
不同一次性拉伸方式对腘绳肌形态的急性影响
2024-02-21 07:49戴菊红鲍小亚
体育科研 2024年1期
戴菊红,李 晨,鲍小亚,刘 晔*
组成腘绳肌群的半膜肌(Semimembranosus,SM)、半腱肌(Semitendinosus, ST)与股二头肌长头(the Long Head of the Biceps Femoris, BFLH)之间的形态结构差异[1-2]会造成三者的功能特性差异[3-4],并与腘绳肌不同部位的损伤概率有关。 柔韧性训练常被用于腘绳肌放松及损伤后重建, 对腘绳肌组织形态结构的作用存在争议[5-7],传统的静态拉伸与受部分学者推崇的动态拉伸对腘绳肌形态的作用有待证实。 前期研究已证实不同一次性拉伸对BFLH 形态的急性影响[8],但一次性拉伸是否能改变其余腘绳肌形态尚不明确, 不同腘绳肌由一次性拉伸造成形态变化程度是否存在差异有待证明。
以往运用肌骨超声的体育科学研究多使用只能观测小范围在体肌肉的常规超声扫描技术[9],而当所要观测的在体肌肉组织范围高于常规超声扫描范围(4~6 cm)时,全景超声成像技术即为首选方法,且观测结果具有更高的可靠性与精准度[10-12]。
本研究通过全景超声模式采集SM、ST 与BFLH一次性动态拉伸与一次性静态拉伸前后的超声图像, 测量并分析肌束长度与肌肉厚度等形态结构参数的变化, 为解释不同拉伸方式对不同腘绳肌的功能特性的影响提供理论参考。
1 研究对象与方法
1.1 研究对象
招募无腘绳肌损伤的健康男青年10 人, 年龄(23.70±1.16) 岁, 身高 (177.30±4.55)cm, 体重(69.30±5.81)kg。 参与者无下肢肌肉疲劳,实验期间避免进行下肢力量训练及实验以外的柔韧性训练。本研究经北京体育大学运动科学实验伦理委员会审批通过(No.2021117H)。
1.2 研究内容
全景肌骨超声测量参与者进行两种一次性拉伸前及拉伸后各2 min 内的SM、ST 与BFLH 的肌束长度与肌肉厚度。参与者在两个测试阶段分别进行腘绳肌的动态拉伸与静态拉伸。为平衡两种拉伸的不同进行顺序对实验结果的潜在干扰,一半参与者在动态拉伸后休息至少72 h 进行静态拉伸, 另一半参与者则在静态拉伸后休息至少72 h 再进行动态拉伸。
1.3 研究方法
1.3.1 测试流程
使用飞利浦Affiniti70 超声检查仪 (e18-4 线阵探头) 定位放松俯卧的参与者的优势侧SM、ST 与BFLH 体表标记线。 标记线以龙胆紫外科手术笔沿肌肉的肌束走行方向描绘(图1),可使针对参与者的SM、ST 与BFLH 全景超声图像重复扫描路径对应于各肌肉的同一纵断面。 整个试验期间禁止参与者清洗标记线。在每一测试阶段的拉伸干预前,参与者慢跑5 min 热身(TECHNOGYM 跑步机,9 km/h)。参与者以非优势腿站立, 优势腿直膝并将小腿近踝关节处置于同髋关节中心(股骨大转子)高度的横杠上方,直背屈髋至个人所能忍受不适感最强烈、拉伸动作不变形位置并保持30 s 为完成一组静态拉伸,组间休息6 s,连续完成5 组。 参与者听从间隔1.5 s的电子节拍,以优势腿直膝支撑并连续完成10 次燕式平衡动作[13](屈髋与伸髋均耗时1.5 s)为完成一组动态拉伸,组间休息6 s,连续完成5 组。在拉伸前后各2 min[5]内使倾斜方向相同的探头充分接触皮肤,沿SM、ST 与BFLH 标记线对放松俯卧的参与者进行全景超声扫描。
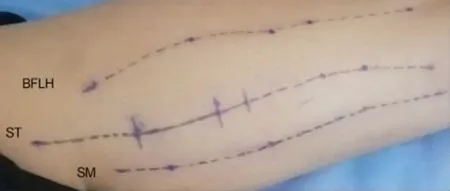
图1 三条腘绳肌肌腹走行路径标记线Figure1 The paths of three hamstrings marked on the skin
1.3.2 超声图像测算与重测信度检验
超声图像测算通过Image pro plus 6.0 进行。 在全景超声图像中将BFLH 和SM 的肌腹沿长轴三等分为近侧端、中部、远侧端区域(图2)。 在每一区域均观测两条位置相近的肌束。 肌束长度为肌束走行方向上的肌束与深、浅腱膜交点之间距,取6 条肌束的长度均值代表该肌肉的肌束长度。 若肌束位于近侧端区域内的长度大于自身长度75%则将其判定为近侧端肌束[14],中部与远侧端肌束均以同一标准判定。 用BFLH 与SM 内3 个区域最厚处的深、浅腱膜间距的均值分别代表BFLH 与SM 的肌肉厚度。
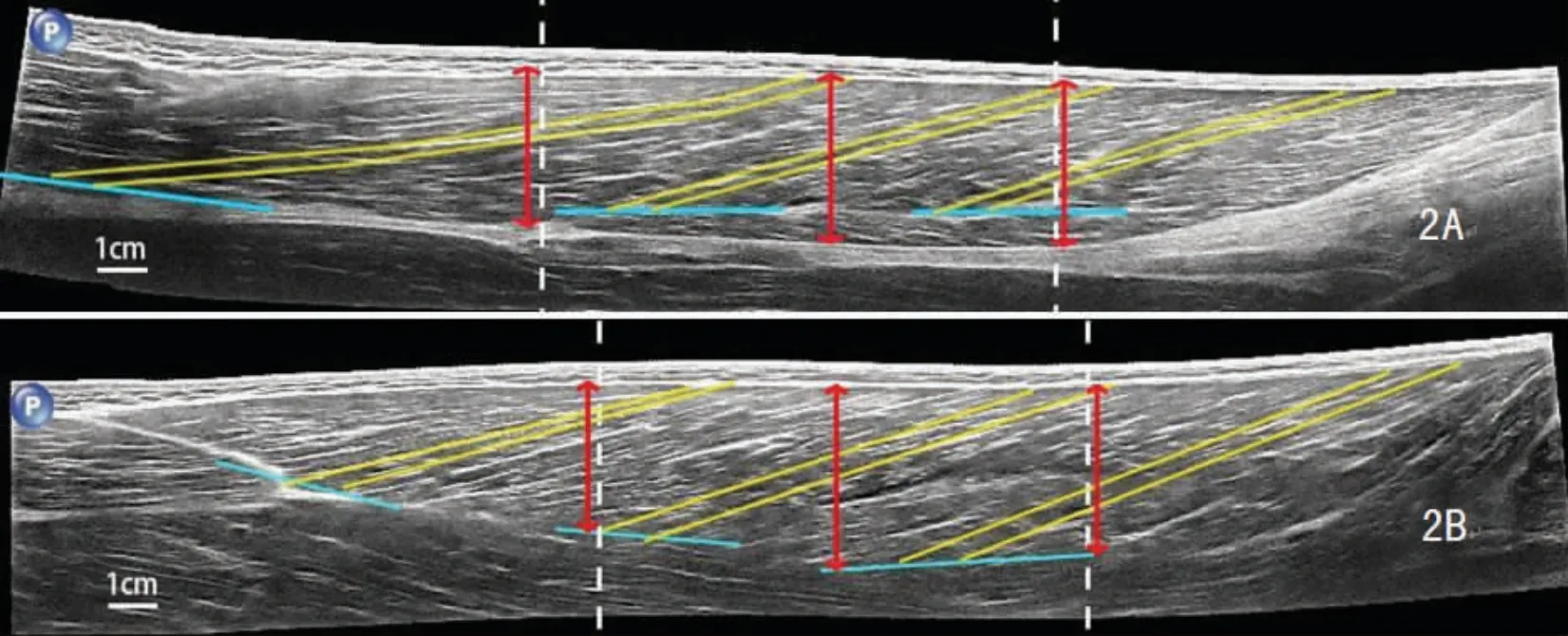
图2 由白色虚线等分为三个区域的BFLH(2A)与SM(2B)肌腹Figure2 The muscle belly of BFLH (2A) and SM (2B) which were both divided equally into three regions by white dashes
由于采集的ST 图像中肌束几乎能够跨越整个狭长的肌腹直接与游离腱相接,测试使用的飞利浦彩色多普勒诊断仪在进行全景超声扫描时连续扫描的长度有限,无法采集到完整涵盖肌束全长的图像。 因此在采集ST 全景超声图像时,以“V”字形腱划为界,沿ST 肌腹走行标记线进行两次扫描 (从近端肌腱结合处至腱划为近侧端,从腱划至远端肌腱结合处为远侧端),分别采集ST 肌腹的近侧端与远侧端全景超声图像(图3)。 在近侧端与远侧端图像中部各取4 条清晰可见的肌束, 以其长度均值分别作为ST 近侧端与远侧端平均肌束长度,以两区域的平均肌束长度之和作为ST 的肌束长度。取腱划近侧端肌肉最厚处、腱划所在区域肌肉最厚处与腱划远侧端肌肉最厚处的深、浅腱膜间距的均值代表ST 的肌肉厚度。
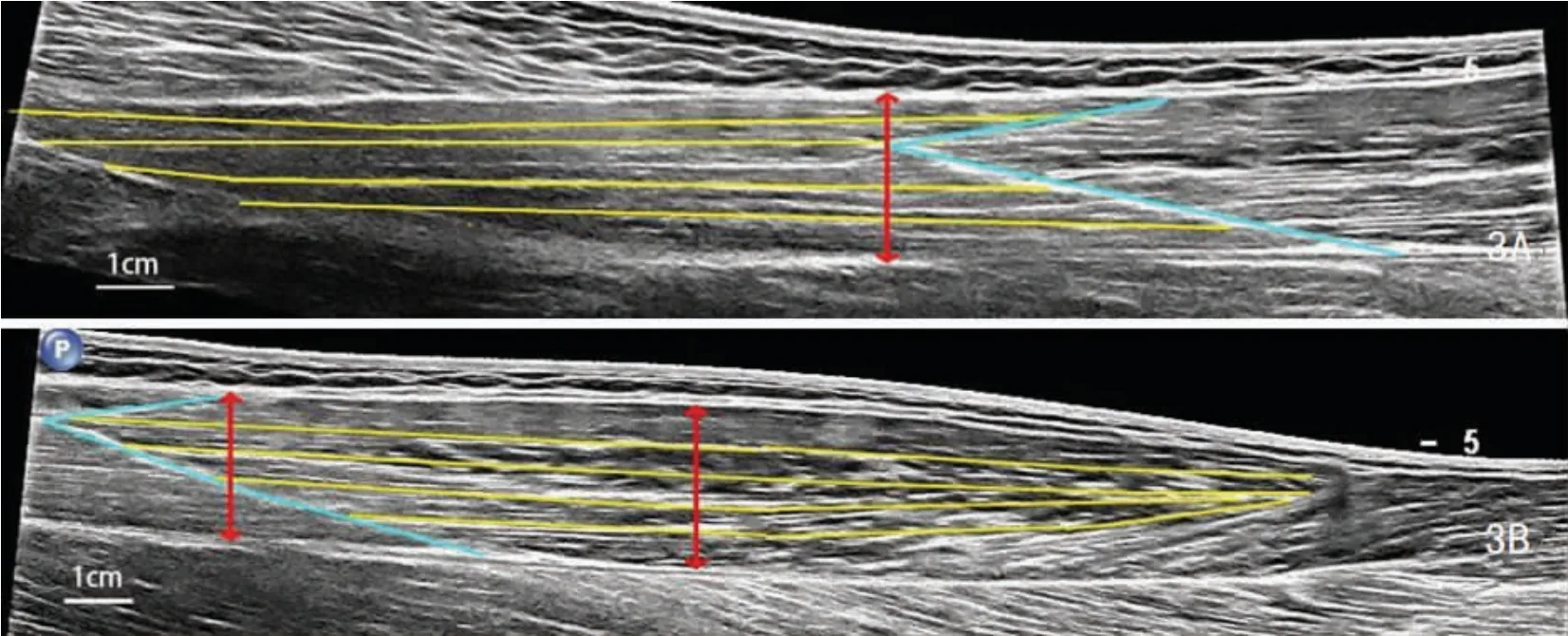
图3 ST 的近侧端(3A)与远侧端(3B)肌腹Figure3 The proximal (3A) and distal (3B) region of the muscle belly of ST
正式测试前两周, 对5 名放松俯卧的参与者进行两次间隔72 h 以上的腘绳肌全景超声测量,检验两次测量结果之间的组内相关系数 (ICC)。 ICC 在0.80 以上代表高重测信度[5]。 检验结果显示了本研究的测试方案对ST、SM 与BFLH 的肌束长度(ICC=0.873、0.845、0.919) 和肌肉厚度(ICC=0.947、0.959、0.985)的重测信度高。
1.4 统计分析
使用SPSS21.0 进行统计分析。 统计学显著性的定义为P<0.05。 各指标经检验后均满足正态分布,表示为均数±标准差。
采用三因素方差分析,检验不同腘绳肌(SM、ST与BFLH)在不同拉伸方式(动态拉伸、静态拉伸)拉伸前后的肌束长度与肌肉厚度的变化。由于前人[15-17]研究证明SM、ST 与BFLH 的结构参数基值存在差异, 为更准确地反映不同腘绳肌间形态参数变化程度的差异, 对于拉伸前后存在统计差异的腘绳肌形态结构参数, 使用双因素方差分析比较其变化率在不同腘绳肌及不同拉伸方式之间的差异,采用Bonferroni 法进行检验。
2 研究结果
2.1 不同腘绳肌间的形态差异
三因素方差分析结果显示, 动态拉伸及静态拉伸前不同腘绳肌之间均存在肌束长度(P<0.001)与肌肉厚度(P<0.001)的差异(表1)。 两两比较结果显示: 动态拉伸及静态拉伸前BFLH (P<0.001)与SM 的肌束 (P<0.001) 均短于ST,BFLH 肌束长于SM(P<0.001);动态拉伸前,BFLH 和ST 的肌肉厚度均小于SM (BFLH 与SM:P=0.003;ST 与SM:P<0.001),ST 肌肉厚度小于BFLH (P<0.001);静态拉伸前,BFLH 与ST 的肌肉厚度均小于SM(BFLH 与SM:P=0.001;ST 与SM:P<0.001),ST 肌肉厚度小于BFLH(P<0.001)。
表1 不同腘绳肌间的结构参数比较(±SD)Table1 Comparison of morphological parameters among three hamstrings(±SD)
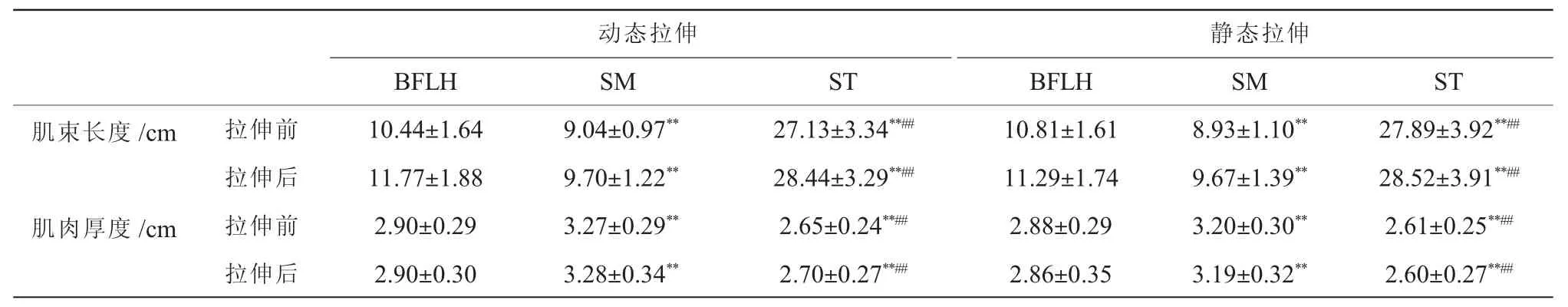
表1 不同腘绳肌间的结构参数比较(±SD)Table1 Comparison of morphological parameters among three hamstrings(±SD)
注:** 表示与BFLH 比较差异具有极显著性,P<0.01;## 表示与SM 比较差异具有极显著性,P<0.01。
动态拉伸静态拉伸BFLHSMSTBFLHSMST肌束长度/cm拉伸前10.44±1.649.04±0.97**27.13±3.34**##10.81±1.618.93±1.10**27.89±3.92**##拉伸后11.77±1.889.70±1.22**28.44±3.29**##11.29±1.749.67±1.39**28.52±3.91**##肌肉厚度/cm拉伸前2.90±0.293.27±0.29**2.65±0.24**##2.88±0.293.20±0.30**2.61±0.25**##拉伸后2.90±0.303.28±0.34**2.70±0.27**##2.86±0.353.19±0.32**2.60±0.27**##
在拉伸前, 各腘绳肌的肌束长度均无拉伸方式间差异(P 分别为0.484、0.761、0.523),各腘绳肌肌肉厚度均无拉伸方式间差异(P 分别为0.836、0.480、0.605), 表明各腘绳肌的肌束长度与肌肉厚度在两种拉伸前的基线水平一致。
动态拉伸及静态拉伸后不同腘绳肌之间均存在肌束长度(P<0.001)与肌肉厚度(P<0.001)的差异(表1)。 两两比较结果显示:动态拉伸及静态拉伸后BFLH 与SM 的肌束均短于ST(P<0.001),BFLH 肌束长于SM (P<0.001); 动态拉伸及静态拉伸后,BFLH 与ST 的肌肉厚度均小于SM (P<0.001),ST肌肉厚度小于BFLH(P<0.001)。
2.2 拉伸造成的腘绳肌急性形态变化
在动态拉伸前后,BFLH(P<0.001)、SM(P<0.001)与ST(P <0.001)的肌束长度均增大,但BFLH(P=0.953)、SM(P=0.802)与ST(P=0.056)的肌肉厚度并无变化(表2)。 在静态拉伸前后,BFLH(P=0.005)、SM(P<0.001)与ST(P=0.044)的肌束长度均增大,但BFLH(P=0.463)、SM(P=0.749)与ST(P=0.603)的肌肉厚度无变化(表2)。
表2 腘绳肌在拉伸前后的结构参数比较(±SD)Table2 Comparison of morphological parameters of hamstrings before and after stretching(±SD)
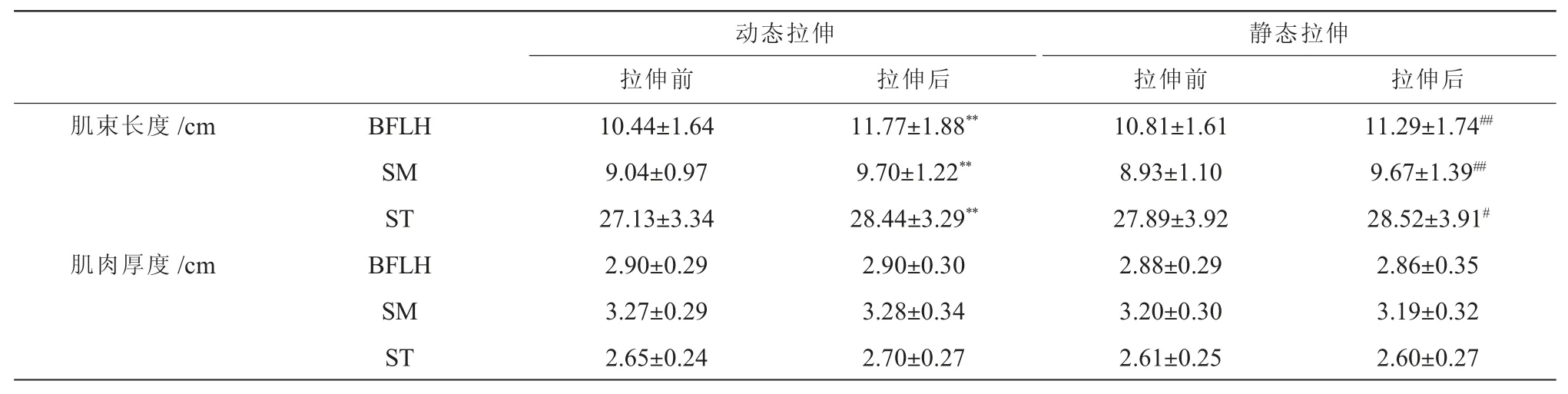
表2 腘绳肌在拉伸前后的结构参数比较(±SD)Table2 Comparison of morphological parameters of hamstrings before and after stretching(±SD)
注:** 表示与动态拉伸前比较差异具有显著性,P<0.01;# 表示与动态拉伸前比较差异具有显著性,P<0.05;## 表示与静态拉伸前比较差异具有极显著性,P<0.01。
动态拉伸静态拉伸拉伸前拉伸后拉伸前拉伸后肌束长度/cmBFLH10.44±1.6411.77±1.88**10.81±1.6111.29±1.74##SM9.04±0.979.70±1.22**8.93±1.109.67±1.39##ST27.13±3.3428.44±3.29**27.89±3.9228.52±3.91#肌肉厚度/cmBFLH2.90±0.292.90±0.302.88±0.292.86±0.35 SM3.27±0.293.28±0.343.20±0.303.19±0.32 ST2.65±0.242.70±0.272.61±0.252.60±0.27
2.3 不同拉伸对腘绳肌形态影响的对比
由于腘绳肌肌肉厚度在拉伸前后不存在统计差异, 仅对腘绳肌肌束延长率进行了不同腘绳肌及不同拉伸方式之间的比较(表3)。 在动态拉伸及静态拉伸作用下, 不同腘绳肌之间均存在肌束延长率的差异(P<0.001)。两两比较结果显示,动态拉伸作用下,BFLH 和ST 的肌束延长率均与SM 无差异(BFLH 与SM:P=0.056;ST 与SM:P=0.852),BFLH的肌束延长率大于ST (P<0.001); 静态拉伸作用下,SM 和ST 的肌束延长率均与BFLH 无差异(BFLH 与SM:P=0.304;ST 与SM:P=0.836),而SM的肌束延长率大于ST(P=0.018)。此外,相比静态拉伸,动态拉伸作用下BFLH 肌束延长率更高(P<0.001),而SM 与ST 肌束延长率不存在拉伸方式间的差异(SM:P=0.604;ST:P=0.582)。
表3 不同拉伸方式作用下的腘绳肌肌束延长率(±SD)Table3 The elongation rate of fascicle length of hamstrings with the effect of different one-time stretching methods(±SD)
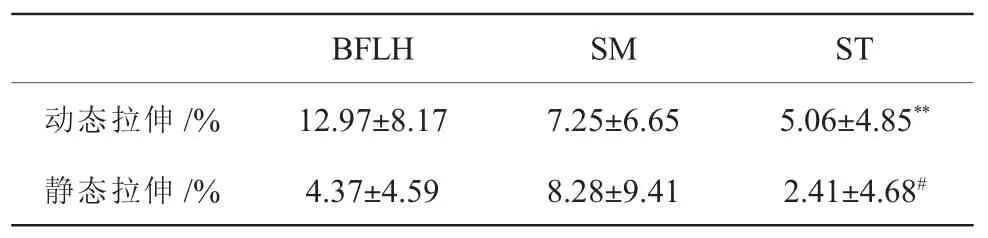
表3 不同拉伸方式作用下的腘绳肌肌束延长率(±SD)Table3 The elongation rate of fascicle length of hamstrings with the effect of different one-time stretching methods(±SD)
注:** 表示与BFLH 比较差异具有极显著性,P<0.01;# 表示与SM 比较差异具有显著性,P<0.05。
BFLHSMST动态拉伸/%12.97±8.177.25±6.655.06±4.85**静态拉伸/%4.37±4.598.28±9.412.41±4.68#
3 分析与讨论
3.1 不同腘绳肌间的形态差异
本研究参考Bennett 等[14]与薛黔等[15]研究所报道的方法对SM、ST 与BFLH 的全景超声图像进行局部划分与测算,发现SM、ST 与BFLH 间存在形态差异,且三者间形态差异在拉伸后依然存在。其中肌束长度大小依次为ST>BFLH>SM,肌肉厚度大小依次为SM>BFLH>ST。 这些发现与前人[1,15]的尸体解剖研究结果相符。 然而,在ST 的全景超声图像中未能发现明显存在的羽状角度 (肌束与腱膜间的夹角),ST 的形态类似长梭状肌, 而非如同SM 与BFLH 的羽状肌。 因此本研究仅涉及三条腘绳肌的肌束长度与羽状角度, 不对羽状角度进行分析。 此外,与前人[15]发现一致,本研究观察到ST 的肌腹由中部开口向远端的不规则“V”字形腱划分为两个串联的肌腹,ST 与单个肌腹的SM 和BFLH 构造迥异, 这或许是挖掘不同腘绳肌生理功能差异来源的重要切入点。
3.2 一次性拉伸造成的腘绳肌形态变化
本研究证实一次性静态拉伸与一次性动态拉伸均可使腘绳肌的肌束长度增大, 但不会改变腘绳肌的肌肉厚度。 静态拉伸可增大腘绳肌柔韧性[18-19],但能否改变腘绳肌肌结构, 相关研究所得出的结论并不统一[6-7]。 对比这些研究可发现,拉伸强度过低、组间间歇时间过长会使得骨骼肌在经受一次性及长期静态拉伸后不发生结构变化, 而拉伸强度过大则易使参与者产生运动损伤。因此,本研究的一次性拉伸干预动作均由参与者主动完成“个人能够达到的、可忍受疼痛感的最大拉伸幅度”,并设置了5 组拉伸动作与较短的间歇时间, 结果证明严格的一次性静态拉伸可使所有腘绳肌的肌束长度明显延长。 在动态拉伸中, 有控制的关节活动使目标肌肉在不完全放松且保持主动张力的状态下受到牵拉[10]。 因此动态拉伸中目标肌肉在进行以部分体重为阻力的离心-向心工作,而离心运动可以增大腘绳肌柔韧性[19]与骨骼肌的肌束长度[20],则解释了本研究中一次性动态拉伸引起腘绳肌肌束长度明显延长的现象。 Lima等[6]与Freitas 等[7]的研究结果均显示拉伸无法对BFLH 的肌肉厚度产生影响, 本研究进一步证实了拉伸不能使包括BFLH 在内的三条双关节腘绳肌发生肌肉厚度的改变。
拉伸后腘绳肌肌束 (由多条肌纤维并列组合而成)延长归因于肌纤维内肌节长度的增加。当肌节长度越接近自身最适初长度时, 肌节内横桥结合数目越多,肌肉收缩力越强。力量训练引起的腘绳肌肌力提升往往伴随着腘绳肌的结构变化[21-28],而拉伸引起的腘绳肌内部结构变化与功能变化间也存在关联[29]。本研究发现不同拉伸方式造成的BFLH 肌束延长程度不一, 这提示在单次运动训练之初的热身阶段选择不同类型拉伸方式, 可能对后续训练过程中的BFLH 的收缩力产生不同程度的影响。 Barbosa 等[30]发现静态拉伸会引起腘绳肌离心力量及下肢爆发力水平的下降, 而动态拉伸却不会影响腘绳肌离心力量及下肢爆发力, 这表明过多的静态拉伸会降低腘绳肌的力量, 不利于人体在拉伸后完成需要腘绳肌参与的抗阻动作, 但动态拉伸却不会有这样的不利影响。 往后研究可基于本研究的发现进一步探究一次性拉伸引起的腘绳肌肌束长度的变化与肌节最适初长度间的关系, 更有力地解释拉伸对腘绳肌运动功能的影响。
本研究证实了一次性动态拉伸作用下的BFLH肌束延长率高于一次性静态拉伸。此现象与Kellis[31]所提出的 “动态拉伸可能比静态拉伸更有助于恢复BFLH 顺应性”猜想一致。 本研究参与者主观反馈显示,相比动态拉伸,尽个人最大努力自主完成的静态拉伸过程会造成更剧烈、持续的不适感。 这可能使得参与者的实际静态拉伸强度与自身预计强度不符,导致静态拉伸后BFLH 的肌束延长率相比动态拉伸低。
3.3 拉伸后不同腘绳肌形态变化的对比
本研究发现SM、ST 与BFLH 在拉伸后的肌束延长率不同, 一次性动态拉伸使ST 的肌束延长率低于BFLH,一次性静态拉伸使ST 的肌束延长率低于SM, 这应当与三条腘绳肌在拉伸时所受牵拉程度不同有关。然而相比BFLH 与SM,ST 在两种拉伸后的肌束延长率均为最低的现象似乎与前人研究结果存在冲突。 Arnold 等[32]使用核磁共振设备进行测量后发现, 在三条双关节腘绳肌中,ST 在髋膝关节处的力臂均为最长,BFLH 在髋关节处的力臂长度大于SM, 而在膝关节处的力臂长度是三条肌肉中最短的,而SM 在膝关节处的力臂长于BFLH,但在髋关节处力臂最短。 ST 的关节力臂最大[33],在髋膝关节的伸展过程中也延长更大[34],ST 理应受到更剧烈的牵拉从而产生明显的肌束延长效果。ST 在拉伸后肌束延长率最低的现象可能与ST 具有三条肌肉中最长最细的远段游离腱有关[35],形态细长的肌腱往往具有较强的顺应性[36],能在拉伸时承担很大比例的应变,从而减轻肌束受到的牵拉。 Dolman 等[37]将短跑冲刺过程中的运动学特征与不同关节角度下的腘绳肌尸体解剖计算结果相结合, 通过计算局部组织位移从而推算并比较了三条腘绳肌在运动中的受力,结果显示在腘绳肌离心收缩时,BFLH 需要对抗的阻力比其他腘绳肌更大。 Pollard 等[20]的研究证明, 阻力水平更大的离心力量训练可使腘绳肌肌束在训练后的延长程度更大。 这也可部分解释本研究中一次性动态拉伸使BFLH 的肌束延长率高于ST的现象。 本研究中未发现拉伸会使SM 与BFLH 之间存在肌束延长率的差异, 这一现象需要结合二者在髋膝关节处的力臂与拉伸动作中的拉伸力作进一步分析才能得到解释。 SM、ST 与BFLH 在拉伸作用下肌束延长程度不同, 这表明同样的拉伸动作对三条腘绳肌柔韧性的改善作用存在差异。
4 结论
相比一次性静态拉伸, 一次性动态拉伸可更明显地改变BFLH 结构。 ST 与SM 的结构均可被两种拉伸方式改变, 但二者结构变化程度均无拉伸方式间的差别。三条腘绳肌之间存在结构差异,且在拉伸作用下发生了不同程度的结构变化。
不同一次性拉伸对多个腘绳肌形态结构存在急性影响的发现, 可为单次运动训练中选择腘绳肌拉伸类型提供理论参考, 并为进一步探索多种拉伸方式对腘绳肌内部组织功能特性的影响提供部分研究依据。
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23
云南画报(2021年8期)2021-11-13
数学小灵通(1-2年级)(2020年9期)2020-10-27
阅读(低年级)(2019年4期)2019-05-20
现代营销(创富信息版)(2018年10期)2018-10-12
作文大王·低年级(2017年11期)2017-12-05
小学生学习指导(低年级)(2017年12期)2017-11-22
华人时刊(2016年13期)2016-04-05
读写算(上)(2015年6期)2015-11-07
电子设计工程(2015年15期)2015-02-27
