
How do lateral septum projections to the ventral CA1 influence sociability?
2024-02-11 10:02DanWangDiZhaoWentaoWangFengaiHuMinghuCuiJingLiuFantaoMengCuilanLiuChangyunQiuDunjiangLiuZhichengXuYamengWangYuZhangWeiLiChenLi
中国神经再生研究(英文版) 2024年8期
Dan Wang, Di Zhao, Wentao Wang, Fengai Hu, Minghu Cui, Jing Liu, Fantao Meng,Cuilan Liu, Changyun Qiu, Dunjiang Liu, Zhicheng Xu, Yameng Wang, Yu Zhang, Wei Li,Chen Li,*
Abstract Social dysfunction is a risk factor for several neuropsychiatric illnesses.Previous studies have shown that the lateral septum (LS)-related pathway plays a critical role in mediating social behaviors.However, the role of the connections between the LS and its downstream brain regions in social behaviors remains unclear.In this study, we conducted a three-chamber test using electrophysiological and chemogenetic approaches in mice to determine how LS projections to ventral CA1 (vCA1) influence sociability.Our results showed that gamma-aminobutyric acid (GABA)-ergic neurons were activated following social experience, and that social behaviors were enhanced by chemogenetic modulation of these neurons.Moreover, LS GABAergic neurons extended their functional neural connections via vCA1 glutamatergic pyramidal neurons, and regulating LSGABA→vCA1Glu neural projections affected social behaviors, which were impeded by suppressing LSprojecting vCA1 neuronal activity or inhibiting GABAA receptors in vCA1.These findings support the hypothesis that LS inputs to the vCA1 can control social preferences and social novelty behaviors.These findings provide new insights regarding the neural circuits that regulate sociability.
Key Words: chemogenetics; GABA receptor; GABAergic neurons; glutamatergic neurons; lateral septum; neural excitability; neural projection; social novelty;social preference; ventral CA1
Introduction
Social behavior is deeply linked to quality of life in that it is required to maintain physical functioning and independently adapt to one′s social environment (Chen and Hong, 2018; Liu et al., 2020).Indeed, social dysfunction is not only considered to be a salient symptom, but is recognized as a critical risk factor for several neuropsychiatric illnesses.These include depression (Achterbergh et al., 2020; Lei et al., 2022), schizophrenia(Savolainen et al., 2019), and autism spectrum disorder (Sissons et al., 2022).In particular, patients with depression face persistent interpersonal difficulties,which may increase vulnerability regarding the onset and recurrence of depression (Yang et al., 2022).Therefore, understanding the neurobiological mechanisms that mediate social function is crucial when exploring potential therapeutic strategies for treating various psychiatric disorders.However, our knowledge about the neural circuits that regulate adult social interactions is limited.
The lateral septum (LS) is a limbic nucleus located in the subcortical region of the forebrain.It essentially comprises gamma-aminobutyric acid (GABA)-ergic neurons (Sheehan et al., 2004; Zhao et al., 2013), and is considered to be a hub that regulates various social behaviors (including social novelty,communication, recognition, parental behavior, and aggression) (Menon et al., 2022; Walsh et al., 2023) through its diverse input and output connections with several regions of the brain, including the hippocampus, medial prefrontal cortex, hypothalamus, and striatum (Shin et al., 2018; Borland et al., 2020; Clemens et al., 2020; Horiai et al., 2020; Kelly et al., 2020; Rizzi-Wise and Wang, 2021).A recent study using both environmental and genetic animal models of autism spectrum disorder demonstrated that the activation of neurons expressing the oxytocin receptor in the LS and projecting to the hippocampal CA1 region ameliorated impaired social memory (Horiai et al., 2020).Furthermore, the activation of neurons that projected from the LS to the ventromedial area of the hypothalamus strongly suppressed aggression in male mice (Wong et al., 2016).The activation of GABA type A (GABAA) receptors in the dorsal LS increased aggressive behavior in male and female hamsters, while social housing markedly decreased the ability of GABAAreceptor activation to exert an aggressive phenotype in male but not female hamsters (Borland et al., 2020).This suggests that LS-associated circuitry is crucial in mediating social behaviors.Moreover, this circuitry could serve as a therapeutic target for treating deficits related to social behaviors in some neuropsychiatric illnesses.However, the precise functional roles of connections between the LS and downstream regions, particularly the majority of GABAergic neuronal projections, are poorly understood with respect to social behavior, necessitating further investigation.
Increasing evidence has shown that the ventral CA1 (vCA1) receives and integrates different streams of information and is involved in hippocampusrelated social behaviors.For example, projections from vCA1 pyramidal neurons to the nucleus accumbens shell or medial prefrontal cortex play an essential role in social memory (Okuyama et al., 2016; Phillips et al., 2019;Rizzi-Wise and Wang, 2021), while neural projections from the amygdala to the ventral hippocampus modulate bidirectional social interactions (Felix-Ortiz and Tye, 2014).In a recent study, specific glutamatergic projections from the vCA1 and ventral CA3 to the LS facilitated approach and avoidance behaviors,respectively (Yeates et al., 2022).Additional investigations are required to clarify the role of other neuronal projections innervating vCA1 in social behaviors.
The purpose of the present study was to evaluate neural excitation in the LS after social interactions and social behaviors following the suppression or activation of LS GABAergic neurons via a chemogenetic strategy, and to determine the role of GABAergic LS-vCA1 projections and neural connection mechanisms in social behaviors.
Methods
Animals
We obtained male C57BL/6J mice (RRID: SCR_007053; Stock No.000664;aged 8–10 weeks, weighing 20–25 g;n= 10) and transgenic mice (Vgat-ires-Cre and Fos-CreERT2mice: respective Stock Nos.028862 and 021882; aged 8–10 weeks, weighing 20–25 g;n= 5) from Jackson Laboratory (Bar Harbor,ME, USA) (Vong et al., 2011; Guenthner et al., 2013) and maintained them in breeding colonies.We reared the animals in regulated environmental conditions (room temperature: 22–24°C; 12-hour light/dark cycle, lights switched on at 7 a.m.) with unlimited access to food and water.We used the following polymerase chain reaction primers for genotyping: Vgat-ires-Cre mice: forward: 5′-CAC CAG TGT CTA CCC CTG GA-3′, reverse: 5′- CGC GCC TGA AGA TAT AGA AGA-3′; and Fos-CreERT2mice: forward: 5′- CTT CGT CAT CGG CGG CAT CTG-3′ and reverse: 5′-CCA AAA GAC GGC AAT ATG GT-3′.
Binzhou Medical University Hospital′s Institutional Animal Care and Use Committee approved all protocol for animal experiments (approval No.20191108-30) on November 8, 2019.The study procedure adhered to the guidelines of the UK Animals (Scientific Procedures) Act of 1986 as well as associated guidelines in the EU Directive for animal experiments [2010/63/EU].
Drugs
Clozapine-N-oxide (CNO; PubChem CID: 135445691) was supplied by Sigma-Aldrich (St.Louis, MO, USA).CNO was mixed with sterile saline at a concentration of 0.5 mg/mL.This was then diluted by adding saline to obtain a final concentration of hM3D (0.3 mg/kg) or hM4D (2 mg/kg).This was used for intraperitoneal injection prior to transcardial perfusion or behavioral tests,based on an earlier report (Zhao et al., 2022).
Tamoxifen (PubChem CID: 2733526) was provided by Sigma-Aldrich, and dissolved in ethanol.After supplementing this solution with corn oil from Sigma-Aldrich, the mixture was shaken, incubated at 37°C, and concentrated via ethanol volatilization under a fume hood to achieve a final concentration of 5 mg/mL.It was then kept in a dark environment at –20°C.Bicuculline(PubChem CID: 124203297) was obtained from Sigma-Aldrich and dissolved in sterile saline (0.1 µg /µL, dose: 0.1 µg/mouse).
Adeno-associated virus
Shanghai Hanbio Biotechnology Co., Ltd.(Shanghai, China) provided the following Adeno-associated viruses (AAVs): AAV2/9-hSyn-DIO-hM3D(Gq)-mCherry (AAV-DIO-hM3D-mCherry), AAV2/Retro-CAG-GFP (AAV-Retro-GFP),AAV2/9-hSyn-DIO-hM4D(Gi)-mCherry (AAV-DIO-hM4D-mCherry), AAV2/Retro-CAG-Cre-GFP (AAV-Retro-Cre-GFP), AAV2/9-hSyn-DIO-mCherry (AAVDIO-mCherry), and AAV2/9-CAG-DIO-GFP (AAV-DIO-GFP), with titers > 1 × 1012viral genomes/mL.BrainVTA (Wuhan) Co., Ltd.(Wuhan, China) provided the following AAVs: AAV-Calcium–calmodulin (CaM)-dependent protein kinase II (CaMKII) -DIO-hM4D(Gi)-mCherry-WPRE-hGHpolyA and AAV2/1-Vgat-Cre-WPRE-hGHpA (AAV2/1-Vgat-Cre) with titers > 1 × 1012viral genomes/mL (Zhao et al., 2022).
Stereotaxic surgery, microinjection, and cannulation
After inducing anesthesia via an intraperitoneal injection of sodium pentobarbital (40 mg/kg, P3761, Sigma-Aldrich), the mice were aligned on a stereotaxic device (David Kopf, Tujunga, CA, USA).Mouse body temperature was regulated using a temperature controller (RWD Life Sciences, Shenzhen,China).The surgical procedure was as previously reported (Liu et al., 2022; Li et al., 2023).A glass pipette and a Micro 4 infusion pump (WPI, Sarasota, FL,USA) were used to infuse AAV into the chosen brain areas with the following coordinates.LS: antero-posterior (AP), mediolateral (ML), and dorsoventral(DV) = +0.5 mm, ±0.40 mm, and –3.20 mm from bregma, respectively; total volume: 0.6 µL (0.30 µL/side); flow rate: 0.10 µL/min; vCA1: AP, ML, and DV = –3.16 mm, ±3.2 mm, and –4.15 mm from bregma, respectively; total volume: 0.80 µL (0.40 µL/side); flow rate: 0.10 µL/min.After infusion, we kept the needle in place for 5 minutes to allow diffusion and prevent backflow.For postoperative analgesia, we administered ketoprofen (5 mg/kg, Sigma-Aldrich) intraperitoneally.At 21 days after injecting the AAV, we conductedin vivoextracellular recordings or behavioral tests.
The vCA1 cannula was implanted as reported previously (Phillips et al., 2019).We secured male C57BL/6 adult mice on a stereotaxic frame (David Kopf)after induction of anesthesia.The surface of the skull was then covered with a phosphoric acid gel etchant provided by Kerr (Orange, CA, USA).Next, we inserted a bilateral guide cannula into vCA1 (coordinates: AP, ML, and DV =–3.16 mm, ±3.2 mm, and –3.15 mm, respectively).The surfaces of the skull and cannula were then coated with an adhesive supplied by GLUMA (Hanau,Germany).The top surface was brushed with resina fluida (Filtek Z350 XT 3 M, USA) and subjected to a 45-second light curing period with a curing light system (VALO, South Jordan, UT, USA) acquired from Ultradent Products.The cannula was sealed with dental cement, and obstruction of the cannula was avoided via insertion of a dummy needle into the guide cannula.Following completion of surgery, each animal was kept in an individual cage for the entire recovery period, which was 21 days with routine management.During the microinjection procedure, the mice were kept conscious, and their movement in their home cages was unrestrained.Subsequently, a stainless steel injector (33-G) with a syringe (5 µL) was inserted into the guide cannula;the injector was extended to 1 mm beyond the guide cannula′s tip.Next,bilateral infusion of vehicle, CNO (0.5 µL per side; 5 µM) or Bicuculline (0.5 µL per side; 0.1 µg /µL) was performed for 2.5 minutes.To prevent backflow, we leftthe injector tipin situfor an additional 5 minutes following completion of the infusion.The mice were subjected to behavioral tests 45 minutes after the microinjection procedure.Mice that received “non-targeted” injections (1–2 mice with missed injections or 2–3 mice with loose or missed canulations per group) were omitted from the statistical analyses (Zhao et al., 2022).
Cell counting and immunofluorescence histochemistry
After anesthesia via an intraperitoneal injection of sodium pentobarbital,as above, transcardial perfusion was performed using ice-cold 4%paraformaldehyde, followed by removal of the brains.The brains were then post-fixed overnight at 4°C in 4% paraformaldehyde and transferred to a sucrose solution (30%) for 2 days.Subsequently, we cut 40-µm-thick coronal sections using a freezing microtome.After washing the tissue three times in 0.01 M phosphate buffer solution (PBS), we used 1% bovine serum albumin(diluted by 0.01 M PBS with 3% Triton X-100 and 0.3% goat serum) to block the brain sections at room temperature for 1 hour.All sections were kept at 4°C overnight in a blocking solution containing the following primary antibodies:rabbit anti-c-FBJ murine osteosarcoma viral oncogene homolog (Fos; 1:1000,CST, Danvers, MA, USA, Cat# 2250S, RRID: AB_2247211), rabbit anti-∆FosB(1:1000, CST, Cat# 2251S, RRID: AB_2106903), rabbit anti-GABA (1:500,Sigma-Aldrich, Cat# A2052, RRID: AB_477652), and mouse anti-vesicular glutamate transporter 2 (Vglut2; 1:400, Abcam, Cambridge, UK, Cat# ab79157,RRID: AB_1603114).After washing the sections three times in 0.01 M PBS,we incubated them at room temperature for 4 hours in a dark environment with the following secondary antibodies: Alexa Fluor® 488 donkey anti-rabbit IgG (1:400, Invitrogen, Carlsbad, CA, USA, Cat# A21206, RRID: AB_2535792),Alexa Fluor® 546 goat anti-rabbit IgG (1:400, Invitrogen, Cat# A11035, RRID:AB_2534093), and Alexa Fluor® 546 goat anti-mouse IgG (1:400, Invitrogen,Cat# A11030, RRID: AB_2534089).The stained sections were then washed three times in 0.01 M PBS.We used Poly-L-lysine-coated glass slides to wetmount the stained sections.This was followed by counterstaining with a fluorescent mounting medium.The slides were then covered with cover slips.A confocal laser-scanning microscope (Olympus FV1000; Shinjuku,Tokyo, Japan) was used to visualize the images.The immunofluorescence positive cells in 5–6 sections per brain containing the whole LS (approximately 0.14–1.18 mm with reference to bregma) or vCA1 (approximately –2.8 to–3.4 mm with reference to bregma) were counted manually by an observer with no awareness of the treatment group (Li et al., 2021a).The results were expressed as the average number of cells per slice.
Whole-cell patch-clamp recordings
We subjected adult male mice (aged 8–10 weeks) to inhalation anesthesia via isoflurane (5 mL/kg, Sigma-Aldrich).After collecting the brains, we rapidly transferred them to an ice-cold solution for sectioning.The sectioning solution contained 3 mM KCl, 254 mM sucrose, 2 mM CaCl2, 10 mM D-glucose,2 mM MgCl2, 24 mM NaHCO3, and 1.25 mM NaH2PO4.Next, 200-µm-thick coronal slices including the LS were prepared using a vibrating microtome(Leica VT1200, Wetzlar, Germany).We transferred the brain slices to artificial cerebrospinal fluid solution (2 mM KCl, 124 mM NaCl, 1.25 mM NaH2PO4,26 mM NaHCO3, 2 mM MgSO4, 2 mM CaCl2, 0.4 mM vitamin C, and 10 mM D-dextrose) saturated with carbon dioxide and oxygen (5% CO2/95% O2).Recovery of the slices was performed at 30°C for at least 1 hour.During whole-cell recording, we continuously perfused the brain slices with the above-mentioned cerebrospinal fluid solution.We used a Sutter P-97 micropipette puller to fabricate whole-cell recording glass electrodes with atip resistance of 4–7 mΩ from borosilicate glass capillaries (thin-walled; TW-150F; World Precision Instruments).The electrodes were filled with 120.0 mM potassium gluconate, 20.0 mM KCl, 2.0 mM MgCl2, 10.0 mM HEPES,0.25 mM GTP, 2.0 mM ATP, and 0.1 mM EGTA, and had an osmotic pressure of 295–300 mOsm; pH 7.4.After the cell membrane had disintegrated and was allowed to stabilize for approximately 5 minutes, current was injected in 10-pA steps.Glutamatergic neurons were differentiated from GABAergic neurons based on the firing frequency and waveform (Magnin et al., 2019;Wang et al., 2021).To record glutamatergic neurons in vCA1, we switched from a voltage clamp to a current clamp.The current was maintained such that the resting membrane potential was close to –70 mV.The recording time was set to 500 ms.The input resistance was subsequently switched to record the super current.We measured the average levels of the current amplitude,firing frequency, threshold, and half-wave width of the first peak current after each injected current.To record GABAergic neurons in vCA1, we set the holding current at –45 mV.LS GABAergic neurons were recorded according to a previous report (Wang et al., 2021).We used pClamp software with a MultiClamp 700B amplifier (Molecular Devices, Sunnyvale, CA, USA) to record clamped GABAergic neurons in the whole-cell mode.A 3 kHz hardware filter(clampfit, Molecular Devices, Sunnyvale, CA, USA) was used to collect the data.The neuronal firing properties were determined via a whole-cell current clamp at 0 mV, and the data were collected after 3 minutes of stabilization.The clampfit was used to count the firing frequency and action potential (AP)amplitude.To determine the effect of CNO on neurons expressing hM4D or hM3D, the slices were kept in artificial cerebrospinal fluid containing 10 µM CNO for approximately 2 hours prior to the beginning of recording (Zhou et al., 2019).
Home cage social interaction task and the acute restraint stress paradigm
For social interaction test, the subject mice were single housed for 2-3 days,then, a stranger mouse was introduced, these two mice were allowed to freely explore and interact in the home cage for 2 hours only once (acute task), or 30 minutes per day for 5 days (chronic task).
For the acute restraint stress, the experimental mice were habituated for 2–3 hours in individual homecages.Restraint stress was performed by placing each mouse into a 50 mL polystyrene tube with ventilation for 30 minutes.
Behavioral procedures
We performed behavioral measurements during the later part of the light cycle.Prior to testing, all animals were acclimated to the behavior room for at least 2 hours.Investigators who were unaware of the treatment groups scored the mouse behaviors.
Three-chamber test
To assess social interaction, we conducted the three-chamber test in a 60 cm × 40 cm × 40 cm rectangular plexiglas arena, according to previously published studies (Rein et al., 2020; Rodriguez et al., 2023) with minor modifications.The arena was segregated into three chambers (each chamber was 20 cm × 40 cm × 40 cm) via dividing walls, and there was a doorway (13 cm width) that enabled mice to access each chamber.We allowed the mice to habituate to the test room for 2 hours before the experimental trials.Initially,the doorways to the side chambers were closed, and the subject mouse was placed in the center chamber.This mouse was allowed to freely explore the center chamber for 5 minutes.In the first test session, an empty enclosure was placed in each of the side chambers, and the doors to these chambers were opened.The test mouse was introduced in the center chamber, and allowed to freely explore the two empty enclosures for 10 minutes.Then, the subject mouse was restricted to the center chamber because we closed both doorways.In the social preference phase, a stranger mouse (designated as stranger-1) was randomly placed in one of the two wire cages, while the other chamber remained with empty wire cage object chamber.The doorways were then opened.The subject mouse was allowed to freely explore the three chambers for 10 minutes.We then conducted an additional social novelty trial, which was 10 minutes long.We placed a new stranger mouse(designated as stranger-2) inside the previously empty wire cage.At this point,stranger-1 was now familiar to the subject mouse.When all three phases were complete, we cleaned the wire cages and chambers using 20% ethanol to eliminate any cues that could influence the behavioral responses of the subsequent mice.The times taken to explore the chambers as well as the duration of sniffing were analyzed using an EthoVision XT v11.5 video tracking system (Noldus Information Technology, Wageningen, The Netherlands).We defined the social interaction index as the ratio of the total amount of time spent in the two chambersversusthe time spent in the chamber containing stranger-1 in the social preference phase or that containing stranger-2 in the social novelty phase.We defined the discrimination index as the ratio of the sniffing duration towards stranger-1 in the social preference phase or towards stranger-2 in the social novelty phase to the total sniffing time.A stimulus interaction was considered to have occurred when the nose of the experimental subject was directed toward an enclosure at a distance < 2 cm.We calculated the time engaged in each interaction.
Open field test
We conducted an open field test using a 60 cm × 60 cm × 40 cm square arena under uniform illumination conditions as previously described (Li et al., 2021a).A mouse was introduced to the centre of the arena and allowed to freely move for 5 minutes.We placed a video camera above the arena to record mouse movement.For analysis purposes, we divided the arena into a 3 × 3 grid with 9 uniform squares.We designated the central square as the central zone.We estimated the time spent in the central zone as well as the total distance covered in the arena using ANY-maze software from Stoelting Co.(Wood Dale, IL, USA).Based on the overall motor activity in the test, the total distance covered was determined.
Locomotor activity
Mice were placed in a 40 cm × 40 cm × 30 cm open field cage (SuperFlex Fusion; Omnitech Electronics Inc., Columbus, OH, USA) in a well-illuminated room, and allowed to freely explore for 30 minutes.Mice activity was recorded, and we estimated the total distance covered using Fusion software provided by Omnitech Electronics Inc.(Meng et al., 2020; Sun et al., 2023).
Statistical analysis
The sample sizes were determined based on previous publications (Li et al.,2021a, b; Zhao et al., 2022).There were no changes in group sizes during the experiments.The mean ± standard error of the mean (SEM) was used to present all experimental data.The data were analyzed using GraphPad Prism version 8.0.1 for Windows (GraphPad Software, San Diego, California USA, www.graphpad.com).We tested the equal variance assumption and normality using theF-test and Shapiro-Wilk test, respectively.Differences between the experimental groups were evaluated using the Mann-WhitneyUtest (for data with a non-normal distribution), two-tailed unpairedt-test with Welch’s correction (for data with a normal distribution and unequal variance), and two-tailed unpairedt-test (for data with a normal distribution and equal variance).We used a one-way analysis of variance with Sidak’s test to analyze differences among three groups.For more than three groups, we used a two-way repeated-measures analysis of variance with Sidak’s test.We used aPvalue of < 0.05 to represent statistical significance.Data acquisition and analysis were conducted by investigators who were blinded to the experimental treatment groups.
Results
Social interaction activates LS GABA neurons
To determine whether LS neuronal activity was related to social activity,we administered a home cage social interaction task (Figure 1A).This task reflected a social experience in a familiar environment (Kim et al., 2019).At 30 minutes after an acute social interaction (2 hours), we measured the number of neurons in the LS labeled with neuronal activity biomarker c-Fos-immunoreactive proteins.We found that the number of c-Fospositive neurons in the LS of mice interacting with a new target mouse was significantly higher than that of mice that did not interact with a new target mouse (t(8)= 4.0720,P= 0.0036; Figure 1B).This finding indicates that LS neuronal activity is involved in acute social interaction.Given that lateral septal neurons are predominantly GABAergic neurons (more than 90%) (Zhao et al., 2013), we conducted double immunofluorescence histochemistry to determine whether the c-Fos-positive neurons were co-localized with GABAergic neurons.In mice interacting with or without a target mouse, colocalization with LS GABAergic neurons was observed for most c-Fos-positive neurons (91.5 ± 1.3% and 90.8 ± 1.2%), with no obvious difference between the groups (t(10)= 0.4118,P= 0.6892; Figure 1C).This finding indicates that LS GABAergic neuronal activity might be critically involved in social interaction.We performed an electrophysiological assay to confirm the spontaneous firing of GABAergic neurons after an acute social interaction.The recording results revealed that, compared with the control mice, those with a target exhibited an increased frequency of action potentials (P= 0.0349; Mann-WhitneyUtest).However, the action potential amplitude (P= 0.2584; Mann-WhitneyUtest) and membrane potential (unpairedt-test with Welch’s correction,P=0.6208) remained constant (Figure 1D).
We further confirmed the association between GABAergic neuronal activity and social interaction by conducting a chronic home cage social interaction task.In this task, mice interacted with a social target mouse in their home cage for 30 minutes per day on five consecutive days (Figure 1E).At 30 minutes after the end of the last social interaction task, we conducted immunofluorescence detection of ∆FosB, an indicator of neuronal activity that can be induced by various stimuli, and is frequently used in chronic stress/experience paradigms (Nestler, 2015; Wang et al., 2021; Zhao et al.,2021).Specifically, we measured the number of ∆FosB-immunoreactive neurons in the mice LS.We found that social interaction-treated mice had an increased number of ∆FosB-positive neurons in the LS (t(6)= 2.8770,P=0.0282; Additional Figure 1A).We then estimated the percentage of ∆FosBpositive GABAergic neurons among the total number of ∆FosB-positive neurons.Consistent with the results of the acute social interaction task,in mice that interacted with a social target mouse, the majority of ∆FosBpositive neurons in the LS showed co-localization with GABAergic neurons(91.48 ± 2.36%).This value was nearly the same in mice without a social target mouse (90.85 ± 1.23%) (t(4)= 0.2368,P= 0.8244; Additional Figure 1B).The electrophysiological results also validated the social treatmentrelated hyperexcitability of GABAergic neurons, which was characterized by an increased number of action potentials (two-tailed unpairedt-test with Welch′s correction,P< 0.0010), decreased membrane potential (t(31)= 9.9110,P< 0.0010), and no change in the action potential amplitude (t(31)= 0.3143,P= 0.7554) of LS GABAergic neurons in mice interacting with a social target mouse compared with that in mice that did not interact with target mice(Figure 1F).
The inhibition of LS GABA neurons attenuates social behaviors
To investigate how LS GABAergic neuronal activity affects social interaction behaviors, we used Vgat-Cre mice that expressed Cre recombinase in inhibitory GABAergic neuronal cell bodies in many brain regions including the LS, without changes in the expression of endogenous vesicular inhibitory amino acid transporters (Vong et al., 2011).We then bilaterally administered an AAV that expressed Cre-dependent hM4D(Gi) (designated AAV-hM4), as well as a control (mCherry virus, designated as AAV-mCherry) into the LS of Vgat-Cre mice.Three weeks after this treatment, we intraperitoneally injected the mice with CNO to inhibit the activity of LS GABAergic neurons.We then performed the three-chamber test to examine social preference and social novelty behaviors 45 minutes following CNO administration (Figure 2A).During the social preference phase, we found that AAV-hM4-injected mice spent significantly less time exploring the stranger-1 chamber (P= 0.0134),while the amount of time spent exploring the middle chamber (P= 0.6764)and the object chamber remained the same (P= 0.1508) (treatment:F(1,13)=0.0001,P> 0.9999; social squares:F(2,26)= 23.7400,P< 0.0010; interactions:F(2,26)= 4.7080,P= 0.0180).Furthermore, the AAV-hM4-injected mice showed a significantly lower social interaction index than the control mice (AAVmCherry) (t(13)= 2.7940,P= 0.0152).Furthermore, the AAV-hM4-administered mice spent less time sniffing the stranger-1 mouse (P< 0.0010), but not the object (P= 0.9885) (treatment:F(1,13)= 32.8800,P< 0.0010; social squares:F(1,13)= 120.6000,P< 0.0010; interactions:F(1,13)= 45.9900,P< 0.0010), and had a significantly lower discrimination index than the control mice (two-tailed unpairedt-test with Welch’s correction,P= 0.0106; Figure 2B).These results demonstrate that social preference abilities were impaired after LS GABAergic neurons were inhibited.
During the social novelty phase, the AAV-hM4-injected mice spent less time exploring the stranger-2 chamber (P= 0.0079).However, we did not observe a significant difference in the duration of time spent in the middle (P= 0.1924)or familiar mouse (stranger-1) chamber (P= 0.4639) (treatment:F(1,13)=0.0001,P> 0.9999; social squares:F(2,26)= 30.4000,P< 0.0010; interactions:F(2,26)= 5.2080,P= 0.0125).The AAV-hM4 group had a significantly lower social interaction index than the control group (P= 0.0358; two-tailed unpairedt-test with Welch’s correction).Additionally, the AAV-hM4-administered mice spent less time sniffing the new mouse (stranger-2) (P<0.0010)versusthe familiar mouse (stranger-1) (P= 0.4590) (treatment:F(1,13)= 9.4930,P= 0.0088; social squares:F(1,13)= 76.010,P< 0.0010; interactions:F(1,13)= 53.7300,P< 0.0010).The AAV-hM4 group had a significantly lower discrimination index than the control mice (P= 0.0289; Mann-WhitneyUtest) (Figure 2C).Social behavior impairment did not appear to be caused by an alteration in anxiety or locomotion, because AAV-hM4 treatment did not affect the amount of time spent in the centre of the open field test (t(13)= 1.6900,P= 0.1149; Additional Figure 2A) or the distance travelled in the locomotor test (treatment:F(1,13)= 0.6951,P= 0.4195; timepoints:F(14,182)=20.1200,P< 0.0010; interactions:F(14,182)= 0.5086,P= 0.9262; total distance:t(13)= 0.8337,P= 0.4195; Additional Figure 2B).Therefore, LS GABAergic neuron inhibition attenuated sociability as well as social novelty behavior in mice.Hence, we validated the efficiency of hM4D in mediating neural inhibitory activity.We used a 30-minute restraint paradigm to increase basal neural activity, and estimated c-Fos-positive cell numbers 45 min after the CNO injection.AAV-hM4 mice showed a dramatic decrease in the number of c-Fos-positive cells in the LS compared with the control mice (t(4)= 4.4270,P= 0.0115; Figure 2D).Furthermore, according to our electrophysiological results, the AAV-hM4 group showed a decrease in action potential frequency(unpairedt-test with Welch’s correction,P= 0.0012) and no change in action potential amplitude (t(19)= 0.7755,P= 0.4476) compared with the control group (Figure 2E).
LS GABA neuronal activation enhances social behaviors
Next, we explored how LS GABAergic neuronal activation influences social interaction behaviors by bilaterally injecting an AAV expressing Cre-dependent hM3D (Gq) (designated as AAV-hM3) or mCherry virus (designated as AAVmCherry) as a control into the LS of Vgat-Cre mice.CNO was intraperitoneally injected 3 weeks afterwards, thereby activating GABAergic neurons in the LS.At 45 minutes after the injection, we assessed social behaviors via the threechamber test (Figure 3A).In the social preference phase, AAV-hM3-injected mice exhibited no remarkable differences in the amount of time taken to explore the stranger-1 chamber (P= 0.9998), middle chamber (P= 0.9979),or object chamber (P= 0.7705) (treatment:F(1,14)= 0.7656,P= 0.3963; social squares:F(2,28)= 37.4200,P< 0.0010; interactions:F(2,28)= 0.2340,P= 0.7929).The social interaction index of the AAV-hM3 group did not differ from that of the control group (AAV-mCherry) (t(14)= 0.9639,P= 0.3515).We observed AAV-hM3-injected mice showing an increase in the amount of time taken to sniffthe stranger-1 mouse (P= 0.0091), but not the object (P= 0.9854)(treatment:F(1,14)= 4.5750,P= 0.0506; social squares:F(1,14)= 40.8600,P<0.0010; interactions:F(1,14)= 4.9310,P= 0.0434), AAV-hM3-injected mice exhibited a significantly higher discrimination index than the control group(P= 0.0164; Mann-WhitneyUtest; Figure 3B).Thus, activated LS GABAergic neurons appear to promote social preference behaviors.
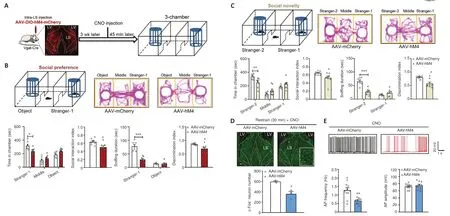
Figure 2 | Inhibition of LS GABA neurons suppressed social behaviors.
In the social novelty phase, the time spent by the AAV-hM3-treated mice to explore the new mouse (stranger-2) (P= 0.2715), middle (P= 0.8408), and familiar mouse (stranger-1) chambers (P= 0.0572) was identical to that in the control mice (treatment:F(1,14)= 0.0001,P> 0.9999; social squares:F(2,28)= 9.4150,P< 0.0010; interactions:F(2,28)= 3.0970,P= 0.0609), although the AAV-hM3-treated mice showed a prominently higher social interaction index than the control mice (t(14)= 2.7930,P= 0.0144).Additionally, the AAV-hM3-treated mice spent more time sniffing a new stranger-2 mouse (P< 0.0010),but not a familiar stranger-1 mouse (P= 0.4245) (treatment:F(1,14)= 11.9500,P= 0.0039; social squares:F(1,14)= 74.7000,P< 0.0010; interactions:F(1,14)=35.9800,P< 0.0010) compared with the control mice, and had a significantly higher discrimination index than the control mice (t(14)= 6.1210,P< 0.0010;Figure 3C).Finally, we tested anxiety and locomotor activity.As with the open field test, the AAV-hM3-treated mice exhibited no alterations in the amount of time spent in the arena center (t(14)= 0.0734,P= 0.9426; Additional Figure 2C), or the distance travelled in the locomotor test (treatment:F(1,14)= 2.9700,P= 0.1068; timepoints:F(14,196)= 16.9100,P< 0.0010; interactions:F(14,196)=0.7406,P= 0.7317; total distance:t(14)= 1.7230,P= 0.1068; Additional Figure 2D).Thus, LS GABAergic neuron activation appears to enhance preference as well as social novelty behaviors in mice.Furthermore, we measured the efficiency of hM3, which mediates neural stimulating activity, and found that the number of c-Fos-positive cells in the AAV-hM3 mice LS was significantly higher than that in the control group (unpairedt-test with Welch’s correction,P= 0.0410; Figure 3D).Moreover, in comparison with the control group, the action potential frequency (t(20)= 6.3800,P< 0.0010) was augmented and the action potential amplitude remained unaltered (unpairedt-test with Welch’s correction,P= 0.5952) in the AAV-hM3 group (Figure 3E).
Re-inhibiting socially induced LS GABA neuronal activation suppresses social behaviors
Next, we examined the effect of specifically modulating social experienceactivated LS GABA neurons on social behaviors.Fos-CreERT2transgenic mice were used for this purpose.The CreERT2fusion protein comprises Cre recombinase, which is fused to the human estrogen receptor with a triple mutant form.This protein is restricted to the cytoplasm, because it does not attach to 17β-estradiol (its natural ligand) at physiological concentrations and enters the nuclear compartment after binding to tamoxifen, a synthetic estrogen receptor ligand (Guenthner et al., 2013).In our experimental design,we bilaterally administered an AAV that expressed Cre-dependent hM4D(Gi)(designated as AAV-hM4) as well as control mCherry virus (designated as AAV-mCherry) into Fos-CreERT2mice LS.At 3 weeks after treatment, these mice were subjected to the chronic social experience paradigm.Tamoxifen was then intraperitoneally injected to induce Cre activity by promoting its transport to the nucleus, and we conducted the three-chamber test 45 minutes following CNO administration (Figure 4A).We found that the Cre-dependent hM4D(Gi) virus was expressed exclusively in the LS, which indicates that the social experience-activated LS GABA neurons were successfully labeled (Figure 4A).
In the social preference phase, compared with the control mice (AAVmCherry), the AAV-hM4-injected mice spent much less time exploring the stranger-1 chamber (P= 0.0030), and roughly the same amount of time exploring the middle chamber (P= 0.9941) and object chamber (P= 0.0976)(treatment:F(1,13)= 2.0220,P= 0.1786; social squares:F(2,26)= 47.7200,P<0.0010; interactions:F(2,26)= 6.1310,P= 0.0066), and the AAV-hM4-injected mice showed a markedly decreased social interaction index (t(13)= 2.4680,P= 0.0282).Furthermore, compared with the control mice, AAV-hM4-injected mice spent less time sniffing the stranger-1 (P= 0.0078), but not the object(P= 0.9114) (treatment:F(1,13)= 3.2920,P= 0.0927; social squares:F(1,13)=35.2600,P< 0.0010; interactions:F(1,13)= 7.6590,P= 0.0160).Moreover, the AAV-hM4-injected mice did not show obvious differences in the discrimination index when compared with the control mice (t(13)= 1.5590,P= 0.1429; Figure 4B).Thus, social preference appears to have been impaired following the reinhibition of social experience-activated LS GABAergic neurons.
Next, during the social novelty phase, we observed similar changes in thetime spent by the AAV-hM4-injected mice in exploring the stranger-2 chamber(P= 0.8521), middle chamber (P= 0.9984), and familiar mouse chamber(P= 0.7697) (treatment:F(1,13)= 0.0001;P> 0.9999; social squares:F(2,26)= 29.5700,P< 0.0010; interactions:F(2,26)= 0.4388,P= 0.6495).The social interaction index of the AAV-hM4-injected mice also remained unchanged in comparison with that of the control group (t(13)= 1.0040,P= 0.3335; Figure 4C).Furthermore, we found no differences in the time spent by the AAV-hM4-injected mice in sniffing the new stranger-2 mouse (P= 0.1862) or the familiar stranger-1 mouse (P= 0.4917) (treatment:F(1,13)= 0.1295,P= 0.7248; social squares:F(1,13)= 29.9700,P< 0.0010; interactions:F(1,13)= 8.3920,P= 0.0125)when compared with the control mice.However, the AAV-hM4-injected mice had a significantly lower discrimination index compared with the control mice(unpairedt-test with Welch’s correction,P< 0.0010; Figure 4C).
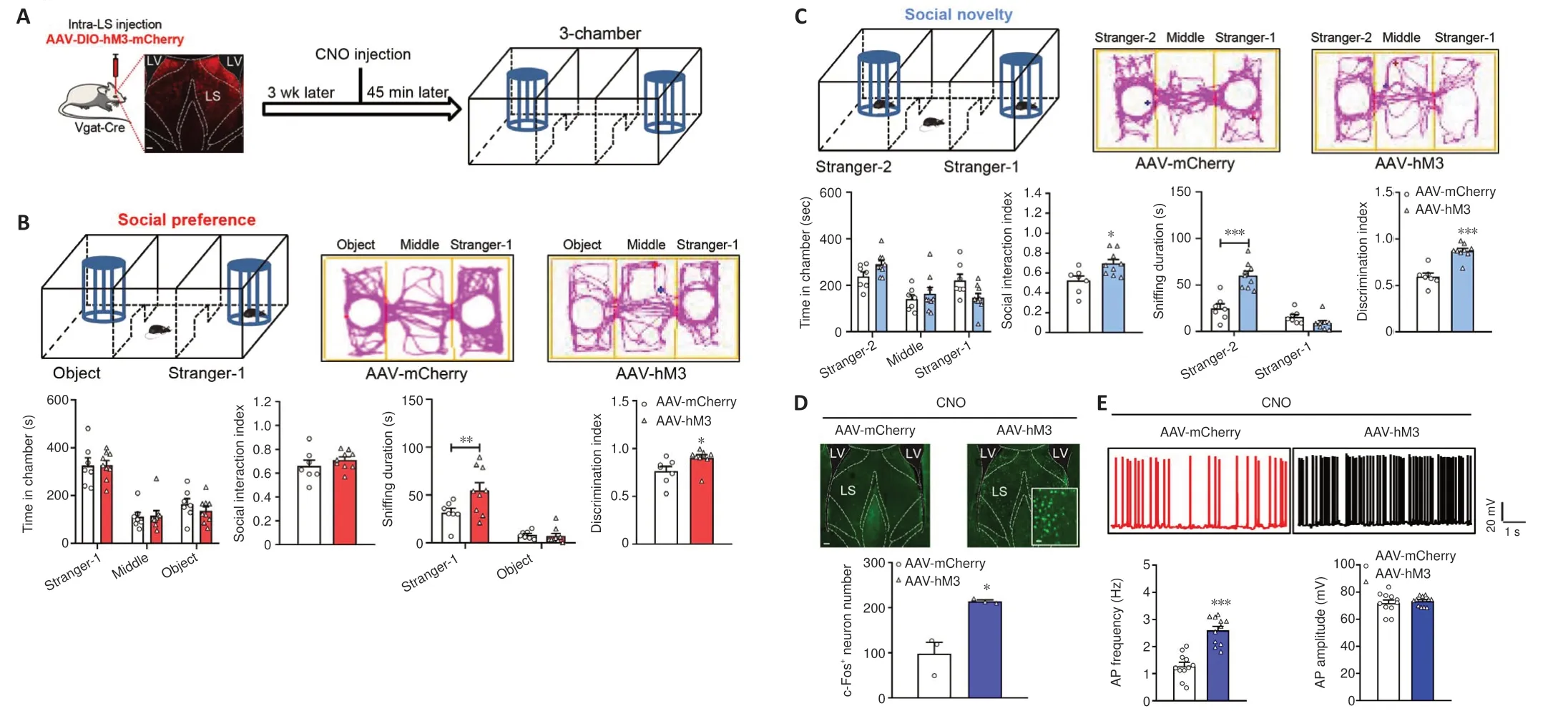
Figure 3 | The activation of LS GABA neurons increases social behaviors.
In the open field test, we found no group differences in the time spent in arena’s center (t(13)= 1.2120,P= 0.2472; Additional Figure 3A) or the distance traveled in the locomotor test (treatment:F(1,13)= 1.7690,P= 0.2064;timepoints:F(14,182)= 22.8900,P< 0.0010; interactions:F(14,182)= 1.2540,P=0.2405; total distance: two-tailed unpairedt-test with Welch′s correction,P= 0.2408; Additional Figure 3B).We also measured the efficiency of hM4 in mediating neural inhibitory activity.We used a 30-minute restraint paradigm to increase basal neural activity, and measured the number of c-Fos-positive cells 45 minutes after the CNO injection.AAV-hM4 mice showed a dramatic reduction in c-Fos-positive cells in the LS compared with control mice (t(4)=4.7540,P= 0.0089; Figure 4D).
Re-activating socially induced LS GABA neuronal activation facilitates social behaviors
We further assessed the effect of specifically re-activating the social experience-activated LS GABA neurons on social behaviors.According to the re-inhibition procedure, we injected an AAV expressing Cre-dependent hM3D(Gq) (designated as AAV-hM3) as well as mCherry virus as a control(designated as AAV-mCherry) into the Fos-CreERT2mice LS.After 3 weeks, we conducted a test using a chronic social experience paradigm.Cre activity was induced by intraperitoneally injecting tamoxifen.We conducted the threechamber test 45 minutes following CNO administration (Figure 5A).The Cre-dependent hM3D(Gq) virus was expressed in the LS, indicating that LS GABA neuron activation was modulated by social experience (Figure 5A).AAV-hM3-treated mice displayed no alterations in the amount of time spent in the stranger-1 chamber (P= 0.2346), middle chamber (P= 0.9726), or object chamber (P= 0.1078) (treatment:F(1,13)= 0.0001,P> 0.9999; social squares:F(2,26)= 132.9000,P< 0.0010; interactions:F(2,26)= 2.6390,P= 0.0905;Figure 5B) or the social interaction index compared with the control mice(AAV-mCherry) (t(13)= 1.3900,P= 0.1877) in the social preference phase.Furthermore, compared with the control mice, AAV-hM3-injected mice spent more time sniffing the stranger-1 (P< 0.0010), but not the object (P= 0.9257)(treatment:F(1,13)= 14.0100,P= 0.0025; social squares:F(1,13)= 220.7000,P<0.0010; interactions:F(1,13)= 24.0700,P< 0.0010).The AAV-hM3-treated mice had a significantly elevated discrimination index compared with the control mice (t(13)= 4.2730,P< 0.0010; Figure 5B).
Compared with the control mice, the test mice showed similar alterations in the amount of time taken to explore the stranger-2 chamber (P= 0.9225),middle chamber (P= 0.7853), and familiar mouse chamber (P= 0.9893)(treatment:F(1,13)= 0.0001,P> 0.9999; social squares:F(2,26)= 35.2200,P<0.0010; interactions:F(2,26)= 0.3739,P= 0.6917).Furthermore, the social interaction index did not significantly differ between the AAV-hM3-injected group and the control group (Mann-Whitney test,P= 0.9551) in the social novelty phase (Figure 5C).However, the AAV-hM3-injected mice spent more time sniffing the new stranger-2 mouse (P< 0.0010) but not the familiar stranger-1 mouse (P= 0.8014) (treatment:F(1,13)= 8.5130,P= 0.0120; social squares:F(1,13)= 70.0700,P< 0.0010; interactions:F(1,13)= 8.4050,P= 0.0124),and showed an increased discrimination index (P= 0.0059; Mann-WhitneyUtest) compared with the control mice (Figure 5C).Thus, the reactivated LS GABAergic neurons appear to have promoted social novelty.
The open field test data revealed no difference in the amount of time spent in the arena’s center (P= 0.2810; Mann-WhitneyUtest; Additional Figure 3C) and the distance traveled in the locomotor test (treatment:F(1,13)=0.1184,P= 0.7363; timepoints:F(14,182)= 38.3100,P< 0.0010; interactions:F(14,182)= 1.3760,P= 0.1685; total distance:P= 0.6126; Mann-WhitneyUtest;Additional Figure 3D) between the two groups, suggesting the absence of anxious behaviors.The hM3-mediated neural activity was efficient, because the c-Fos-positive cell number was substantially higher in the AAV-hM3 mice LS than in the control mice at 45 minutes following CNO administration (t(4)=4.0250,P= 0.0158; Figure 5D).
The inhibition of LS-vCA1 neural projection represses social behaviors
We previously reported on the presence of neural projections from the LS to the periaqueductal gray (Zhang et al., 2022).Herein, we investigated another brain region, vCA1 that is closely associated with social behaviors.The Vgat-Cre mice LS was infused with the AAV-DIO-mCherry virus.After 3 weeks,mCherry+cell bodies were found in the LS; furthermore, many mCherry+fibers were detected in the vCA1 region, but not in the dorsal CA1 region(Additional Figure 4A).To confirm LS-vCA1 projections, we injected the retrograde AAV-Retro-GFP virus into the vCA1 of wild-type mice (Additional Figure 4B).After 4 weeks of treatment, neurons showing GFP expression were found in the LS (Additional Figure 4B).We also used a virus tracing approach to demonstrate active synaptic connections between LS neuronal terminals and vCA1 neurons.The AAV2/1-Vgat-Cre virus enables the Cre enzyme to transfer monosynaptically as well as anterogradely into the soma of LS neurons (Zingg et al., 2017).We injected this virus into the LS, and then injected the Cre-dependent AAV-DIO-GFP virus into the vCA1.At 4 weeks after the injection, GFP-labeled neurons were localized in the vCA1 region(Additional Figure 4C).
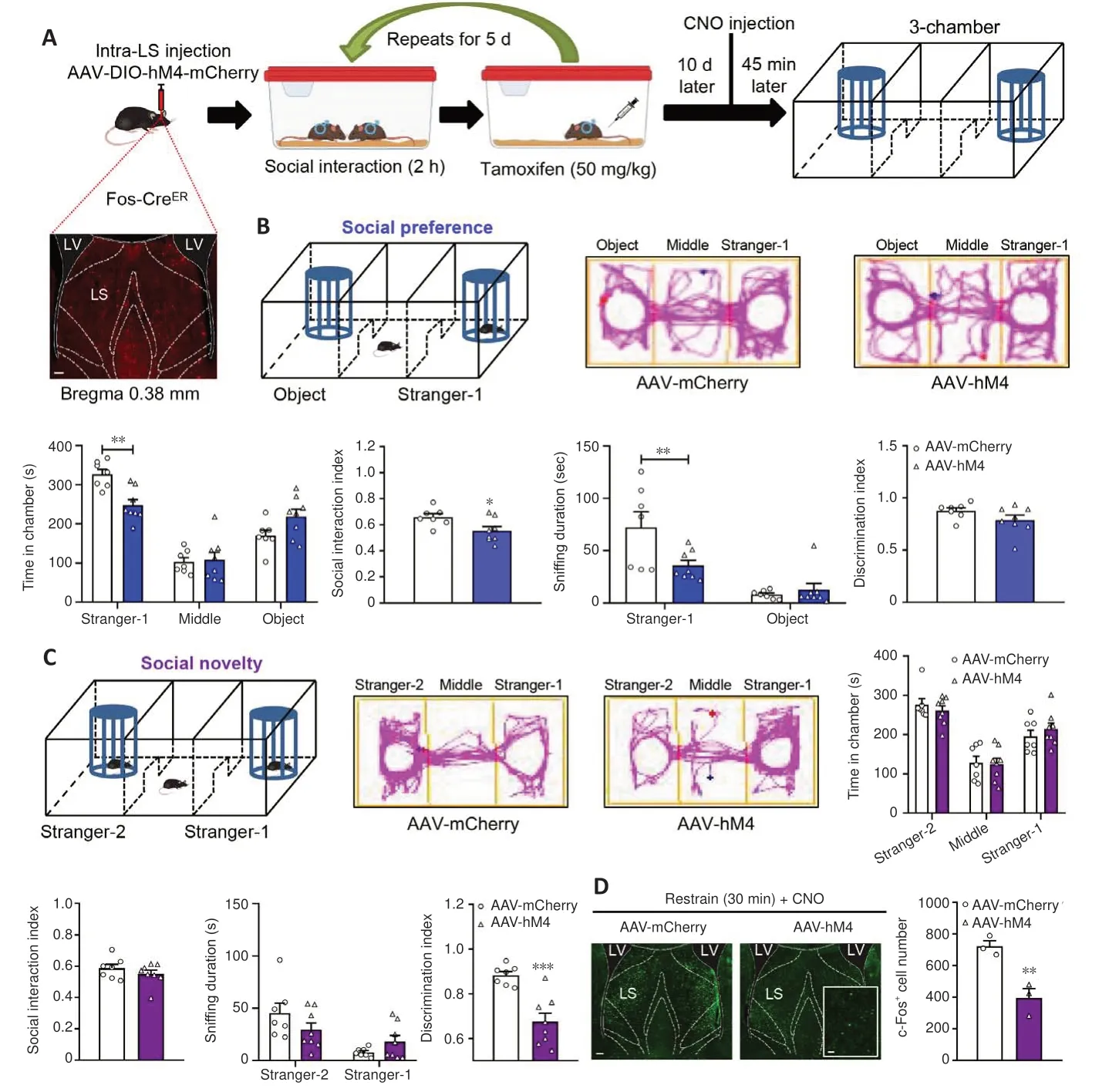
Figure 4 |Re-inhibition of socially activated LS GABA neurons attenuates social behaviors.
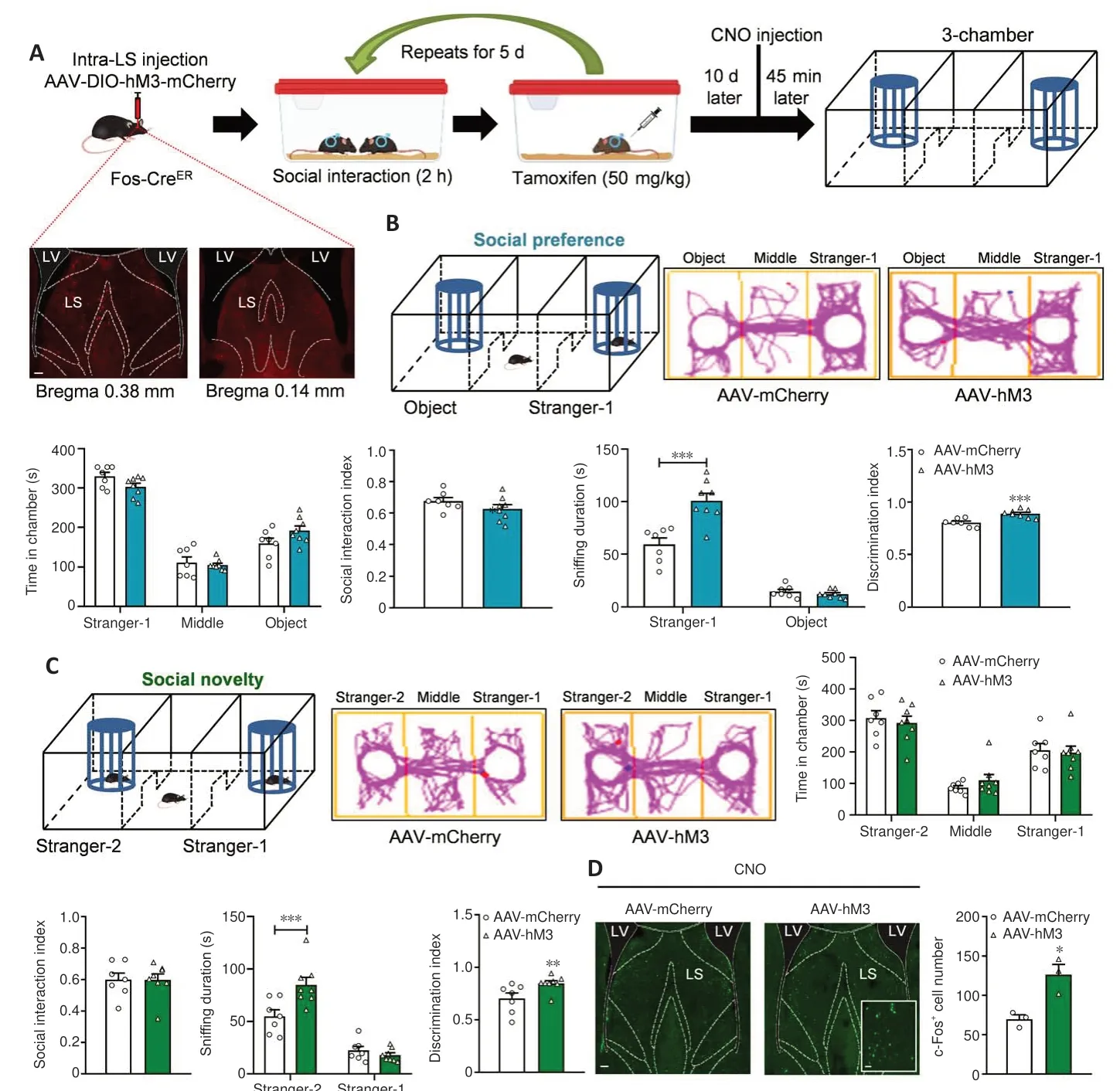
Figure 5 |Social behaviors are enhanced by reactivating socially activated LS GABA neurons.
We next assessed the role of LS-vCA1 projections in modulating social behavior.First, we managed the activity of vCA1-projecting LS neurons by injecting the AAV-Retro-Cre-GFP virus into the vCA1 and an AAV expressing Cre-dependent hM4D(Gi) (designated as AAV-hM4) into the LS.After 3 weeks,we measured social behaviors of these mice (Additional Figure 5A).The AAVhM4-treated mice did not show any alterations in the amount of time spent in the stranger-1 chamber (P= 0.9117), middle chamber (P= 0.9045), or object chamber (P= 0.5496) (treatment:F(1,14)= 0.0001,P> 0.9999; social squares:F(2,28)= 39.8400,P< 0.0010; interactions:F(2,28)= 0.7307,P= 0.4905), or in the social interaction index when compared with the control mice (t(14)= 1.0470,P= 0.3130) in the social preference phase (Additional Figure 5B).Nevertheless,compared with the control mice, the AAV-hM4 mice spent less time sniffing the stranger-1 chamber (P< 0.0010) but not the object chamber (P= 0.0828)(treatment:F(1,14)= 24.1700,P< 0.0010; social squares:F(1,14)= 106.4000,P< 0.0010; interactions:F(1,14)= 75.6600,P< 0.0010).The discrimination index of the AAV-hM4 mice was significantly reduced compared with that of the control mice (t(14)= 7.6250,P< 0.0010; Additional Figure 5B).
In the social novelty phase, the AAV-hM4 mice spent less time in the stranger-2 chamber (P= 0.0038), the same amount of time in the middle chamber (P= 0.9979), and more time in the familiar mice chamber (P= 0.0024)(treatment:F(1,14)= 0.0001,P> 0.9999; social squares:F(2,28)= 31.0000,P< 0.0010; interactions:F(2,28)= 8.3440,P= 0.0014) when compared with the control mice.Furthermore, the AAV-hM4 mice had a reduced social interaction index (t(14)= 3.3180,P= 0.0051) (Additional Figure 5C).The AAVhM4 mice spent less time than the control mice sniffing the new stranger-2(P< 0.0010), but not the familiar stranger-1 mouse (P= 0.1049) (treatment:F(1,14)= 11.4300,P= 0.0045; social squares:F(1,14)= 35.2600,P< 0.0010;interactions:F(1,14)= 69.9700,P< 0.0010) and had a lower discrimination index (t(14)= 8.3240,P< 0.0010; Additional Figure 5C).The open field test data revealed no changes in the time spent in the arena’s center (t(14)= 0.7040,P= 0.4930; Additional Figure 5D) or in the distance covered in the locomotor test (treatment:F(1,14)= 2.5630,P= 0.1317; timepoints:F(14,196)= 33.2800,P<0.0010; interactions:F(14,196)= 1.2800,P= 0.2223; total distance:t(14)= 1.6010,P= 0.1317; Additional Figure 5E), suggesting anxiety-negative behaviors.Thus, the inhibition of vCA1-projecting LS GABAergic neurons appeared to attenuate social preference and novelty behaviors.
Next, we investigated whether the direct suppression of neural fibers in vCA1 deriving from the LS affects social behaviors.We injected an AAV expressing Cre-dependent hM4D(Gi) (designated as AAV-hM4) into the Vgat-Cre mice LS.After 3 weeks, we implanted cannulas in the vCA1, and CNO microinfusion was applied 1 week later, the three-chamber test was conducted 45 minutes after CNO infusion (Figure 6A) The social preference phase data showed that the time spent in the three chambers (Stranger-1:P= 0.9664; middle:P= 0.2480; object:P= 0.1088) (treatment:F(1,12)= 0.0001,P> 0.9999; social squares:F(2,24)= 104.2000,P< 0.0010; interactions:F(2,24)= 2.6190,P= 0.0936)and the social interaction index (t(12)= 1.6420,P= 0.1265) of the AAV-hM4-administered mice were almost identical to those of the control mice (Figure 6B).However, the AAV-hM4 mice spent less time sniffing the stranger-1 mouse (P< 0.0010), but not the object (P= 0.5907) (treatment:F(1,12)=20.5800,P< 0.0010; social squares:F(1,12)= 98.1800,P< 0.0010; interactions:F(1,12)= 43.7900,P< 0.0010), and had a lower discrimination index (t(12)=4.6100,P< 0.0010) than the control mice (Figure 6B).
Furthermore, the AAV-hM4 mice spent less time at the stranger-2 chamber(P= 0.0322), the same amount of time in the middle chamber (P= 0.0528),and a large amount of time in the familiar mice chamber (P< 0.0010)(treatment:F(1,12)= 0.7347,P= 0.4082; social squares:F(2,24)= 37.5700,P< 0.0010; interactions:F(2,24)= 11.9100,P< 0.0010), although their social interaction index was reduced (t(12)= 6.0980,P< 0.0010).Furthermore, in comparison with the control mice, AAV-hM4 mice spent less time sniffing the new stranger-2 (P< 0.0010), but the same amount of time sniffing familiar stranger-1 mouseP= 0.0932) (treatment:F(1,12)= 0.1527,P= 0.6987;social squares:F(1,12)= 7.6620,P= 0.0170; interactions:F(1,12)= 37.2300,P<0.0010), and had a reduced discrimination index (t(12)= 11.5300,P< 0.0010;Figure 6C).
The assessment of anxiety and locomotor activity indicated that in the open field test, there were no significant differences of the time spent in the arena’s center (t(12)= 0.1897,P= 0.8527; Additional Figure 6A) or the distance covered in the locomotor test (treatment:F(1,12)= 0.0192,P= 0.8921;timepoints:F(14,168)= 10.8800,P< 0.0010; interactions:F(14,168)= 1.2190,P= 0.2656; total distance: Mann Whitney test,P= 0.5728; Additional Figure 6B).These results indicate that social novelty behaviors were repressed by inhibiting the neural fibers in vCA1 derived from LS GABAergic neurons.
Identification of targeted neurons and their social function in vCA1 neurons receiving synaptic connections from the LS
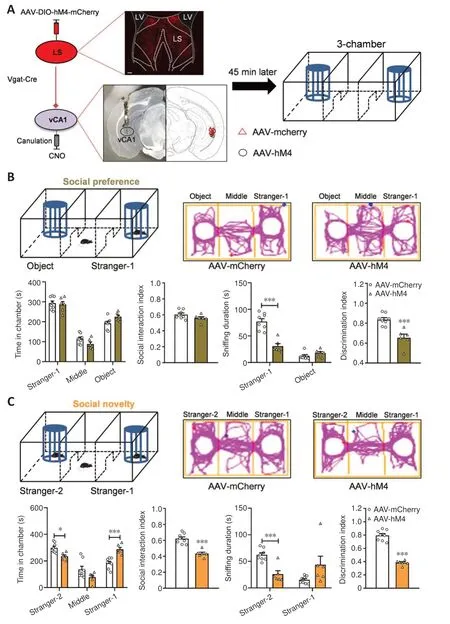
Figure 6 |Regulation of social behaviors by inhibiting neuronal fibers in vCA1 originating from GABAergic neurons in the LS.
We confirmed that LS GABA neuron activation enhances social behaviors and confirmed the effect of LS-vCA1 projection on social behavior.We then analyzed the neural types of LS synaptic targeted neurons in vCA1 using the monosynaptic and anterograde AAV2/1-Vgat-Cre virus and immunofluorescence co-localization analysis (Figure 7A).We injected a mixture of AAV2/1-Vgat-Cre and AAV-DIO-mCherry into the LS, and AAV-DIOGFP into vCA1, and 3 weeks later found that the mCherry labeled neurons in the LS were co-labeled with GABA (97.52 ± 1.21%).This supports the specificity of AAV2/1-Vgat-Cre in GABAergic neurons (Additional Figure 7A and B).LS synaptic targeted neurons in the vCA1 were co-localized with both glutamatergic and GABAergic neurons (80.93 ± 1.25% and 19.07 ± 1.24%,respectively).We further determined whether the LS GABAergic neurons regulated the activity of these two types of targeting neurons.The patch clamp test results showed increased neural activity of glutamatergic targeted neurons following the inhibition of LS GABAergic neurons, with more action potentials elicited by the same amount of current injection (treatment:F(1,27)= 25.8900,P< 0.0010; current:F(12,324)= 206.3000,P< 0.0010; interactions:F(12,324)= 11.3700,P< 0.0010), lower rheobase current (t(27)= 6.3290,P< 0.0010), higher input resistance (t(27)= 3.7670,P< 0.0010), and lower hyperpolarization amplitude (t(27)=2.3550,P= 0.0260) and action potential threshold (P= 0.0019; Mann-WhitneyUtest) than the control group (Figure 7B).Other membrane properties, including the action potential half width(t(27)= 1.1720,P= 0.2514) and amplitude of action potentials (P= 0.3942;Mann-WhitneyUtest) were not significantly different between these groups(Figure 7B).Furthermore, both groups showed no changes in action potential frequency (t(12)= 0.0239,P= 0.9813) or action potential amplitude (t(12)=1.1840,P= 0.2593) for GABAergic neurons (Figure 7B).
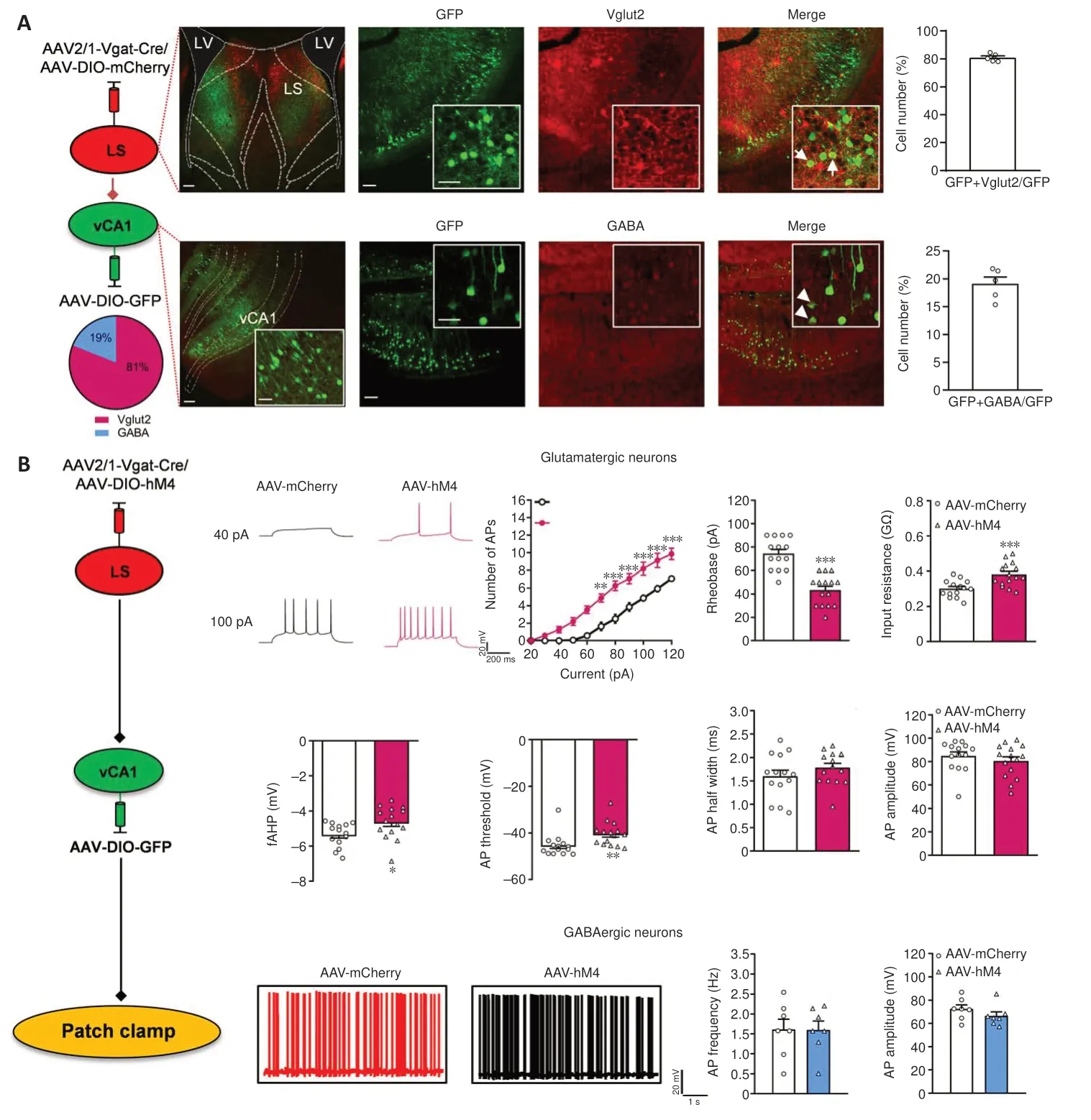
Figure 7 |Determining the effect of LS GABAergic neurons on vCA1-targeted projection neurons.
Next, to further confirm the specific modulatory role of LS synaptic targeted glutamatergic neurons in the vCA1 in social behaviors, we infused the anterograde and monosynaptic AAV2/1-Vgat-Cre virus into the LS and the CaMKII promoter-derived Cre-dependent hM4D(Gi) virus (designated as AAV-hM4) into the vCA1 (Figure 8A), which is specifically expressed in glutamatergic neurons (Zhao et al., 2021).We measured social behaviors 45 minutes following CNO administration.In line with our expectations, the AAV-hM4 mice spent much longer in the stranger-1 chamber (P= 0.0035),the same amount of time in the middle chamber (P= 0.9994), and less time in the object chamber (P= 0.0026), and had an elevated social interaction index (t(14)= 3.4530,P= 0.0039) (treatment:F(1,14)= 0.0001,P> 0.9999; social squares:F(2,28)= 25.2200,P< 0.0010; interactions:F(2,28)= 8.3430,P= 0.0014)in the social preference phase.Moreover, compared with the control mice,the test mice spent a similar amount of time sniffing stranger-1 mice and object (P= 0.7925 andP= 0.7300) (treatment:F(1,14)= 0.0046,P= 0.9468;social squares:F(1,14)= 88.2000, P < 0.0010; interactions:F(1,14)= 1.0030,P=0.3336) and had an increased discrimination index (t(14)= 2.4790,P= 0.0265;Figure 8B).Furthermore, in the social novelty phase, AAV-hM4 mice spent a comparable amount of time in the three chambers (Stranger-2:P= 0.7635,middle:P= 0.9998, and Stranger-1: P = 0.7203) (treatment:F(1,14)= 0.0001,P> 0.9999; social squares:F(2,28)= 21.5900,P< 0.0010; interactions:F(2,28)=0.5650,P= 0.5747) and a comparable amount of time sniffing the stranger-2 and stranger-1 mice (P= 0.2891 andP= 0.9937); (treatment:F(1,14)= 0.7597,P= 0.3981; social squares:F(1,14)= 29.9400,P< 0.0010; interactions:F(1,14)=1.5260,P= 0.2371; Figure 8C).The social interaction index (t(14)= 1.0070,P= 0.3310) and discrimination index (t(14)= 1.0570,P= 0.3083) were similar between the groups.These results showed that social preference behaviors were accelerated by inhibiting LS synaptic targeted glutamatergic neurons.
Inhibiting the activity of LS-projected glutamatergic neurons in vCA1 blocked the effect on social behaviors induced by LS GABAergic neuron suppression
After evaluating the role of LS synaptic targeted glutamatergic neurons in vCA1 in social preference behaviors, we next determined whether these neurons mediated the effect exerted by LS GABAergic neurons in terms of modulating social behaviors.We co-injected the monosynaptic and anterograde AAV2/1-Vgat-Cre and Cre-dependent hM4D(Gi) virus into the LS (designated as AAVhM4 (LS)) and the CaMKII promoter-derived Cre-dependent hM4D(Gi) virus into vCA1 (designated as AAV-hM4 (vCA1)) (Figure 9A).We tested social behaviors 45 minutes following CNO administration.In the social preference phase, the AAV-hM4 (LS)-treated mice spent a less amount of time in stranger-1 chamber and spent less time sniffing (P< 0.0010 andP= 0.0110,respectively).Furthermore, such effects were restored in the AAV-hM4(vCA1)-treated mice (P= 0.0049 andP= 0.0054, respectively).No changes were noted in the amount of time spent in the other chambers (middle:P= 0.8132 andP= 0.8670, respectively; object:P= 0.1190 andP= 0.4408,respectively) (treatment:F(2,20)= 5.3360;P= 0.0139; social squares:F(2,40)=55.8500,P< 0.0010; interactions:F(4,40)= 3.6120,P= 0.0132) or the sniffing duration for the object (P= 0.8402 andP= 0.9655, respectively) (treatment:F(2,20)= 1.9680,P= 0.1659; social squares:F(1,20)= 154.5000,P< 0.0010;interactions:F(2,20)= 6.2930,P= 0.0076).The reduced social interaction index(P= 0.0383) and discrimination index (P= 0.0015) in the AAV-hM4 (LS)-treated mice were also recovered by AAV-hM4 (vCA1) treatment (P= 0.0482,F(2,20)=4.8320,P= 0.0194; andP= 0.0068,F(2,20)= 10.0900,P< 0.0010; Figure 9B).
In the social novelty phase, the AAV-hM4 (LS)-treated mice spent less time in the new stranger-2 chamber as well as engaged in sniffing stranger-2 (P<0.0010 andP= 0.0117, respectively), whereas AAV-hM4 (vCA1) treatment recovered this decline (P< 0.0010 andP= 0.0289, respectively).Furthermore,the time spent in the other chambers did not differ significantly, except for the decreased time spent in the stranger-1 chamber after AAV-hM4 (vCA1)treatment (middle:P= 0.9988 andP= 0.9465; stranger-1:P= 0.6813 andP= 0.0183) (treatment:F(2,20)= 5.0320,P= 0.0170; social squares:F(2,40)=76.5000,P< 0.0010; interactions:F(4,40)= 8.6850,P< 0.0010) and the sniffing duration for the stranger-1 (P= 0.8475 andP= 0.5284) (treatment:F(2,20)=1.2860,P= 0.2982; social squares:F(1,20)= 81.1700,P< 0.0010; interactions:F(2,20)= 5.5240,P= 0.0123; Figure 9C).Finally, the decreased social interaction index (P= 0.0353) for the AAV-hM4 (LS) mice was also recovered by AAVhM4 (vCA1) treatment (P< 0.0010;F(2,20)= 9.8400,P= 0.0011; Figure 9C).No changes were observed in the discrimination index (F(2,20)= 2.7020,P= 0.0915).Meanwhile, the locomotor activity remained unchanged in the treated groups in the locomotor test (treatment:F(2,20)= 0.6998,P= 0.5084; social squares:F(14,280)= 18.9600,P< 0.0010; interactions:F(28,280)= 0.4404,P= 0.9944; total distance:F(2,20)= 0.6998,P= 0.5084; Additional Figure 8).Thus, inhibiting LS synaptic targeted glutamatergic neurons in vCA1 appeared to restore the attenuated social behaviors induced by LS GABAergic neuron suppression.

Figure 8 |The effect of inhibiting LS-targeted vCA1 glutamatergic neurons on social behaviors.
Blocking the GABA receptor in vCA1 abolished the accelerating effect on social behaviors induced by LS GABAergic neuron activation
After confirming that synaptic projections from LS GABAergic neurons to vCA1 pyramidal neurons modulate social behaviors, we assessed how GABA receptors play a role in this process.We used Bicuculline, a GABAA antagonist(Johnston, 2013), to determine whether GABA receptors mediate the effects on social behaviors induced by LS GABAergic neuron activation by bilaterally injecting AAV expressing Cre-dependent hM3D (Gq) (designated as AAVhM3 (LS)) into the LS of Vgat-Cre mice.Then, we implanted cannulas in the vCA1, Bicuculline was infused into the vCA1 for 30 minutes before CNO intraperitoneal injection (Figure 10A).At 45 minutes after the injection,in the social preference phase, we found no significant differences in the amount of time taken to explore the stranger-1 chamber, middle chamber,or object chamber (treatment:F(2,20)= 0.0001,P> 0.9999; social squares:F(2,40)= 56.5100,P< 0.0010; interactions:F(4,40)= 1.5290,P= 0.2122), and no differences in the social interaction index (F(2,20)= 1.5920,P= 0.2282)in the treated mice.Moreover, AAV-hM3 (LS)-administered mice spent a considerable amount of time sniffing the stranger-1 (P< 0.0010), and bicuculline treatment attenuated this behavior (P< 0.0010), although thetime spent sniffing the object remained unchanged (treatment:F(2,20)= 7.5900,P= 0.0035; social squares:F(1,20)= 85.0400,P< 0.0010; interactions:F(2,20)=20.8000,P< 0.0010).The elevated discrimination index was also significantly reversed by bicuculline treatment (P= 0.0269 andP< 0.0010) (F(2,20)=16.0600,P< 0.0010; Figure 10B).
In the social novelty phase, the time spent exploring the stranger-2 chamber,middle chamber, and familiar stranger-1 chamber was similar (treatment:F(2,20)= 0.0001,P> 0.9999; social squares:F(2,40)= 51.2800,P< 0.0010;interactions:F(4,40)= 0.9840,P= 0.4272); moreover, these treated mice had a similar social interaction index (treatment:F(2,20)= 0.7930,P= 0.4662).Bicuculline treatment also reduced (P= 0.0118) the enlarged amount of time spent sniffing the stranger-2 chamber in the AAV-hM3 (LS)-injected mice (P<0.0010), but not the familiar stranger-1 chamber (treatment:F(2,20)= 2.7430,P= 0.0886; social squares:F(1,20)= 25.0700,P< 0.0010; interactions:F(2,20)= 15.7300,P< 0.0010).Furthermore, bicuculline treatment (P= 0.0180)reversed the heightened discrimination index in these mice (P= 0.0057) (P= 0.0035; Kruskal-Wallis test; Figure 10C).We found no differences in the total distances travelled by the different groups (F(2,20)= 0.7877,P= 0.4685;Figure 10D).The microinjection sites were exclusively located in the desired vCA1 area (Figure 10E).Thus, blocking the GABAAreceptor appears to have abolished the promoted social behavior through LS GABAergic neuron activation.
Discussion
Here, we examined whether social interaction triggered the accelerated neural excitability of LS GABAergic neurons and whether the activation or re-activation of these triggered neurons increased social behaviors.We also assessed whether the inhibition or re-inhibition of these related neurons reduced social behaviors.The LS GABAergic neurons extended robust neuronal fibers to the vCA1 and regulated the activity of pyramidal neurons, and inhibition of this LSGABA→ vCA1Gluneural circuit abolished social behaviors.Finally, the enhanced social behaviors induced by LS GABAergic neuronal activation were attenuated via inhibition of these projected vCA1Gluneurons or a GABAAreceptor block in vCA1.Overall, our data provide direct evidence for the functional effect of the LSGABA→vCA1Gluneural circuit on social behaviors in mice.
Incentive behaviors, including eating, drinking, touching, sex, and social interaction, are important for survival and reproduction, and these behaviors are reinforced by so-called “natural rewards” (Kelley and Berridge, 2002).Our current data indicate that both acute and chronic social activity activate LS GABA neurons.We found some activated GABA neurons in the LS in the control group, likely as a result of nonspecific reactivity, indicating that not all sensitized GABA neurons were associated with the tested social behaviors.However, our findings regarding the re-activation of socially induced LS GABA neurons in sociability validated the specific effect of these activated GABA neurons.A recent study suggested that LS activation modifies behavior in some social contexts by enhancing sex-naïve social approach behaviors as well as prosocial behavior in pair-bonded males (Sailer et al., 2022).We further showed that chemogenetic activation of LS GABAergic neurons promoted not only social preference (objectvs.stranger-1) but also social novelty behavior (stranger-2vs.stranger-1).However, a previous study found that LS was activated with abundant c-Fos-positive cells after the suppression of natural reward-seeking behaviors by adverse stress factors, and chemogenetic inhibition of the broad neurons in the intermediolateral septum restored the reward-seeking behavior (Zhang et al., 2022).This finding seems to be contradictory to the regulation of natural rewards by LS neurons.Furthermore, several studies have demonstrated that specific marker neurons in the LS, such as neurotensin, type 2 corticotropin-releasing hormone receptor and vasopressin 1A receptor neurons, also modulate sociabilityrelated behaviors, using different regulatory patterns (Bielsky et al., 2005;Hashimoto et al., 2022; Li et al., 2023).This complex phenomenon might be because of the following factors: complexity and heterogeneity of neuronal subtypes, subregions in the LS, and individual modulatory strategies.The LS not only contains dense populations of GABAergic inhibitory neurons, but also includes cholinergic and glutamatergic neurons (Kiss et al., 1990; Zhao et al.,2013; Wang et al., 2021).Based on the functional connections and distinct anatomical locations, the LS is classified into three subdivisions: the ventral,intermediate, and dorsal regions.Each region receives complicated extraand intra-septal inputs and projects to many brain regions (Sheehan et al.,2004).Our previous study showed that vesicular GABA transporter-positive GABAergic neurons in the LS have a higher level of c-Fos activation following chronic unpredictable stress, and that the activation of LS GABA neurons and their projections to the periaqueductal gray increased depressivelike behaviors.In contrast, inhibition of the LS GABA circuit decreased the phenotypes of depression in mice (Wang et al., 2021).As deficits in social reward are known to be one of the primary symptoms of major depression,it was interesting to find that social experience also activated LS GABA neurons, and that this activation was similar to that of LS GABA neurons by chronic unpredictable stress.This phenomenon could be related to the heterogeneous characteristics of the GABA neurons in the LS.For example,somatostatin-expressing neurons account for approximately 20% of LS GABA neurons, indicating that the LS includes several other subtypes of GABAergic neurons (Li et al., 2022).Therefore, the precise role of the LS in social ability requires further investigation via careful examination of the complicated and/or diverse characteristics of LS neurons.
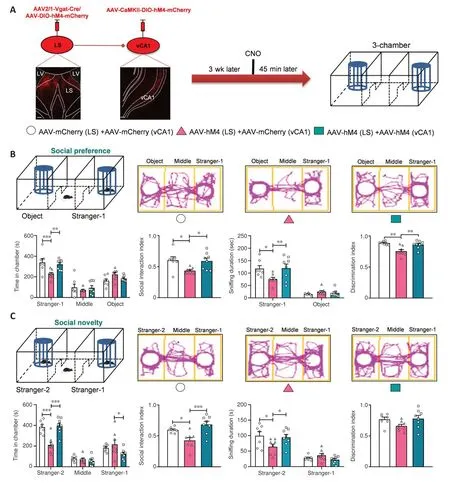
Figure 9 |Inhibition of neurons in the LS targeted vCA1 glutamatergic neurons affected social behaviors regulated by LS GABAergic neurons.
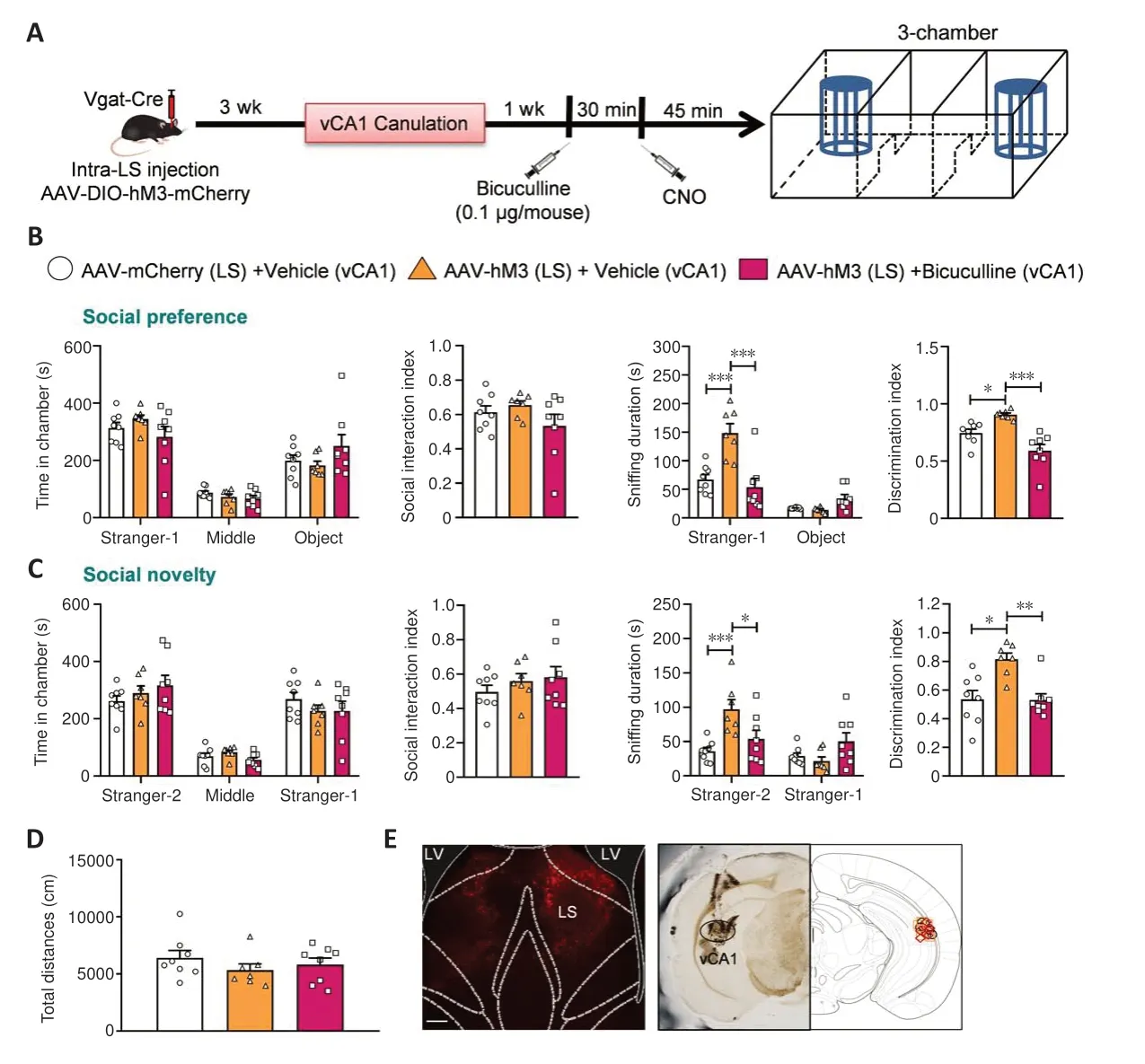
Figure 10 |Social behaviors induced by blocking GABA receptors in vCA1 glutamatergic neurons via the regulation of LS GABAergic neurons.
The LS is an essential nodal point that integrates information, establishes multiple neural connections with many brain areas connected through unidirectional or bidirectional pathways (especially the hippocampus,which is a crucial component for social memory), and promotes social aggression behavior in mice depending on the CA2 output to the LS (Leroy et al., 2018).The present study is novel in that it is the first to indicate that GABAergic neural projections from the LS to vCA1 form neural connections with glutamatergic and GABAergic neurons in the vCA1.Unexpectedly, our electrophysiological recordings showed that the activation of LS GABAergic neurons suppressed only glutamatergic neurons.Further investigation showed that specific attenuation of these LS-projected glutamatergic neurons in the vCA1 promoted social behaviors and reversed the decrease in social behaviors induced by the inactivation of LS GABAergic neurons.According to one report, the ablation of vCA1 neurons that extend inputs to the LS did not affect social novelty recognition (Arszovszki et al., 2014).Thus, vCA1 neurons that receive input from the LS or provide direct input to the LS should be classified differently and may have diverse projecting routes (Arszovszki et al.,2014).
GABA receptors are involved in the regulation of social behaviors (Han et al., 2012; Kratsman et al., 2016; Deng et al., 2019).Microinjection of the GABAAreceptor blocker bicuculline in the parvafox nucleus of chronic social defeat-treated mice blocked an increase in social approach behavior and reduction in avoidance behavior induced by optogenetic activation of the LS GABA→parvafox nucleus pathway (Liu et al., 2022).There are additional lines of evidence to support a possible role of the GABAAreceptor in regulating social behaviors.For example, microinjection of the GABAAreceptor agonist clonazepam into the medial prefrontal cortex rescued impaired social interaction in prenatally valproic acid-exposed mice in the three-chamber test (Yang et al., 2021).Furthermore, muscimol administration restored autism spectrum disorder-like behaviors in akr1c14 deletion mice (Vacher et al., 2021).In the present study, vCA1 was infused with bicuculline after the activation of LS GABA neurons.As expected, the bicuculline injection completely blocked the increase in social behaviors,indicating that the GABAAreceptor signaling pathway in vCA1 mediated the effects on sociability generated by modulation of LS GABAergic neurons.A previous study demonstrated that NMDAR-dependent hippocampal SC–CA1 synaptic plasticity was impaired in Shank2 knockout mice (Won et al.,2012).Furthermore, brain-specific Foxp1 deletion mice displayed autisticlike behaviors, as well as reduced excitability and imbalanced excitatory to inhibitory input in CA1 neurons (Bacon et al., 2015).Increased hippocampal astrogenesis during development damaged social interaction behaviors in mice and enhanced GABAergic transmission in CA1 pyramidal neurons,while the suppression of elevated GABAergic synaptic transmission in the hippocampal CA1 area rescued autistic-like behaviors (Chen et al., 2022).All of these findings support the involvement of CA1 neurons in social or autisticlike behaviors.However, the precise function of the vCA1 GABAAand other GABA receptors in sociability remains unclear, necessitating future research.In conclusion, we report a novel neural circuit, LSGABA→vCA1Glu, and revealed that modulation of this circuit affects social preference/novelty behaviors in the three-chamber paradigm.This finding enriches the roles of the LS in regulating sociability and suggests a new potential neuropathological mechanism underlying social behaviors.However, we did not investigate the functions of different GABAergic neuron subtypes or precise subregions in the LS in terms of social behaviors, and the specific molecular mechanism of the dysfunctional LSGABA→vCA1Gluneural circuit warrants further exploration.Finally, the present study provides new data that could guide the development of potential therapeutic agents for treating neuropsychiatric illnesses involving social impairment.
Author contributions:Conceptualization,formal analysis,writing-original draft,writing-review and editing,supervision,project administration:CL,YZ,WL,DW;investigation and funding acquisition:CL,YZ,WL,DW,DZ,WTW,JL;methodology:CL,YZ,WL,DW,DZ,WTW,JL,MHC,FTM,FAH,CLL,CYQ,DJL,ZCX,YMW;resources,software:DW,DZ,WTW,JL,MHC,FTM,FAH,CLL,CYQ,DJL,ZCX,YMW.All authors read and approved the final version of the manuscript.
Conflicts of interest:The authors declare no conflict of interest.
Data availability statement:All data relevant to the study are included in the article or uploaded as Additional files.
Editor’s evaluation:The manuscript has a very interesting topic(brain circuitry underlying social behavior)and uses a very elegant approach to establish causal relationships(using virus-mediated activation and inhibition of specific neurons).The authors used electrophysiological and chemogenetic approaches to study the role of projections from LS tov CA1 in sociability in the three-chamber test.They found that LS GABAergic neurons extend
functional neuronal connections with vCA1 glutamatergic pyramidal neurons;attenuation of the LSGABA-vCA1Glu neuronal projection impairs social behavior,which was impeded by suppressing the activity of LS-projecting vCA1 neurons.Pharmacological inhibition of the GABAA receptor in vCA1 blocked the enhanced sociability induced by activation of LS GABAergic neurons.They reported a novel neural circuit,LSGABA-vCA1Glu.Overall,the findings are likely to be important,novel,and of broad interest.
Open access statement:This is an open access journal,and articles are distributed under the terms of the Creative Commons
AttributionNonCommercial-ShareAlike 4.0 License,which allows others to remix,tweak,and build upon the work non-commercially,as long as appropriate credit is given and the new creations are licensed under the identical terms.
Additional files:
Additional Figure 1:Chronic social interaction activates LS GABAergic neurons.
Additional Figure 2:Anxiety-related behaviors and locomotor activity after the inhibition or activation of LS GABAergic neurons.
Additional Figure 3:Anxiety-related behaviors and locomotor activity after resuppressing or re-activating social-induced LS GABAergic neurons.
Additional Figure 4:Identification of the neuronal projections from the LS to vCA1.
Additional Figure 5:The effect of inhibiting vCA1-projecting GABAergic neurons in the LS on social behaviors.
Additional Figure 6:Anxiety-related behaviors and locomotor activity after inhibiting the neuronal fibers in vCA1 originating from GABAergic neurons in the LS.
Additional Figure 7:The co-localization analysis of AAV2/1-Vgat-Cre marked neurons with GABAergic neurons.
Additional Figure 8:The locomotor activity of inhibiting the LS targeted vCA1 glutamatergic neurons regulates social behaviors induced by LS GABAergic neurons.

- 中国神经再生研究(英文版)的其它文章
- Glycolysis and glucose metabolism as a target for bioenergetic and neuronal protection in glaucoma
- MAP4K inhibition as a potential therapy for amyotrophic lateral sclerosis
- RNA sequencing of exosomes secreted by fibroblast and Schwann cells elucidates mechanisms underlying peripheral nerve regeneration
- Crosstalk among mitophagy, pyroptosis, ferroptosis,and necroptosis in central nervous system injuries
- Clustering of voltage-gated ion channels as an evolutionary trigger of myelin formation
- Using microglia-derived extracellular vesicles to capture diversity of microglial activation phenotypes following neurological injury