
施加生物炭对谷子干物质积累、转运、分配和土壤理化性质的影响
2024-02-27 07:54李博洋楚睿雯张岁岐严加坤
作物学报 2024年3期
李博洋 叶 茵 楚睿雯 井 苗 张岁岐 严加坤,*
1 榆林学院生命科学学院, 陕西榆林 719000; 2 九江市农业科学院, 江西九江 332000; 3 榆林市农业科学研究院, 陕西榆林 719000;4 中国科学院水利部水土保持研究所 / 黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西杨凌 712100
谷子(Setariaitalica(L.) Beauv.)起源于中国, 在新石器时代中早期完成驯化, 成为农耕文化的主要栽培作物[1]; 谷子具有较好的耐旱、耐贫瘠性, 适宜在我国北方干旱和半干旱地区种植[2]。我国谷子种植面积约占世界谷子种植面积的80%, 产量占世界谷子总产量的90%[3]。目前, 随着人们对健康生活的追求日益增加, 具有较高营养价值的谷子(小米)已成为我国北方种植业结构调整和民众膳食结构改善的主体作物而得到更加广泛的关注[4]。
生物炭是秸秆等农林废弃物生物质在缺氧条件下热裂解形成的稳定的富碳产物[5], 具有原料来源广泛、空隙结构好、吸附力强等特点[6], 是理想的农用基质材料。国内外研究表明, 适量生物炭可以提高玉米[7-8]、水稻[9]、小麦[10]等作物的产量; 并适当提高了他们的光合速率、干物质积累量、籽粒的分配比例和转运效率等; 但是有研究发现生物炭对植物生长和作物产量产生的影响会表现出两面性, 高生物炭施用量使得番茄生物量显著降低[11]。因此,在不同地区不同作物上使用生物炭需要经过严格试验验证。目前生物炭农业应用研究主要集中在小麦、水稻等大宗主粮作物上, 少有生物炭对谷子生长影响研究报道。并且相关研究多局限于某一时期或某部位的干物质积累方面, 缺乏对谷子干物质积累转运及分配方面的系统性研究。生物炭在土壤中可以充当庇护所、提供养分和改善土壤条件; 然而, 生物炭携带的可能威胁微生物群落的有毒物质不应被忽视[12]。此前, 主要应用于土壤固碳减排机制[13]、根际微生物群落和根系分泌物[14]以及生态修复[15]等方面, 并且研究多集中于潮土、黑土、壤土等[16]类型土壤上, 有关其对沙地尤其是新开垦沙地土壤理化性质影响的研究还相对较少。因此, 研究生物炭对沙地土壤理化性质的影响并评估其在沙地农业中的应用潜力对于沙地农业可持续发展具有重要意义。
基于此, 本研究利用陕北毛乌素新开垦沙地,以谷子(榆谷抗1)为研究对象, 通过2 年的大田试验,研究施用不同用量的玉米秸秆生物炭对土壤理化性质及谷子光合参数、干物质及产量等相关指标的影响, 以期为毛乌素沙地及其类似环境新垦地土壤改良及生物炭的合理利用提供参考依据。
1 材料与方法
1.1 试验设计
本试验于2021 年5 月至2022 年10 月在陕西省榆林市榆林学院农牧科教试验基地(38°23′43′′N、109°43′15′′E, 海拔1080 m)进行。该基地耕地表层覆盖30 cm 未耕作过的黄土。试验生物炭的原料为玉米秸秆, 购自辽宁金和福农业科技股份有限公司(生物炭及黄土、沙土相关性质见表1); 供试谷子品种为榆谷抗1, 榆林市农业科学研究院新育成品种, 第十三届优质食用粟品质鉴评会被评为春谷第1 名;分别于2021 年和2022 年5 月播种, 于2021 年和2022 年10 月收获。

表1 生物炭及黄土、沙土的性质Table 1 Properties of biochar, loessal soil, and sandy soil
本试验采用随机区组设计, 设置对照组CK (不添加生物炭)和3 个生物炭施用量处理: C1 (3.0 t hm-2)、C2 (4.5 t hm-2)、C3 (6.0 t m-2), 3 个重复, 共计12 个小区, 每个小区的种植面积为20 m2(4 m×5 m)。播种前进行翻耕处理, 所有小区均施相同底肥:氮180 kg hm-2, 磷(五氧化二磷计) 90 kg hm-2, 钾(氧化钾计) 70 kg hm-2, 不同生物炭所含N、P 以尿素和过磷酸钙平衡。穴播, 每穴2 株谷子, 穴距8 cm, 行距0.5 m。
1.2 项目测定与方法
1.2.1 光合特征参数 使用LI-6400R 便携式光合作用仪(LI-COR, Inc, 美国)于晴天上午 09:00—11:30 测定谷子开花期旗叶的净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr), 光合仪参数设定为CO2浓度设置为 500 μmol mol-1, 光合有效辐射(PAR)为1000 μmol m-2s-1。
1.2.2 茎、叶、穗等干物质重 在开花期(2021年8 月和2022 年8 月)至收获期(2021 年10 月和2022年10 月), 分别在花后0 d (开花期)、7、14、21、28和45 d, 在小区选择生长良好, 长势基本一致的谷子, 沿种植列采集种植长度为50 cm (12 株)谷子地上部, 沿着根与茎的分界处收割, 分离茎叶穗, 烘箱105 ℃ 30 min 杀青, 80℃烘干, 烘干后测量全叶、茎、根和穗的干重。2021 年仅测定开花期和收获期干物质, 各项指标参照宋世龙等[18]和隋鹏祥等[19]的方法, 计算下列指标。计算公式如下:
收获指数=籽粒产量/地上部干物质总量;
花前干物质转运量=开花期地上部干物质积累量-成熟期地上部营养器官干物质积累量;
花前干物质转运效率=花前干物质转运量/开花期地上部干物质积累量×100%;
花前贮藏同化物对籽粒产量的贡献率=花前干物质转运量/成熟期籽粒干重×100%;
花后同化物积累量=成熟期地上部干物质积累量-开花期地上部干物质积累量;
花后同化物对籽粒产量的贡献率=花后同化物积累量/成熟期籽粒干重×100%;
地上部干物质在各器官中的分配比例=各器官干物质量/地上部干物质总量×100%。
1.2.3 土壤理化性质和相关酶活性 收获期采集根际土壤, 按照鲍士旦主编《土壤农化分析》相关方法进行电导率、碱解氮、有效磷和有关酶活性的测定。电导率: 电极法(HJ 802-2016); 有效磷: Na-HCO3浸提-钼锑抗比色法; 碱解氮: 碱解扩散法;脲酶: 苯酚钠-次氯酸钠比色法; 蔗糖酶: 3,5-二硝基水杨酸比色法; 过氧化氢酶: 高锰酸钾滴定法。
1.3 统计分析
采用Microsoft Excel 2016 和SPSS 18.0 进行试验数据计算和单因素方差分析、双因素方差分析以及相关分析; 采用Graphpad Pism 9.5 作图。
2 结果与分析
2.1 生物炭添加对谷子开花期旗叶光合特性的影响
2.1.1 净光合速率(Pn) 谷子开花期旗叶净光合速率在不同处理间存在差异(图1-A)。在2021 年, 施加生物炭显著增加谷子旗叶的净光合速率, 且与CK 相比分别提高44.25%、65.62%和107.45% (P<0.05)。而在2022 年施加生物炭提高谷子旗叶净光合速率, 分别提高42.25%、9.50%和4.65%, 且C1 与CK 差异显著(P< 0.05)。多因素方差分析表明, 年份和处理以及其相互作用极显著影响谷子的开花期旗叶净光合速率(PY*T< 0.01)。
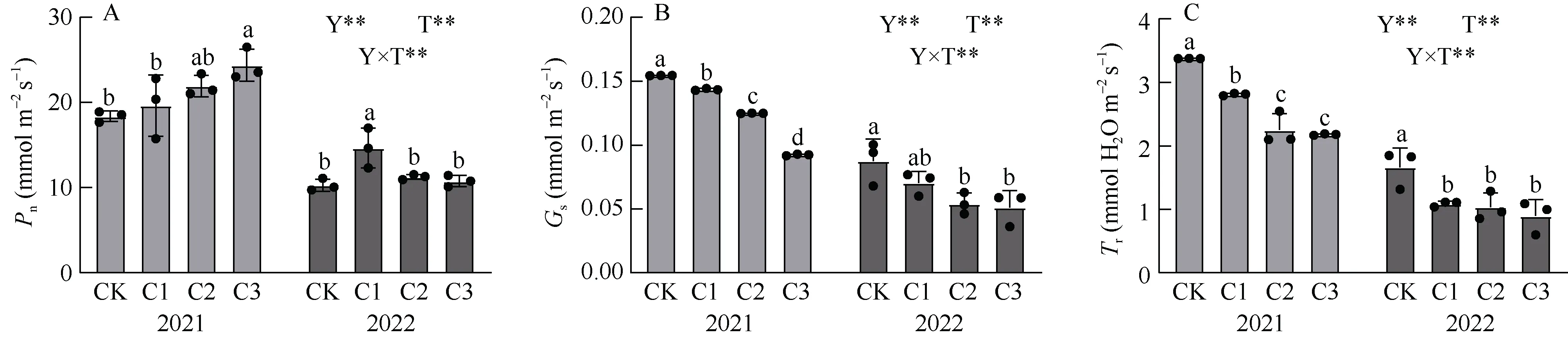
图1 生物炭对谷子旗叶光合参数的影响Fig. 1 Effects of biochar application on flag leaf photosynthetic parameters of foxtail millet
2.1.2 气孔导度(Gs) 谷子开花期旗叶气孔导度在不同处理间存在差异(图1-B)。在2021 年, 施加生物炭显著与对照组之间均存在显著差异, 施加生物炭显著降低谷子旗叶的气孔导度, 与CK 相比分别降低7.05%、19.13%和40.23% (P< 0.05)。在2022年, 施加生物炭均可以不同程度地降低谷子旗叶气孔导度, 与CK 相比分别降低19.57%、38.13%和41.46%, 且C2、C3 与CK 之间差异显著(P< 0.05)。多因素方差分析表明, 年份和处理以及其相互作用极显著影响谷子的开花期旗叶气孔导度(PY*T< 0.01)。
2.1.3 蒸腾速率(Tr) 谷子开花期旗叶蒸腾速率在各处理间存在差异(图1-C)。在2021 年, 施加生物炭与对照之间存在显著差异(P< 0.05), 施加生物炭显著降低谷子旗叶的气孔导度, 随着生物炭施加量的增加, 谷子旗叶蒸腾速率的逐渐下降, 与CK 相比分别降低16.73%、33.33%和35.48% (P< 0.05)。在2022 年, 施加生物炭均可以不同程度地降低谷子旗叶气孔导度, 与CK 相比分别降低34.66%、37.91%和46.21% (P< 0.05)。多因素方差分析表明, 年份和处理以及其相互作极显著影响谷子的开花期旗叶蒸腾速率(PY*T< 0.01)。
2.2 生物炭施加对谷子干物质的影响
2.2.1 生物炭施加对花后谷子叶干物质重的影响
除花后45 d 的旗叶重外, C3 处理其余时期的叶片干物质质量均高于CK 处理; 除花后28 d 的倒七叶和花后21 d 的倒十三叶外, 花后所有时期所有叶片C2 处理下的叶片干物质质量均高于CK处理; 但是C1 处理CK 处理之间不同时期不同部位叶片干物质质量差别不大(图2-A~M)。对于总叶重来说(图2-N), C2、C3 处理下花后不同时期叶片总干物质重均高于CK 组, C1 处理与CK 组差别不大。在2021 年开花期, 谷子叶干物质重呈现先增加后减小的趋势, 以C1 处理下叶干物质增加最多; 在2022 年开花期, 生物炭施加后谷子叶重呈现逐渐上升的趋势, 以C3 处理下叶干物质增加最多(表2)。在2021 年和2022 年收获期, 施加生物炭均增加了谷子叶干物质重, 在2021 年叶干物质重呈现先增加后减小的趋势, 以C2 处理下叶干物质增加最多; 在2022 年以C3 处理下叶干物质增加最多(表3)。年份的变化对开花期叶的干物质重有显著影响(P< 0.01)。
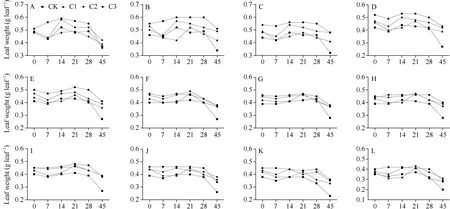
(图2)

表2 生物炭对谷子开花期叶重、茎重和地上部总重的影响Table 2 Effects of biochar application on leaf weight, stem weight, and gross aboveground weight of millet at flowering stage

表3 生物炭对谷子收获期叶重、茎重和地上部总重的影响Table 3 Effects of biochar application on leaf weight, stem weight, and gross above-ground weight of millet harvest
2.2.2 生物炭施加对花后谷子茎干物质重的影响
除花后14 d 和花后28 d C1 和C2 茎干物质重略低于CK 外, 其余时期C1、C2、C3 处理下茎干物质重均高于CK 组(图2-O); 在2021 年开花期, C1 与CK 茎干物质重接近, C2、C3 处理下茎干物质重略有下降; 但是在2022 年开花期, 施加生物炭均提高了谷子茎干物质重, 较 CK 相比分别提升 2.53%、2.92%、6.35% (表2)。在2021 年收获期(表3), 较CK 相比施加生物炭均提高了谷子茎干物质重, 分别提升14.69%、42.71%、24.95%; 在2022 年收获期, 较CK 相比施加生物炭均提高了谷子茎干物质重, 分别提升0.46%、1.19%、25.36%。年份的变化对开花期茎的干物质重有显著影响(P< 0.01)。
2.2.3 生物炭施加对花后谷子穗干物质重的影响
施加生物炭在花后不同时期均增加了谷子的穗干物质重, 并且拟合CK、C1、C2、C3 的穗干物质重(图2-P)发现, 不同处理间谷子穗重的增长率由小到大分别为CK (y= 0.245x+ 3.686,R2= 0.9182)、C1(y= 0.292x+ 3.5467,R2= 0.9663)、C2 (y= 0.3041x+4.0501,R2= 0.9712)、C3 (y= 0.3596x+ 4.3814,R2=0.9733)。表明, 谷子穗重以C3 处理下增长率最高,生物炭的施加对穗干物质重具有积极影响, 且随着生物炭施加量的增加, 穗干物质增加效果越好。
2.2.4 生物炭施加对花后谷子地上部总重的影响
在2021 年开花期, 施加生物炭降低了谷子地上部总重; 但是在2022 年开花期, 施加生物炭增加了谷子地上部总重, 与CK 相比分别增加6.28%、7.31%、10.26% (表2)。在2021 年和2022 年收获期(表3), 施加生物炭均增加谷子地上部总重, 与CK相比分别增加33.12%、40.62%、16.74%和9.62%、12.77%、32.08%。年份的变化对叶的干物质重有显著影响(P< 0.01)。
2.3 生物炭施加对谷子花后干物质转运和干物质分配的影响
生物炭处理下花前干物质转运量(图3-A)、花前干物质转运效率(图3-B)和花前干物质对籽粒产量的贡献率(图3-C)均低于CK 组, 且C2、C3 处理显著(P< 0.05)。施加生物炭谷子花后同化物积累量较CK 分别增加125.72%、114.04%、105.33% (图3-D),其中以C1 与CK 差异显著(P< 0.05); 花后同化物积累量对籽粒产量的贡献率较CK 分别增加61.18%、58.49%、64.38% (图3-E), 但是各处理间无显著差异。说明, 生物炭施加可以提高花后同化物积累量和花后同化物积累量对籽粒产量的贡献率进而提高收获期穗干物质重。
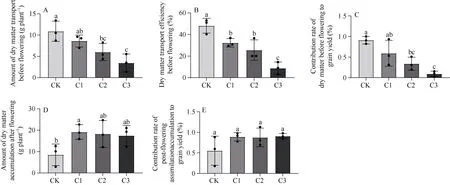
图3 生物炭施加对谷子干物质转运的影响Fig. 3 Effect of biochar application on dry matter transport of millet
干物质是生物学产量形成的基础, 干物质的转运决定营养物质的流向和产量的高低。干物质的分配比例对谷子高产有着重要贡献。在开花期, 谷子叶和茎在不同生物炭处理下干物质占比增加且具有显著性(P< 0.05), 且随着生物炭施加量的增加,叶的干物质占比分别提高5.56%、7.83%、16.72%(图4-A), 茎的干物质占比分别提高2.70%、4.45%、6.06% (图4-B); 但是穗的干物质重在各处理略低于CK 处理(图4-C)。在收获期, 不同生物炭处理下谷子叶和茎干物质占比逐渐降低且差异具有显著性(P< 0.05), 随着生物炭施加量的增加, 叶干物质占比分别较CK 组降低5.76%、10.16%、20.15%, 茎干物质占比分别较CK 组降低8.77%、14.10%、13.07%;但是在不同的生物炭施加量下穗干物质重均高于CK 处理, 且C2、C3 要显著高于CK (P< 0.05), 各处理下穗干物质重较CK 分别提高31.79%、20.64%、15.66%。说明, 生物炭的施加可以提高收获期穗干物质分配的比例, 促进花后谷子干物质在穗部的积累, 从而达到增产的效果。
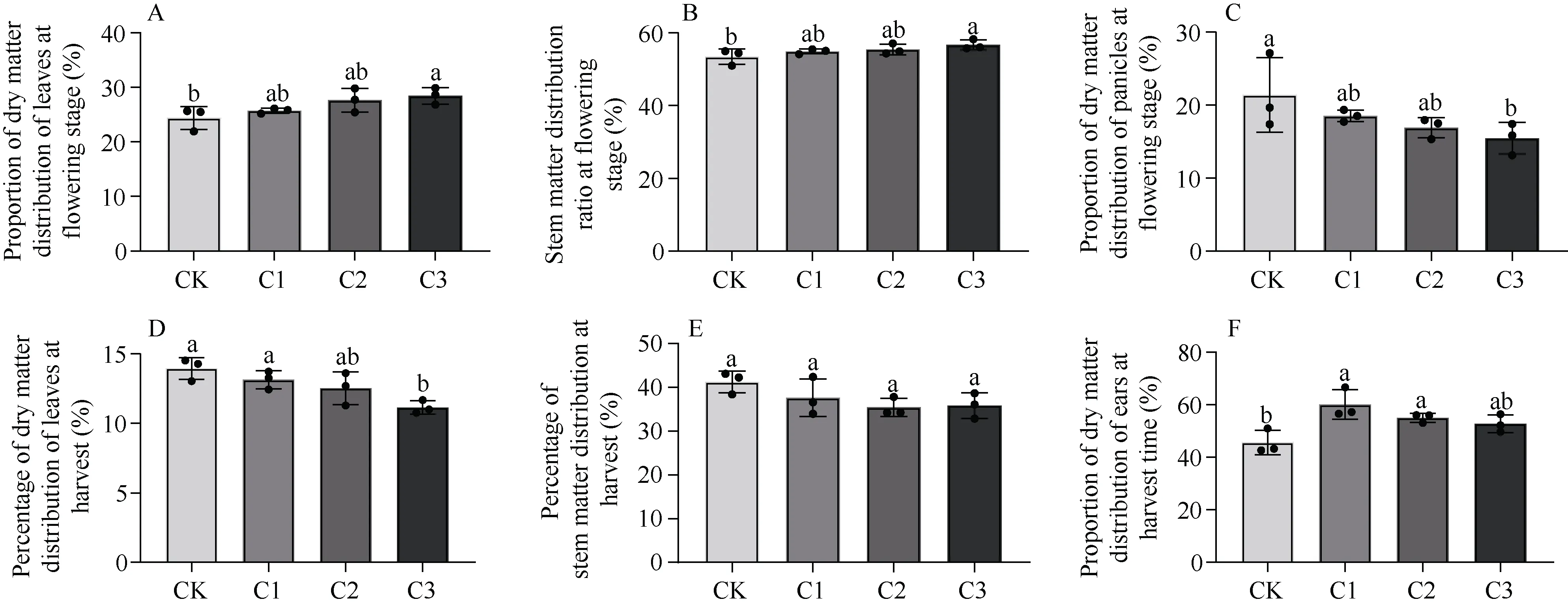
图4 生物炭施加对谷子开花期和收获期各营养器官干物质分配比例的影响Fig. 4 Effects of biochar application on the proportion of dry matter distribution in various vegetative organs at flowering and harvest stages of millet
2.4 生物炭施加对谷子2 年产量、千粒重及收获指数的影响
由表4 可知, 不同生物炭施加量对谷子产量的影响不同。在2 年的大田试验中, 不同生物炭处理下谷子产量均呈上升趋势; 在2021 年中, C2、C3 处理下产量显著(P< 0.05)增加; 在2022 年中, C1、C2、C3 处理下产量显著增加(P< 0.05), 且C3 处理与C2、C1 处理之间产量也增加显著(P< 0.05)。施加生物炭后C1、C2、C3 处理下2 年谷子产量平均增加19.31%、28.26%、38.46%。在2 年的大田试验中, 不同处理对谷子千粒重的和收获指数影响不同; 在2021 年, 不同处理间具有一定差异, 生物炭处理下千粒重和收获指数略低于CK 处理; 在2022 年, 不同的生物炭处理下谷子千粒重和收获指数均呈现先增大后减小的趋势, 且C2 处理下的谷子千粒重显著增高(P< 0.05)。在2022 年。另外在2 年的试验中,生物炭施加量对谷子产量和千粒重具有极显著影响(P< 0.01), 年份和生物炭施加量的交差影响对千粒重也有极显著影响(P< 0.01)。
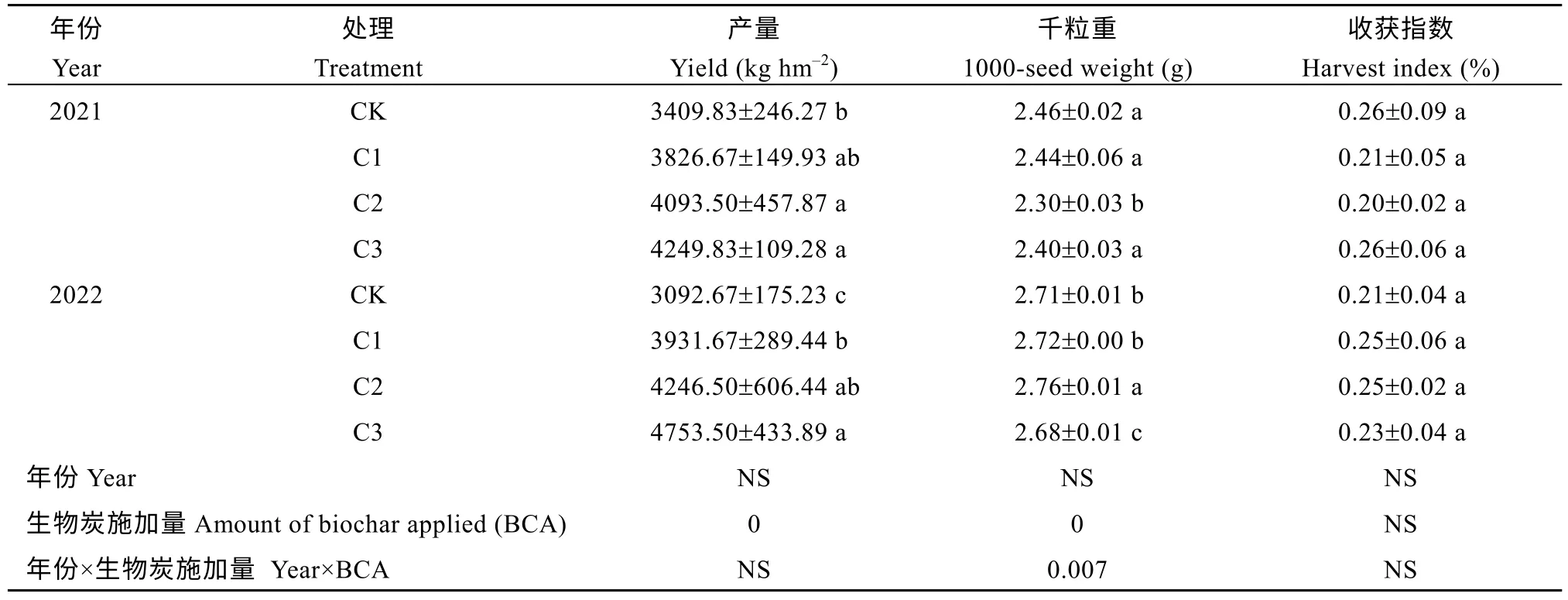
表4 生物炭对对谷子产量、千粒重及收获指数的影响Table 4 Effects of biochar application on millet yield, 1000-seed weight, and harvest index
2.5 相关性分析
相关性分析(表5)表明, 净光合速率与收获期叶重、开花期旗叶重、开花期叶重、开花期茎重、开花期地上部总重呈极显著正相关(P< 0.01); 与收获期茎重、开花期全株总重呈显著正相关(P< 0.05)。收获期产量与收获期穗重呈极显著正相关(P< 0.01);与收获期茎重、收获期地上部总重、收获期全株总重呈显著正相关(P< 0.05)。收获期茎重与开花期旗叶重、开花期叶重、开花期茎重、开花期地上部总重、开花期全株总重呈极显著正相关(P< 0.01)。收获期穗重与收获期茎重、收获期地上部总重、收获期全株总重呈显著正相关(P< 0.05)。收获期地上部总重与收获期全株总重呈极显著正相关(P< 0.01)。
2.6 生物炭施加对土壤理化性质的影响
2.6.1 生物炭施加对土壤养分的影响 在2 年的试验中, 生物炭处理下土壤电导率在2 年的试验中均呈现先上升后下降的趋势(图5-A), 且所有处理均高于CK; C2、C3 在2 年中均与CK 处理存在显著差异(P< 0.05), 且均以C2 处理下最高; 在2 年生物炭处理下, 年份和处理对土壤电导率影响极显著(P<0.01), 其相互作用对土壤电导率影响显著(PY*T<0.05)。碱解氮在2021 年所有处理均与CK 存在显著差异(P< 0.05), 且C2 与C1、C3 叶存在显著差异(图5-B,P< 0.05); 在2022 年呈现上升的趋势, C3 处理与CK 存在显著差异(P< 0.05); 在2 年生物炭处理下,年份和处理及其相互作用对土壤碱解氮影响极显著(PY*T< 0.05)。土壤有效磷含量在2 年的试验中均呈现先上升后下降的趋势, 在2021 年, C1、C2 处理显著高于CK (P< 0.05), C3 处理略低于CK (图5-C);在2022 年, C2 处理显著高于CK 处理(P< 0.05), C1、C3 处理高于CK 处理, 但无显著差异; 在2 年生物炭处理下, 年份和处理及其相互作用对土壤有效磷影响极显著(PY*T< 0.01)。

图5 生物炭对土壤理化性质的影响Fig. 5 Effects of amounts of biochar on soil physicochemical properties
2.6.2 生物炭施加对土壤酶活性的影响 生物炭施加后过氧化氢酶(图6-A)和蔗糖酶活性(图6-C)在2021 年呈现逐渐上升, 在2022 年呈现先上升后下降的趋势。2021 年, 过氧化氢酶活性C2、C3 处理与CK 之间差异显著(P< 0.05); 蔗糖酶活性所有处理均与CK 存在显著差异, 且C3 与C2、C1 之间也存在显著差异(P< 0.05)。2022 年, 不同生物炭处理下过氧化氢酶均高于CK 处理, C1、C2 处理与CK 处理存在显著差异(P< 0.05); 不同生物炭处理下蔗糖酶活性均高于CK 处理, 且C2 与CK、C1、C3 存在显著差异(P< 0.05)。脲酶活性在2 年试验中均呈现先增大后减小的趋势(图6-B), 在2021 年, C1、C2与CK 存在显著差异(P< 0.05); 在2022 年, C2 与CK、C1、C3 存在显著差异(P< 0.05)。在2 年生物炭处理下, 在2 年生物炭处理下, 年份和处理及其相互作用对土壤有效磷影响极显著(PY*T< 0.01)。

图6 生物炭对土壤酶活性的影响Fig. 6 Effects of biochar application amounts on soil enzyme activity
3 讨论
3.1 生物炭施加对谷子旗叶光合特性的调控作用
光合作用是作物合成有机物的基本生理代谢过程[20], 它所产生的有机物为作物提供90%~95%的干物质积累, 籽粒产量的20%~30%也来自于植物的光合作用[21], 可见光合作用与作物产量密切相关。叶片是作物进行光合作用的主要器官, 其光合能力的高低是作物生长发育及产量形成的重要因素[22]。提高作物叶片的净光合速率, 可促进光合产物积累及转运, 可提高粒重, 进而增加产量[23-26]。Liu 等[27]发现不同生物炭施加量均增强了植物叶片对叶片CS的光能吸收、捕获和运输, 从而有助于提高番茄产量。赵辉等[28]在对玉米的研究中发现净光合速率、气孔导度和蒸腾速率均随施炭量的增加而增大。还有研究发现施入生物炭可以提高作物叶片的净光合速率、叶片气孔导度和蒸腾速率[29-31], 有利于干物质的积累, 从而提高产量。本研究结果表明, 不同生物炭施加量均可以提高谷子的净光合速率, 从而增加干物质的积累; 但是随着生物炭施用量的增加气孔导度和蒸腾速率呈下降趋势, 说明净光合速率受气孔和非气孔因素的共同作用, 且非气孔因素起主导作用。结果表明不同的生物炭施用量可以促进谷子旗叶光合色素含量和净光合速率使其在花后维持较高水平, 增加同化生产能力, 促进光合产物的合成与积累[32]。与CK 处理相比, 不同生物炭施加量均可以改善谷子旗叶开花期光和性能, 加快光合产物的合成、转运和积累, 促进谷子发育, 提高谷子产量。

3.2 生物炭施加对谷子干物质积累和转运的影响
作物花后同化物的积累和转运对产量形成有直接的影响[33]。程凯乐等[34]对棉花的研究中发现棉花各时期单株干物质积累量均随着炭基肥配施比例的升高而出现先上升后下降的趋势。柳瑞等[35]研究发现施生物炭在一定程度上能增加水稻干物质积累量。宋明丹等[36]研究发现生物炭施加对马铃薯块茎干物质重也有显著促进效果。但是生物炭添加下谷子干物质积累及转运特性的研究较少。本研究表明,生物炭施加可以提高谷子茎、叶、穗和地上部总的干物质重以及花后同化物积累量、花后同化物积累量对籽粒产量的贡献率, 从而提高作物产量, 并以C3 处理下干物质量提高最多。从干物质分配比例角度来说, 生物炭施加提高开花期茎和叶的分配比例,茎和叶生物量积累增强, 使其积累的光合产物可以在花后大量向籽粒转移, 成为谷子产量形成的关键过程[37-38]; 图5 中C、D、E 收获期茎叶干物质量下降, 穗的干物质量上升则证明了这一过程。在本试验中, C1 处理下谷子花后同化物积累量及穗的干物质分配比例最高, 随着生物炭施加花后同化物积累量及穗的干物质分配比例逐渐降低; 但是由于C3 处理下积累了较多的干物质, 可以抵消由于转运量降低带来的产量减少的可能性。可见生物炭能够影响谷子花后同化物积累和转运特性, 有利于穗部积累较多的生物量, 从而对最终产量形成一定的正面影响。
3.3 生物炭施加对土壤理化性质及酶活性的影响
土壤中包含作物生长所必须的养分, 土壤理化性质直接影响作物根系的生长与养分吸收, 根系区域环境是作物生存之根本[39]。生物炭本身含有氮、磷、钾等植物生长所需的各种养分元素, 施入土壤中可以起到补充土壤养分的作用[40]。生物炭丰富的孔隙结构和巨大的比表面积使它能够吸附大量的养分有效抑制淋溶、下渗等因素导致的养分损失, 可以缓慢持续的释放养分[41]。姚奇等[42]在对冬小麦土壤养分和作物产量的研究中发现, 施用生物炭为5 t hm-2时, 土壤有效磷含量最高。张芙蓉等[43]研究发现施用生物炭可以显著降低土壤电导率。魏永霞等[44]在黑土区的研究中发现施炭量和施用年限对土壤碱解氮含量均有极显著的影响。本研究结果表明,除2022 年脲酶含量随生物炭施用量上升外, 生物炭施用后土壤电导率、碱解氮和有效磷的含量, 均呈现先上升后下降的趋势。这是因为生物炭自身矿物质多的特性, 施加到土壤中可增加土壤养分, 但施炭量过高导致氮和磷的固定, 导致含量降低。由此可见施炭量和施用年限均对土壤养分有影响。为此本研究将施炭量、施用年限以及二者交互作用对土壤养分含量进行方差分析, 结果表明, 除了施用年限和生物炭的交叉作用为显著影响, 其余均有极显著影响。上述结果亦可证明施加生物炭可提高土壤养分, 且以C2 处理下, 土壤养分含量增高最多。
武雪萍等[45]和张美存等[46]研究表明, 土壤脲酶、蔗糖酶和过氧化氢酶可作为关键土壤酶学指标,反映土壤的健康状况。王智慧等[47]研究表明, 施用生物炭能够显著提高草甸土土壤脲酶、蔗糖酶、碱性磷酸酶和过氧化氢酶活性。本试验结果表明, 生物炭施加增加了土壤脲酶、蔗糖酶以及过氧化氢酶的活性, 这与郭书亚等[48]的研究结果一致; 但随着生物炭施用量的增加及年份的变化, 较高的生物炭施用量反而不利于酶活性的提高。生物炭还田后,由于自身具有多孔结构及良好的吸附性能, 可以吸附酶促反应的底物, 为土壤酶提供更多的结合位点,提高酶活性; 但是随着施用量及年份的变化, 有研究指出较高的生物炭施用量会呈现更多的C-O 结合位点, 表现出较强的吸附能力[49-50], 可吸附更多的酶分子, 阻碍酶与底物的结合, 进而影响酶活性。
3.4 生物炭施加对谷子产量调控及相关性的影响
谷子千粒重对产量的形成具有重要意义。许多学者认为施加生物炭可提高作物产量及其构成要素[51]。陈欢等[52]认为施加生物炭可以提高小麦产量稳定性;姜慧敏等[53]经过3 年试验研究得出施加生物炭可以提高东北春玉米产量的可持续性, 王志丹等[54]在对玉米的研究中发现施用生物炭增加了玉米穗长、穗行数、穗质量和穗粒质量, 从而提高产量。除此之外研究表明生物炭能够提高番茄[55]、高粱[56]、大麦[57]、水稻[58]等作物的产量。本研究结果表明, 不同生物炭施用量均可增加谷子产量, 增加谷子干物质重以及收获期穗重比例。相关分析结果表明, 谷子产量与收获期穗重、收获期茎重、收获期地上部总重和收获期总重呈显著或者极显著相关。生物炭可以促进花后谷子总的干物质积累, 促进茎、叶干物质向穗部转移, 促进穗部干物质积累, 最终增加谷子产量。
4 结论
不同生物炭施加量均施加提高了谷子开花期净光合速率、花后穗干物质积累量、收获期总干物质量和产量, 且随着生物炭施用量的增加而增加, 以C3 处理下增加最为显著, 与CK 相比提高22.53%、44.42%、62.25%、38.46%; 在干物质转运及营养器官干物质量分配方面, 施加生物炭还提高了谷子花后同化物积累量、花后同化物积累量对籽粒产量的贡献率以及收获期穗的干物质分配比例, 但是不同生物炭施加量处理之间增长不显著或略有下降。在土壤理化性质方面, 不同生物炭施加量下土壤电导率、碱解氮及有效磷含量均有所提升, 且以C2 处理下提升最高, 与CK 相比提高20.24%、32.88%、108.74%; 在土壤酶活性方面, 不同生物炭施加量下土壤脲酶、蔗糖酶和以及过氧化氢酶较CK 处理均有所提高, 且以C2 处理下提升最高, 与CK 相比提高66.76%、81.82%、9.78%。不同生物炭施加量均可以谷子产量, 干物质积累、转运及土壤理化性质有积极影响, 其中以C3 (6.0 t hm-2)处理对谷子增产效果最好。
猜你喜欢
浙江农业科学(2023年10期)2023-10-28
作物学报(2022年1期)2022-11-05
河北科技师范学院学报(2022年2期)2022-08-26
作物学报(2022年9期)2022-07-18
农业工程学报(2022年7期)2022-07-09
中国饲料(2022年5期)2022-04-26
南方农业(2021年18期)2021-12-25
干旱地区农业研究(2021年6期)2021-12-08
今日农业(2021年17期)2021-11-26
浙江农业科学(2019年11期)2019-11-09
