
Characterization, expression dynamics, and potential function of OPA1 for regulation of mitochondrial morphology during spermiogenesis in Phascolosoma esculenta*
2024-02-27 08:28XinmingGAOBinbinFENGChenDUCongcongHOUShanJINDaojunTANGJunquanZHU
Xinming GAO, Binbin FENG, Chen DU, Congcong HOU, Shan JIN,Daojun TANG,**, Junquan ZHU,**
1 Key Laboratory of Applied Marine Biotechnology by the Ministry of Education and Key Laboratory of Marine Biotechnology of Zhejiang Province, School of Marine Sciences, Ningbo University, Ningbo 315211, China
2 College of Ecology, Lishui University, Lishui 323000, China
Abstract Mitochondria undergo morphological changes during spermatogenesis in some animals.The mechanism and role of mitochondrial morphology regulation, however, remain somewhat unclear.In this study, we analyzed the molecular characteristics, expression dynamics and subcellular localization of optic atrophy protein 1 (OPA1), a mitochondrial fusion and cristae maintenance-related protein, to reveal the possible regulatory mechanisms underlying mitochondrial morphology in Phascolosoma esculenta spermiogenesis.The full-length cDNA of the P.esculenta opa1 gene (Pe-opa1) is 3 743 bp in length and encodes 975 amino acids.The Pe-OPA1 protein is highly conservative and includes a transmembrane domain, a GTPase domain, two helical bundle domains, and a lipid-interacting stalk.Gene and protein expression was higher in the coelomic fluid (a site of spermatid development) of male P.esculenta and increased first and then decreased from March to December.Moreover, their expression during the breeding stage was significantly higher than during the non-breeding stage, suggesting that Pe-OPA1 is involved in P.esculenta reproduction.The Pe-OPA1 protein was more abundant in components consisting of many spermatids than in components without, indicating that Pe-OPA1 mainly plays a role in the spermatid in coelomic fluid.Moreover, Pe-OPA1 was mainly detected in the spermatid mitochondria.Immunofluorescence experiments showed that the Pe-OPA1 are constitutively expressed and co-localized with mitochondria during spermiogenesis, suggesting its involvement in P.esculenta spermiogenesis.These results provide evidence for Pe-OPA1’s involvement in the regulation of mitochondrial morphology during spermiogenesis.
Keyword: optic atrophy protein 1 (OPA1); spermiogenesis; Phascolosoma esculenta; mitochondrial dynamics; spermatid
1 INTRODUCTION
Mitochondria are bilayer membrane organelles of variable shape (e.g., spherical, elliptical, tubular, and irregular, among others).Mitochondria are the sites of the tricarboxylic acid cycle and oxidative phosphorylation and play an important role in cell survival, metabolism, and signaling (Jeong and Seol, 2008; Breitenbach et al., 2014; Akbari et al.,2019; Abate et al., 2020).In addition, mitochondria play important roles in spermatogenesis (Vertika et al., 2020), a process of spermatogonia developing into sperm.Spermatogenesis comprises the multiplication and differentiation of spermatogonia,meiosis of spermatocytes, and spermiogenesis, wherein the spermatid develops into sperm.Abnormal mitochondrial function affects the progression of spermatogenesis and therefore affects male fertility,sometimes leading to infertility (Nakada et al.,2006; Rajender et al., 2010; Baklouti-Gargouri et al.,2014; Jiang et al., 2017).One of the characteristics of mitochondria in spermatogenesis is mitochondrial morphology, which changes in shape, size, and cristal conformation.For example, inPelodiscus sinensis, mitochondria are round in the early spermatid,elongated in the granular nucleus spermatid, swollen and crescent-shaped in the compact nucleus spermatid,round in the late spermatid, and onion-like in the mature sperm (Haseeb et al., 2018).Moreover,mitochondrial size decreased in the granular nucleus spermatid and then increased in subsequent developmental stages (Haseeb et al., 2018).In addition, the shape of mitochondrial cristae from septa developing into concentric rings during spermiogenesis (Haseeb et al., 2018).In rats (Rattus norvegicus), mitochondria change from orthodox form to condensed form and then from the condensed form to an orthodox-like form during spermatogenesis(De Martino et al., 1979).Although morphological changes of mitochondria in spermatogenesis have been widely reported, the mechanism and role of mitochondrial morphology regulation during animal spermatogenesis remain somewhat unclear.
It has been reported that mitochondrial dynamics(mitochondrial fusion and fission) model the mitochondrial shape and size (Huang et al., 2011;Pernas and Scorrano, 2016).Mitochondrial fusion increases mitochondrial length and size, while mitochondrial fission does the opposite (Huang et al., 2011; Pernas and Scorrano, 2016).Mitochondrial dynamics, therefore, may be a mechanism regulating mitochondrial morphology during spermatogenesis.Indeed, researchers have found that mitochondrial fusion and fission-related proteins can affect mitochondrial morphology in spermatogenic cells,which affects spermatogenesis and male fertility(Varuzhanyan et al., 2019, 2021a, b).
The optic atrophy protein 1 (OPA1), belonging to the dynamin superfamily, is involved in mitochondrial fusion by changing its own conformation through GTP hydrolysis (Tilokani et al., 2018; Gao and Hu, 2021).The OPA1 protein contains a mitochondrial targeting signal, a transmembrane domain, a GTPase domain, two helical bundle domains, and a lipid-interacting stalk(Yapa et al., 2021).The OPA1 protein enters the mitochondrial membrane space using its mitochondrial targeting signal.Once there, peptidases process the protein to form long OPA1 isoforms (L-OPA1; Del Dotto et al., 2018).The L-OPA1 isoform can localize to the inner mitochondrial membrane(IMM) to mediate IMM fusion (Del Dotto et al.,2018; Tilokani et al., 2018; Gao and Hu, 2021).Knockdown of theopa1gene or disrupting L-OPA1 hydrolysis to short OPA1 isoform (S-OPA1), which disrupts its ability to localize to the IMM, resulted in mitochondria being unable to fuse and subsequently fragmenting into smaller sizes (Olichon et al., 2003;Song et al., 2009; Del Dotto et al., 2018).The OPA1 is also involved in the formation and maintenance of mitochondrial cristae.Lake of OPA1 or L-OPA1 hydrolysis to S-OPA1 leads to a reduction or even disappearance of mitochondrial cristae (Olichon et al., 2003; Griparic et al., 2004; Del Dotto et al., 2018).The important role of OPA1 on mitochondrial fusion and cristae maintenance in somatic cells implies that it is potentially involved in the regulation of mitochondrial morphology by affecting mitochondrial fusion and cristae shape in spermatogenic cells during spermatogenesis.
In the study, we analyzed the expression of OPA1 in different tissues, during the breeding cycle, in coelomic fluid components, and during spermiogenesis inPhascolosomaesculenta.This peanut worm species is a member of the Sipuncula phylum and is an economically important marine species for aquaculture industry at the southeast coast of China.It was also deemed an excellent model for studying spermiogenesis in oceanic invertebrates (Gao et al.,2019).In addition, mitochondria undergo morphology changes during its spermiogenesis (Gao et al.,2022).Our results revealed that OPA1 is an important putative protein that regulates mitochondrial morphology by participating in mitochondrial fusion and modeling cristae morphology during spermiogenesis and, therefore, may provide insight into the mechanism of mitochondrial plasticity during animal spermatogenesis.Additionally, the results provide basic data for the reproductive biology ofP.esculenta, which helps to improve its artificial breeding technology.
2 MATERIAL AND METHOD
2.1 Animal and tissue
Phascolosomaesculentaworms were sampled between March and December 2020 from the intertidal zone of the Xizhou of Ningbo city (China).Coelomic fluids, intestines, retractor muscles, nephridia,and body walls were collected monthly from 12 males and immediately snap frozen in liquid nitrogen.The samples were then stored at -80 °C until processing.In July, the collected worms were treated with CCCP (cyanogen-3-chlorobenyl; an oxidative phosphorylation uncoupler) by injecting it into the coelomic fluid at a concentration of 12.5 nmol/g.Following treatment for 1 h, the coelomic fluid was collected.Worms obtained between July and September (the breeding season ofP.esculenta)were used for coelomic fluid component separation.Briefly, the coelomic fluids were collected and centrifuged at 3 000×gat 4 °C for 2 min and washed twice with PBS.The components of each cell layer were then identified by micro-examination and stored at -80 °C.For the spermatid components, the mitochondria and cytoplasms were separated by removing the mitochondria using a tissue mitochondria isolation kit (Beyotime, Shanghai, China).To obtain spermatids at different stages, spermiogenesis ofP.esculentawas artificially induced in August,according to Zhu et al.(2007).The coelomic fluids were fixed in 4% (m/v) paraformaldehyde buffer before and after artificial induction for immunofluorescence experiments.
2.2 Full-length cDNA cloning of Pe-opa1
2.2.1 Primer design
The coding sequence of a homologousopa1gene inMizuhopectenyessoensis(Sequence ID: XM_021516852.1) was used for a BLAST search against the transcriptional database of sipunculid (Sequence Read Archive number are SRR1646442, SRR1646260,and SRR619011) in the National Center for Biotechnology Information (NCBI) to obtain homologous segments ofopa1in sipunculid species.The homologous segments were aligned using Vector NTI 11.5 to find the conservative segments.Primer Premier 5.0 was then used to design primers based on the conservative segments to clone the intermediate segment sequence.After cloning the intermediate segment sequence, primers were designed for rapid amplification of cDNA ends(RACE).All of the primers used in this study are listed in Supplementary Table S1.
2.2.2 Total RNA extraction and cDNA synthesis
Total RNA was extracted from the coelomic fluid using the TRIzol Reagent (Cwbio, Beijing, China).For cloning the intermediate segment sequences,cDNA was obtained by reverse transcription using the PrimeScript®RT reagent Kit (TaKaRa, China).For RACE, the cDNA was obtained by reverse transcription using a Smart RACE cDNA Amplification Kit (TaKaRa, China).
2.2.3 Full-length cDNA cloning
The PCR reaction mixture comprised 10 µL of 2×Taq MasterMix (Cwbio, Beijing, China), 7 µL of ddH2O, 1 µL of cDNA, and 1 μL of each primer(10 μmol/L).For cloning the intermediate segment,the cycling conditions were as follows: 94 °C for 5 min; 35 cycles of 94 °C for 30 s, 55 °C for 30 s,and 72 °C for 90 s; and 72 °C for 5 min.For RACE,the cycling conditions were as follows: 94 °C for 5 min; 8 cycles of 94 °C for 30 s, 69.5 °C for 30 s(decreased by 0.5 °C/cycle), and 72 °C for 90 s; 27 cycles of 94 °C for 30 s, 65.5 °C for 30 s, and 72 °C for 90 s; and 72 °C for 5 min.The resulting amplicons were separated using agarose gel electrophoresis,purified, and sequenced according to Zhang et al.(2017).Finally, the intermediate segment sequences were stitched to the cDNA sequences from RACE to obtain the full-length cDNA ofopa1.
2.3 Bioinformatics analyses of the Pe-opa1 gene and Pe-OPA1 protein
Identification of the open reading frame (ORF),prediction of primary structure, and determination of the molecular weight and isoelectric point ofPe-OPA1 were carried out using BioXM 2.6.The tertiary structure and GTP binding sites were predicted using the following online tools:https://zhanglab.ccmb.med.umich.edu/I-TASSER/ and https://blast.ncbi.nlm.nih.gov/Blast.cgi.Multiple sequence alignments and evolutionary tree analyses were carried out using Vector NTI 11.5 and MEGA 5.1,respectively.The structural domain ofPe-OPA1 was predicted by referencing the structural domains in homologous proteins inSaccharomycescerevisiae(Yan et al., 2020) and the predicted tertiary structure ofPe-OPA1.The amino acid (aa) sequences of thePe-OPA1 homologous proteins used in this study were downloaded from NCBI, and their GenBank accession numbers are listed in Supplementary Table S2.
2.4 Quantitative analysis of Pe-opa1 expression
A real-time quantitative PCR (qPCR) was conducted to analyze the expression ofPe-opa1.The qPCR primers were designed using Primer Premier 5.0 and are listed in Supplementary Table S1.Thegapdhgene was used as the internal control.Total RNA was extracted, and cDNA was synthesized,as before, from coelomic fluids, body walls, intestines,retractor muscles, and nephridia.The reaction mixture for the qPCR consisted of 10-μL Master Mix, 5 μL of 1:20 diluted cDNA, 3 μL of PCRgrade water, and 1 μL of each primer (10 μmol/L).The qPCR was carried out in the Roche LightCycler480 System, with the following cycling conditions: 95 °C for 3 min; 40 cycles of 95 °C for 10 s, 55 °C for 15 s, and 72 °C for 15 s.The relative expression levels ofPe-opa1were calculated using the 2-ΔΔCtmethod.Five to six samples were used for each experiment.
2.5 Western blotting
Total proteins were extracted from the various samples using the RIPA Lysis Buffer (Beyotime,Shanghai, China).Its concentrations were measured using the BCA protein assay kit (Beyotime,Shanghai, China).The protein samples, then, were denatured at 100 °C for 8 min in loading buffer(Beyotime, Shanghai, China).
Western blotting was carried out according to Zhang et al.(2017).The protein samples were separated using 4%-15% SDS-PAGE gels (ACE biotechnology, China) and transferred to a PVDF membrane (Solarbio, Beijing, China) using wet transfer.The membranes were blocked in 5% (w/v)dried skimmed milk in TBST (Solarbio, Beijing,China) for 2 h, washed two times in PBST, and incubated in primary antibody solution at 4 °C for 12 h.Next, the membranes were washed three times in TBST and incubated in second antibody solution.The membranes were washed as before and incubated with the developer solution.Finally, the membranes were photographed using a chemiluminescence imaging system (Tanon, Shanghai, China).The grey levels of protein blotting measured by using ImageJ software were used to analyze the relative expression of the OPA1 protein.Three samples were used for each experiment.
The primary antibodies used in this study included β-Actin (ACTB) Rabbit Monoclonal Antibody (1:500 dilution; Beyotime, Shanghai,China) and rat anti-Pe-OPA1 antibodies (1:500 dilution).The second antibodies included HRPlabeled Goat Anti-Rabbit IgG(H+L) (Beyotime,Shanghai, China) and HRP-labeled Goat Anti-Rat IgG (H+L) (Beyotime, Shanghai, China).The rat anti-Pe-OPA1 antibodies were in-house generated antibodies and obtained based on Gao et al.(2019).The immunogens forPe-OPA1 antibodies included 335F-565C (OPA1-1) and 672K-882F (OPA1-2).The recombinantPe-OPA1 peptides were obtained using prokaryotic expression and protein purification(Supplementary Fig.S1).After animal immunization using the purified peptides, the rat anti-OPA1 antibodies were obtained (Supplementary Fig.S1).
2.6 Statistical analysis
Data were analyzed using SPSS 21.0 (SPSS, Inc.,Chicago, IL, USA).The significance of differences was determined via one-way analysis of variance(ANOVA) for normally distributed datasets with homogeneity of variance or determined using a nonparametric test if datasets did not meet the above criteria.Data withP<0.05 were considered significant.
2.7 Immunofluorescence
Coelomic fluid samples were fixed in 4%paraformaldehyde at 4 °C for 2 h, washed thrice in PBS for 30 min, and osmosed in 0.5-mol/L sucrose.Next, the samples were embedded in optimal cutting temperature compound (Sakura, Torrance, CA,USA) and frozen at -80 °C to obtain embedding blocks.The blocks were cut using an HM525NX sliding microtome (Thermo Fisher Scientific, Waltham,MA, USA) to obtain 5-μm thick frozen sections.The sections were stored at -80 °C prior to their use.
The frozen sections were dried for 10 min at 20 °C, permeated in 0.3% PBST for 15 min, blocked in 5% (w/v) bovine serum albumin (BSA) for 1.5 h,and then incubated in a mixed primary antibody solution (rabbit anti-SDHA and rat anti-OPA1; 1:75 dilution) for 12 h at 4 °C.The sections were washed thrice in 0.1% PBST for 45 min and incubated in a mixed secondary antibody solution (Alexa Fluor 488-labeled Goat Anti-Rabbit IgG (H+L) and Cy3-labeled goat anti-rat IgG (H+L); 1:500 dilution) for 1.5 h at 25 °C.Next, the sections were washed six times in 0.1% PBST for 90 min and stained with diamidino-phenyl-indole buffer (DAPI) for 5 min.The sections were observed and photographed using a Zeiss laser scanning confocal microscope(LSM880, Carl Zeiss, Jena, Germany).The antibodies and DAPI solution used for immunofluorescence were purchased from Beyotime, China.
3 RESULT
3.1 Characteristics of the Pe-opa1 cDNA and the Pe-OPA1 protein
The full-length cDNA ofPe-opa1is 3 743-bp(GenBank accession number: OP067229), with a 40.37% GC content.The sequence comprises a 92-bp 5′ non-coding region, a 726-bp 3′ non-coding region, and a 2 925-bp open reading frame, encoding 975 aa (Fig.1).The molecular weight ofPe-OPA1 is about 112.66 kDa with a theoretical isoelectric point of 8.80.ThePe-OPA1 protein contains a transmembrane domain, a GTPase domain, two helical bundle domains (HB1 and HB2), and a lipidinteracting stalk (Fig.2).Within the GTPase domain,there are 19 GTP binding sites, which are clustered in a close region in the tertiary structure ofPe-OPA1(Supplementary Fig.S2).

Fig.1 Full-length cDNA of Pe-opa1
The consensus and identity positions ofPe-OPA1 with its homologous proteins inHomosapiens,Gallusgallus,Xenopustropicalis,Daniorerio,Drosophilamelanogaster,Pomaceacanaliculate,andCaenorhabditiseleganswere 65.6% and 53.7%,65.5% and 53.4%, 66.5% and 53.0%, 65.4% and 52.2%, 65.8% and 51.0%, 69.8% and 57.5%, and 63.0% and 47.4%, respectively (Fig.3).Moreover,there are high consensus positions between OPA1 in vertebrates and vertebrates, vertebrates and invertebrates, and invertebrates and invertebrates(Fig.4), suggesting that OPA1 is highly homologous between species and is conserved during animal evolution.Finally, the evolutionary relationship of OPA1 was consistent with the evolutionary relationship among the species (Fig.5).Among the selected OPA1 homologies,Pe-OPA1 had the closest relationship with mollusk OPA1.
3.2 High expression of OPA1 in coelomic fluid
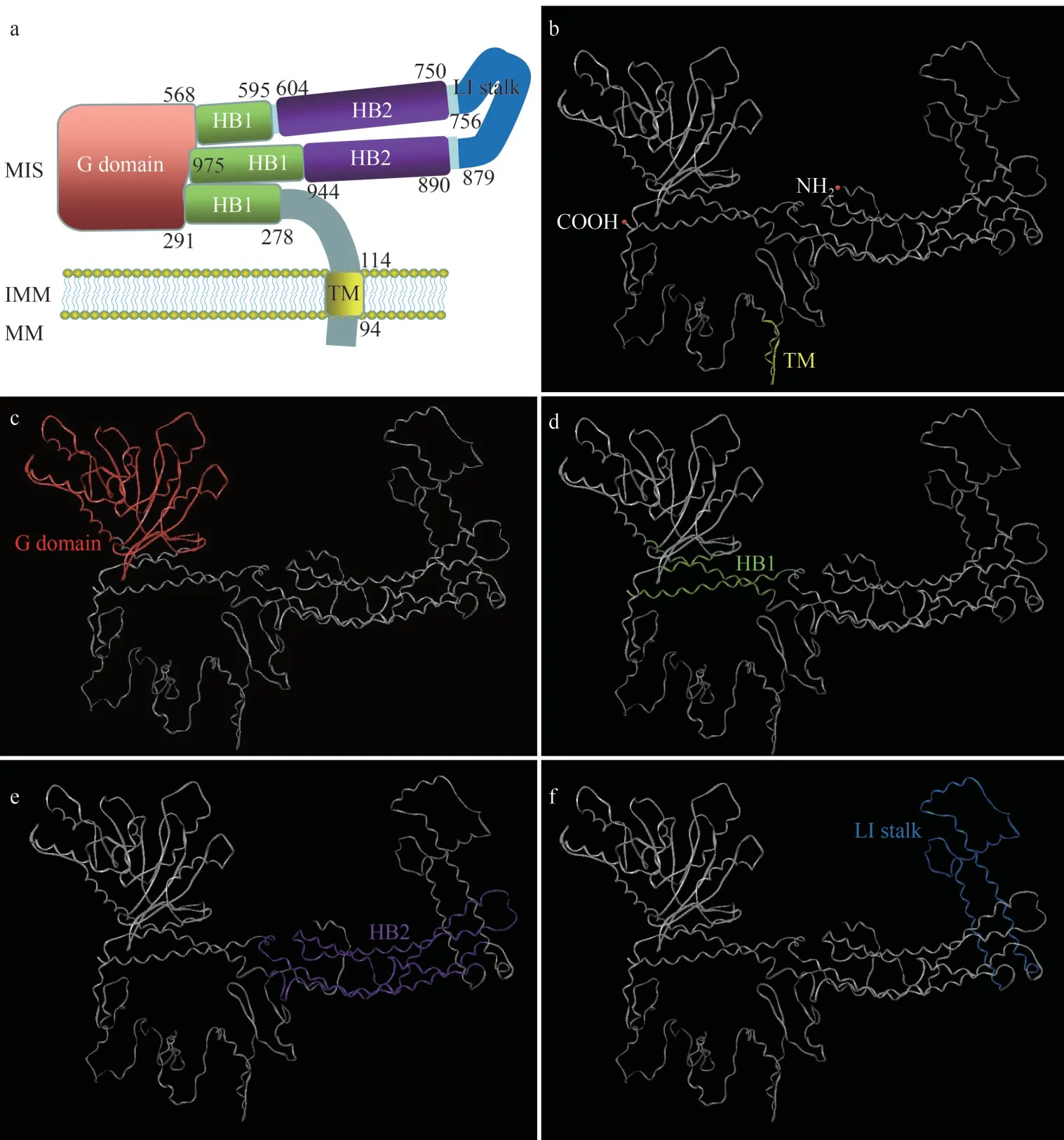
Fig.2 The protein structure of Pe-OPA1
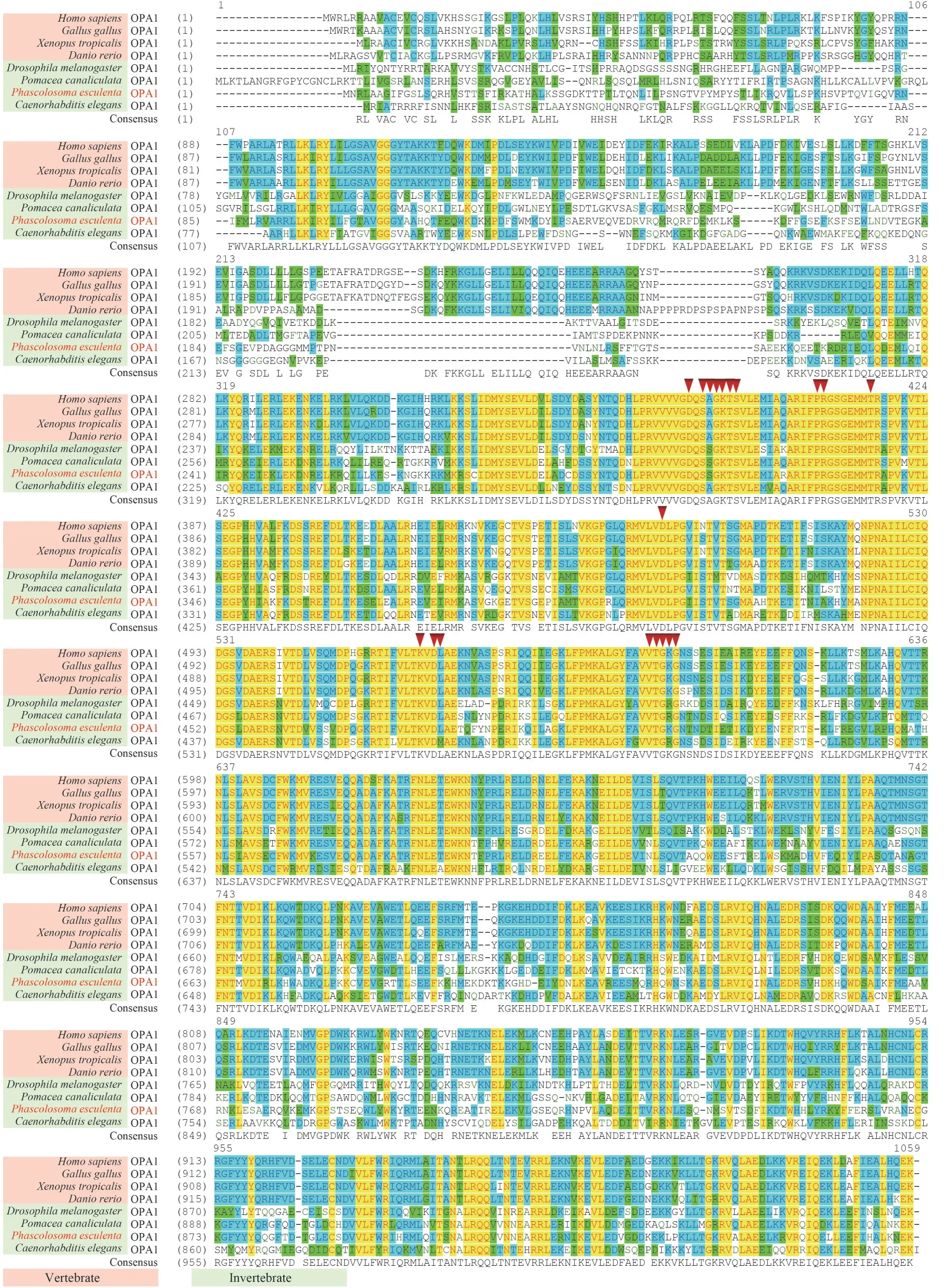
Fig.3 Comparison of OPA1 homologous proteins

Fig.4 Consensus positions between the OPA1 homologous proteins

Fig.5 Phylogenetic tree based on the amino acid sequences of OPA1 proteins
Regarding spatial expression,Pe-opa1mRNA was expressed in coelomic fluids, intestines,retractor muscles, body walls, and nephridia, with the highest expression observed in the coelomic fluid (Fig.6a).Studies have shown that mammalian OPA1 has multiple isoforms.Following CCCP treatment, only the lower molecular weight isoform was detected, corresponding to the S-OPA1 isoform(Fig.6b).ThePe-OPA1 protein was highly expressed in coelomic fluids and nephridia, significantly higher than in retractor muscles (Fig.6c).
3.3 High expression of OPA1 during breeding season
Regarding temporal expression,Pe-opa1mRNA andPe-OPA1 protein were expressed in different months, with a first increasing and then decreasing trend with the month (Fig.7).When the samples from the breeding (July-September) and nonbreeding (March-June and October-December)seasons were analyzed as a whole, we found thatPeopa1andPe-OPA1 had significantly higher expression during the breeding season (Fig.7).
3.4 High expression of OPA1 in coelomic fluid spermatid
ThePe-OPA1 protein was highly expressed in the components containing spermatids (ST1 and ST2) and lowly expressed in the components lacking spermatids (RST), indicating thatPe-OPA1 is mainly distributed in spermatids in the coelomic fluid (Fig.8a).
3.5 OPA1 localization in mitochondria and consistent expression during spermiogenesis
To clarify its subcellular localization in spermatid components, the mitochondria and cytoplasms were separated, and ACTB (actin B) and SDHA(succinate dehydrogenase subunit A) was used to analyze the purity of the separated proteins.We found thatPe-OPA1 was mainly localized in mitochondria (Fig.8b).
Moreover,Pe-OPA1 consistently co-localized with mitochondria during spermiogenesis (Fig.9).In early and middle spermatids,Pe-OPA1 and mitochondria are randomly distributed in the cytoplasm or unilaterally in cells (Fig.9a1-a4 & b1-b4).In late spermatids, OPA1 and mitochondria were distributed mainly on one side of the cell(Fig.9c1-c4).In mature sperm,Pe-OPA1 and mitochondria are located in the midpiece (Fig.9d1-d4).
4 DISCUSSION
Mitochondrial morphology changes during spermatogenesis (De Martino et al., 1979; Hales and Fuller, 1997; Haseeb et al., 2018; Gao et al., 2020).In somatic cells, OPA1 regulates mitochondrial fusion and cristae morphology (Herlan et al., 2003;Frezza et al., 2006; Agier et al., 2012; Wai and Langer, 2016; Gao and Hu, 2021).We,therefore, hypothesized thatPe-OPA1 is involved in regulating mitochondrial morphology by regulating mitochondrial fusion and cristae morphology during spermatogenesis.In this study, we cloned the full-length cDNA ofPe-opa1fromP.esculentaand analyzed its (gene and protein product)expression and localization in different tissues, the breeding cycle, coelomic fluid components, and spermiogenesis, to determine whether thePe-OPA1 potentially regulates mitochondrial morphology during spermiogenesis.
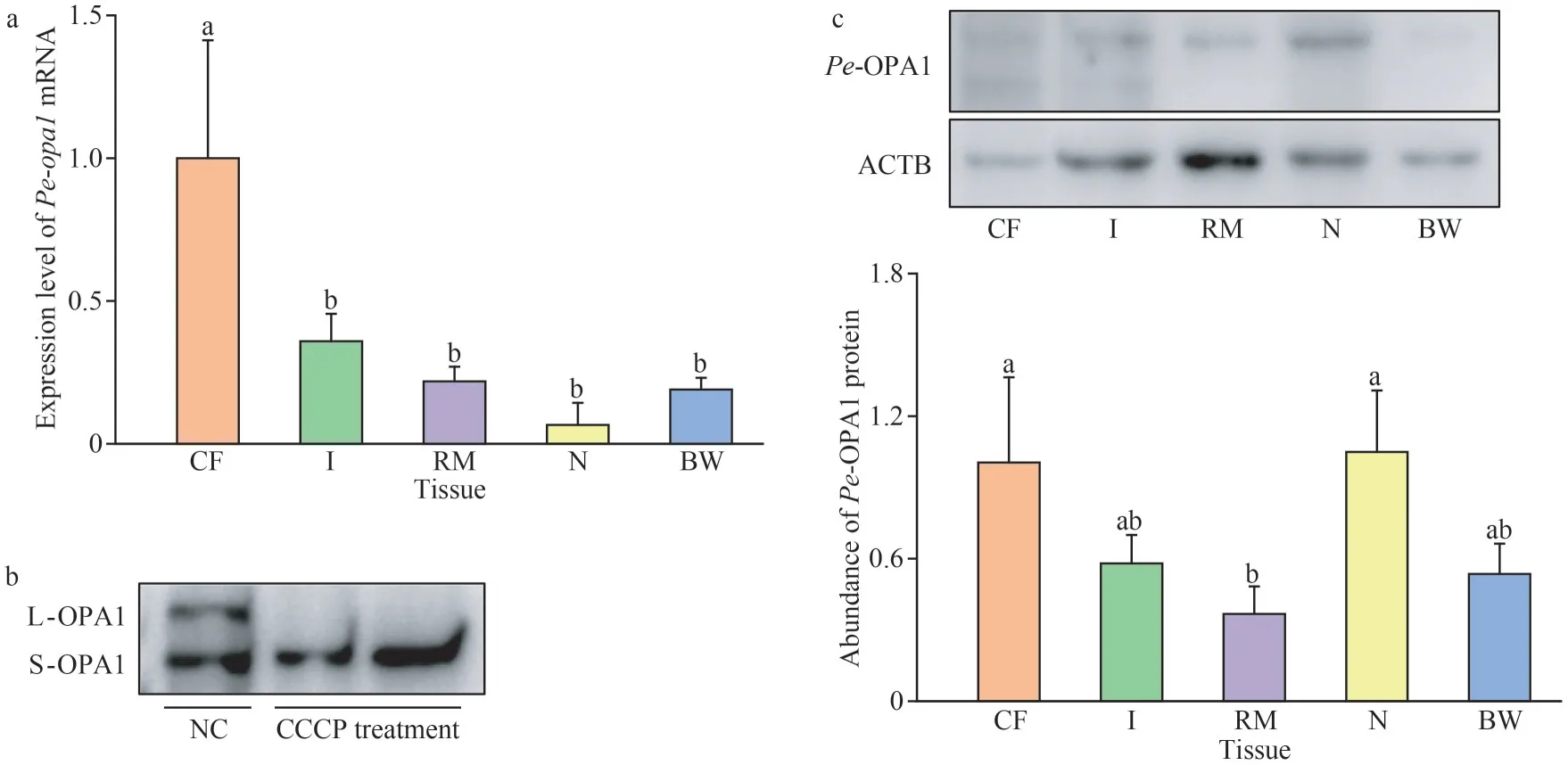
Fig.6 Spatial expression of Pe-opa1 mRNA and Pe-OPA1 protein in P.esculenta tissues
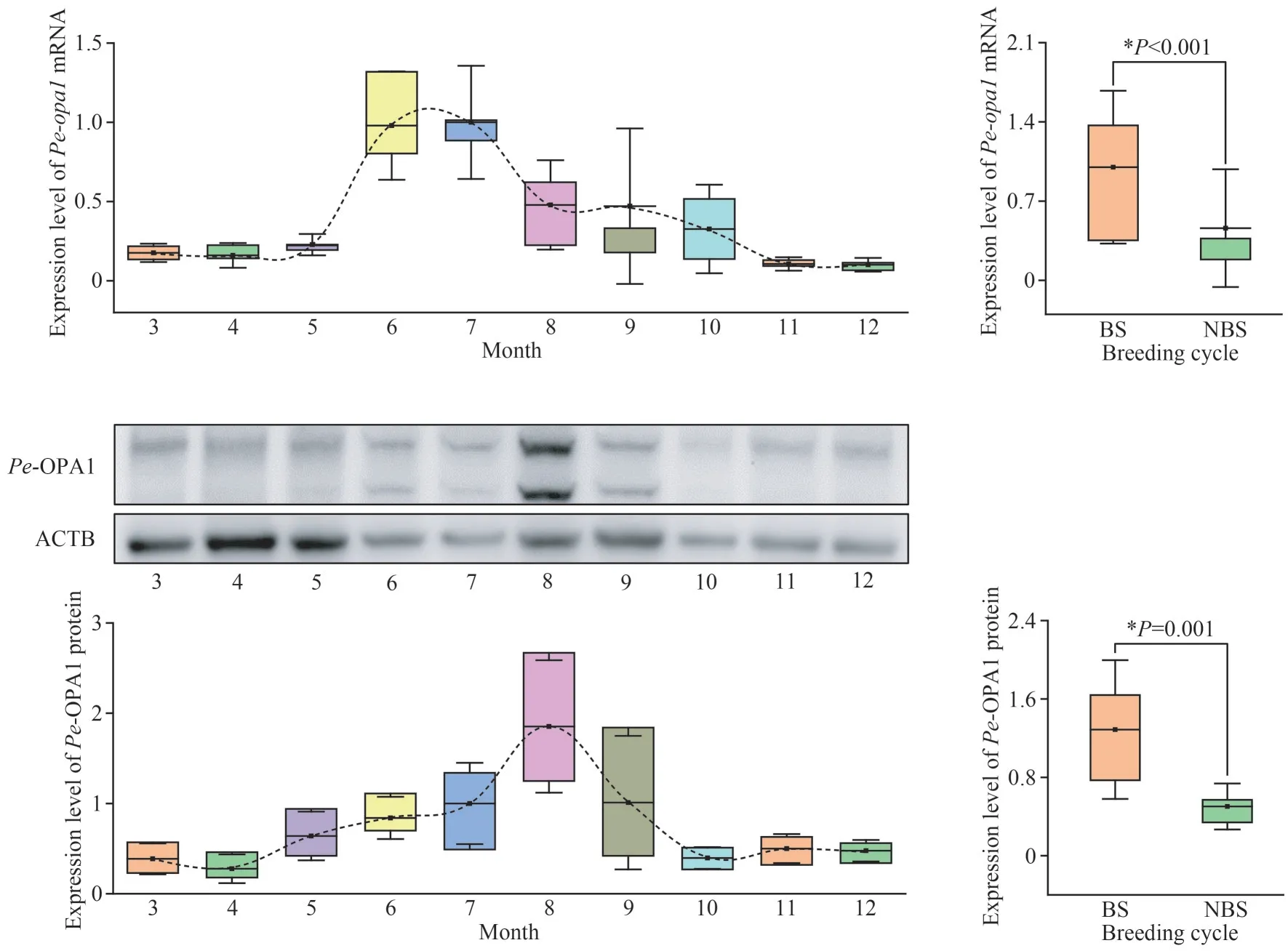
Fig.7 Temporal expression of Pe-opa1 mRNA and Pe-OPA1 protein in the coelomic fluid of P.esculenta in different months and during the breeding cycle
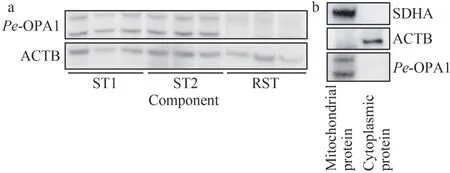
Fig.8 Expression of Pe-OPA1 in different components of the coelomic fluid and subcellular localization in early spermatids
4.1 Protein structure of OPA1
It has been reported that the mammalian OPA1 protein has a mitochondrial targeting signal, a transmembrane domain, a GTPase domain, two helical bundle domains, and a lipid-interacting stalk(Yapa et al., 2021).The transmembrane domain targets OPA1 to the inner mitochondrial membrane;the GTPase domain binds and hydrolyzes GTP; the helical bundle domains are responsible for oligomerization between OPA1 proteins; the stalk is responsible for interaction with lipids (Yapa et al.,2021).Here, through sequence alignment and protein structure comparisons with human andS.cerevisiaeOPA1 (also known as MGM1; Li et al.,2019; Yan et al., 2020) and bioinformatics analyses,we predicted and constructed the protein domain pattern ofPe-OPA1 and found it to be consistent with other OPA1 proteins.Over the course of evolution, the OPA1 protein are conservative, with the consensus positions of OPA1 homologs more than 45% (in the sequences analyzed).The GTP binding site is consistent in invertebrates to humans,indicating that the ability and molecular mechanism ofPe-OPA1 to bind and hydrolyze GTP are likely conservative.Considering our data, we infer thatPe-OPA1 has similar functions to the functionally characterized OPA1 proteins in other species.

Fig.9 Expression and localization of Pe-OPA1 during spermiogenesis in P.esculenta
4.2 Function of OPA1 in mitochondria
OPA1 regulates inner mitochondrial membrane fusion and affects the physiological function of mitochondria by regulating the morphology of the mitochondrial cristae (Del Dotto et al., 2018; Gao and Hu, 2021; Gilkerson et al., 2021).For example,the absence of OPA1 leads to mitochondrial fragmentation and abnormal cristae morphology in HeLa cells (Olichon et al., 2003).OPA1 has several isoforms; for example,S.cerevisiaeOPA1 (MGM1)has two isoforms, a long and short isoform (Herlan et al., 2003).Humans have eight splice variants ofopa1(Del Dotto et al., 2018).In mice, five OPA1 isoforms were detected by western blotting, two larger molecular weight (L-OPA1) and three lower molecular weight isoforms (S-OPA1; Anand et al.,2014).It has been reported that L-OPA1 and SOPA1 have different functions.OPA1 can regulate mitochondrial morphology through changes in the abundance of L-OPA1 and S-OPA1 (Wai and Langer, 2016).It has been shown that L-OPA1 promotes mitochondrial inner membrane fusion and maintains the morphology of mitochondrial cristae,while S-OPA1 promotes mitochondrial fission (Wai and Langer, 2016).Under conditions of reduced mitochondrial membrane potential, L-OPA1 is hydrolyzed to S-OPA1.Treatment of HeLa cells with CCCP, an oxidative phosphorylation uncoupler,reduces the level of L-OPA1 and increases the level of S-OPA1, leading to mitochondrial fragmentation(Ishihara et al., 2006; Griparic et al., 2007).In this study, we found twoPe-OPA1 isoforms inP.esculenta, indicating that the presence of multiple isoforms of OPA1 is conserved in different animals.Moreover, CCCP treatment led to the hydrolysis of L-OPA1 to S-OPA1 inP.esculenta, which is consistent with that reported in mammals (Ishihara et al., 2006; Griparic et al., 2007).Therefore, we speculate thatPe-OPA1 coordinates the levels of L-OPA1 and S-OPA1 to regulate mitochondrial fusion and physiological functions through similar mechanisms as in mammals.
4.3 OPA1 is a potential regulator for mitochondrial morphology during spermiogenesis in P.esculenta
During spermatogenesis, mitochondria undergo morphological changes.This has been reported inDrosophilamelanogaster,Pampusargenteus,Pelodiscussinensis, andRattusnorvegicusamong others (De Martino et al., 1979; Hales and Fuller,1997; Haseeb et al., 2018; Gao et al., 2020).InD.melanogaster, abnormal morphology of mitochondria causes the failure of spermiogenesis, leading to male sterility (McQuibban et al., 2006).InMusmusculus,abnormal mitochondria length and size leads to stagnation of spermatogenesis or poor sperm quality,which reduces male fertility or leads to sterility(Varuzhanyan et al., 2019, 2021a, b).We have observed mitochondria undergoing morphological changes during spermatogenesis inP.esculenta(Gao et al., 2022), implying the involvement thereof inP.esculentaspermiogenesis.The mechanism and role of mitochondrial morphology regulation, however,remains to be fully clarified.Recently, proteins related to mitochondrial dynamics regulation, such as mitofusins (MFN1 and MFN2) and mitochondrial fission factor (MFF), have been reported to be involved in mitochondrial morphology during spermatogenesis.For instance, knockdown ofmfn1or/andmfn2 inM.musculusreduced mitochondrial fusion and triggered abnormal mitochondrial morphology,which affected the mitochondrial respiratory chain complex and mitochondrial ribosomal subunits,resulting in abnormal differentiation of spermatogonia,spermatocyte meiotic arrest, and male sterility(Varuzhanyan et al., 2019).Moreover, a lack of MFF reduced mitochondrial fission and caused abnormal mitochondrial morphology (length and size increased) in spermatids (Varuzhanyan et al.,2021a).This led to abnormal morphology of sperm midpieces, flagellum kinks, and discontinuous distribution of mitochondria in the midpiece and triggered the poor activity of the respiratory chain complex IV in sperm, thereby reducing the motility of sperm and causing low fertility in males(Varuzhanyan et al., 2021a).OPA1 is an important regulator for mitochondrial dynamics as discussed above.We, therefore, inferred that OPA1 is a regulator for mitochondrial morphology during spermiogenesis.In this study, we found that the OPA1 is highly expressed in coelomic fluid, the site of spermiogenesis, and higher expression in the breeding stage than non-breeding stage.In addition, thePe-OPA1 is mainly in spermatid in coelomic fluid and incessantly expressed and localized in mitochondria during spermiogenesis, suggesting its involvement in spermiogenesis by functioning within the mitochondria.These results support our hypothesis that OPA1 is a regulator for mitochondrial morphology during spermiogenesis inP.esculentapossibly by regulating mitochondrial fusion and cristae morphology during spermiogenesis.This study, therefore, provided some new insights into the mechanism of mitochondrial plasticity during animal spermatogenesis.
5 CONCLUSION
The OPA1 protein is conservative in animal evolution with the domains of a transmembrane domain, a GTPase domain, two helical bundle domains, and a lipid-interacting stalk.ThePeopa1mRNA and protein are higher expressed in the coelomic fluid in different tissues and in the breeding stage.The OPA1 protein is mainly in spermatid in coelomic fluid, and incessantly expressed and localized in mitochondria during spermiogenesis, suggesting its involvement inP.esculentaspermiogenesis.These results indicate that the OPA1 may be a potential protein in modeling mitochondrial morphology by regulating the mitochondrial fusion and cristae during spermiogenesis, which may affect the mitochondrial physiology and are propitious to spermiogenesis inP.esculenta.
6 DATA AVAILABILITY STATEMENT
The data of this study are available from the corresponding author upon reasonable request.
7 ACKNOWLEDGMENT
The authors wish to thank all the members from the Fish Reproduction Physiology Laboratory at Ningbo University for fruitful discussions.
 Journal of Oceanology and Limnology2024年1期
Journal of Oceanology and Limnology2024年1期
- Journal of Oceanology and Limnology的其它文章
- Contrasts of bimodal tropical instability waves (TIWs)-induced wind stress perturbations in the Pacific Ocean among observations, ocean models, and coupled climate models*
- Variability of the Pacific subtropical cells under global warming in CMIP6 models*
- Identification of thermal front dynamics in the northern Malacca Strait using ROMS 3D-model*
- Magmatic-tectonic response of the South China Craton to the Paleo-Pacific subduction during the Triassic: a new viewpoint based on Well NK-1*
- An improved positioning model of deep-seafloor datum point at large incidence angle*
- Microplastics in sediment of the Three Gorges Reservoir:abundance and characteristics under different environmental conditions*