑曲霉的Pluszyme 2000P 仅水解桔梗皂苷C-3 位葡萄糖基[106]。利用蜗牛酶水解桔梗总皂苷[107],可以将结构相近的桔梗皂苷E、D3 等转化为桔梗皂苷D;从Trichodermareesei、Aspergillusoryzae[108]、AspergillususamiiKFRI 1004[109]、P.grandiflorus[110]和Aspergillusaculeatus[111]分离纯化得到的纤维素酶、β-半乳糖苷酶、β-葡萄糖苷酶和果胶酶均能将桔梗皂苷E 和D3 转化为桔梗皂苷D。
商陆的主要活性成分之一为商陆皂苷甲(esculentoside A,EsA),研究发现EsA 具有抗炎、抗菌和利尿等多种活性[112],但EsA 的溶血性限制了其应用。EsA 经生物转化脱糖基后,生成EsB,该化合物溶血性降低,增强了抗炎活性[113-114]。EsA经微生物StreptomycesgriseusATCC 13273 脱去C-3 位木糖基和葡萄糖基,生成EsB 和苷元-商陆酸甲酯[115]。
重组糖苷酶和蜗牛酶与EsA 反应均使C-3 位末端葡萄糖基被水解,生成 EsB[91],蜗牛酶在48.28 ℃、pH 值为6.4 时产率最高[116]。赖道万[117]使用黑曲霉β-葡萄糖苷酶、苦杏仁酶、蜗牛酶等酶也能使EsA 中的葡萄糖基发生水解,生成EsB,其中只有蜗牛酶能使EsA C-3 位糖基侧链完全水解,得到其皂苷元。
Watanabe 等[118-119]从AspergillusoryzaePF1224、N.vasinfectavar.vasinfectaP F1225 和EupenicilliumbrefeldianumPF1226 中纯化出大豆皂苷水解酶,显示该酶不仅可以水解大豆皂苷I 的C-3位连接的鼠李糖基和半乳糖基,还可以进一步水解其内端的葡萄糖醛酸基生成大豆甾醇B。Abdel 等[120]筛选了5 种菌株,发现A.terreusII 能将大豆总皂苷最大效率地转化为大豆甾醇B。田晶等[121]利用8 种霉菌菌株对大豆皂苷糖基的水解能力进行探讨,得到米曲霉A.oryzae39s、A.oryzaeslows 和黑曲霉A.niger848s 3 种具有较高活性的菌种,均使大豆皂苷部分糖基水解生成低糖基皂苷及苷元。
牛膝三萜皂苷和大鼠肠道菌群共培养,产生39个转化产物,结果显示牛膝皂苷在肠内的主要转化途径是去糖基化,竹节参皂苷V、IV、IVa 等原型皂苷的转化产物为姜状三七皂苷R1、28-去葡萄糖基竹节参皂苷IVa 和齐墩果酸等。根据产物特征碎片推断,脱糖基反应主要发生在C-3 和C-28 位的糖苷键[122]。
在人参皂苷、黄芪皂苷和甘草皂苷等三萜皂苷的生物转化中,反应机制主要为脱糖基的水解反应,通常发生在C-3、C-6、C-20、C-21、C-23、C-24 和C-28 位。转化生成的次级皂苷和苷元不仅可以提高生物利用度,也能增强抗炎、抗病毒和抗癌等药效[79,123]。
2 甾体皂苷的生物转化
2.1 薯蓣皂苷
薯蓣皂苷在人源肠道菌群中糖基被逐个水解,形成相应的次级皂苷和薯蓣皂苷元[8,124]。Tang等[125]表征了给药后大鼠体内总薯蓣皂苷、薯蓣皂苷和薯蓣皂苷元的代谢轮廓,结果表明薯蓣皂苷的逐级脱糖基是最重要的转化过程。
Chen 等[126]利用Lactobacilluscasei脱去薯蓣皂苷C-3 位糖基生成薯蓣皂苷元,Liu 等[127]从米曲霉Aspergillusoryzae中分离纯化的水解酶,可以水解原薯蓣皂苷中C-3 位末端α-1,4-鼠李糖基和C-26 位葡萄糖基生成薯蓣次皂苷 A。从新月弯孢霉Curvularialunata中分离纯化出的α-L-鼠李糖苷酶能够水解C-3 位末端α-1,2-鼠李糖基,生成progenin V[128]。从新鲜牛肝中分离得到的鼠李糖苷酶能够水解2 个鼠李糖基生成C-3 位连有1 个葡萄糖基的次级薯蓣皂苷[129]。Jin 等[130]则从Absidiasp. d38 中分离纯化出了薯蓣皂苷水解酶,可将薯蓣皂苷侧链糖基全部脱去,水解生成薯蓣皂苷元。
2.2 知母皂苷B-II/III
在大鼠肠道菌群中,知母皂苷B-II 和B-III 发生的转化反应主要有脱糖基化、脱水、羟基化、E环裂解和氧化反应[7,131-133],脱糖基是我们重点关注的途径,转化途径为知母皂苷B-II 的C-3 位依次脱去葡萄糖基及半乳糖基,得到次级知母皂苷B-II-a和B-II-b;C-26 位脱去葡萄糖基,生成知母皂苷A-III;知母皂苷B-III 的C-3 位脱去葡萄糖基及半乳糖基,得到次级知母皂苷B-III-a、B-III-b。利用微生物AspergillusnigerAS 3.0739[134]和Colletotrichumgloeosporioideis[135]转化知母皂苷B-II,均会水解脱去C-3 和C-26 位糖基,得到次级皂苷。
甾体皂苷的生物转化中,大多以皂苷与皂苷元之间的相互转化为主,脱糖基水解反应主要发生在C-3 和C-26 位。与三萜皂苷类化合物相比,甾体皂苷的生物转化机制更简单,涉及到的反应类型少。
3 结语与展望
以几种具有代表性的三萜皂苷和甾体皂苷为例,综述了中药中皂苷类成分生物转化研究进展。其中,肠道菌群种类多样,逐级脱糖基是最主要的转化方式,即生成一系列次级皂苷及对应的皂苷元。另外,肠内菌还可以使皂苷发生羟基化、氧化和去甲基化等反应,是次要的肠内菌转化途径。因人体肠道菌群与模型动物在物种组成及多样性上的差异,皂苷的肠内转化也存在物种差异。如ADS-I 在人和大鼠肠道菌群中转化产物部分一致[99-100]。从微生物中筛选合适的菌株或酶进行生物转化,是制备次级皂苷的有效途径。大多数生物转化研究所选用的菌株主要包括4 个来源:(1)从人源或大鼠的肠道菌群中筛选出能够转化皂苷的优良菌株;(2)从产生糖苷酶的已知菌株中筛选出转化特异性高的优良菌株[79];(3)从药用植物生长土壤中筛选出具有转化优势的优良菌株;(4)从植物内生菌中筛选优良菌株。随着酶工程和基因工程研究的发展,部分课题组储有用于生物转化的特异性纯化酶或重组酶,若能将特异性高的糖苷酶转化为产品,实现水解酶的工业化生产,对皂苷类化合物的生物转化研究和经济效益具有深远意义,以满足后续成药性研究和新药开发的需求。
利益冲突所有作者均声明不存在利益冲突
猜你喜欢
食品与生物技术学报(2022年1期)2023-01-11 09:11:11
宁夏医学杂志(2020年3期)2021-01-21 08:23:38
空间科学学报(2020年4期)2020-04-22 01:17:16
中成药(2018年5期)2018-06-06 03:11:58
中成药(2018年4期)2018-04-26 07:12:45
中成药(2017年8期)2017-11-22 03:19:32
天然产物研究与开发(2016年1期)2016-06-05 10:29:25
首都医科大学学报(2015年4期)2015-12-16 13:00:08
应用化工(2014年9期)2014-08-10 14:05:08
天然产物研究与开发(2014年3期)2014-04-27 14:15:35
