
6株H9N2亚型禽流感病毒分子特征及遗传演化分析
2024-02-03 05:39邢燕茹祝宇翔范春燕于海汪秀会
畜牧与兽医 2024年2期
邢燕茹,祝宇翔,范春燕,于海,汪秀会*
(1. 河北工程大学生命科学与食品工程学院,河北 邯郸 056038;2. 中国农业科学院上海兽医研究所,上海 200241)
禽流感病毒(avian influenza virus,AIV),属于正黏病毒科甲型流感病毒属,是一种人畜共患病原体,禽类是其天然的宿主。流感病毒基因组由8段单链RNA组成,具有很高的原位重组潜力[1]。其中,H9N2亚型AIV属于低致病性禽流感病毒(low pathogenic avian influenza virus,LPAIV),也是目前世界上传播最广泛和最具破坏性的AIV[2]。H9N2亚型AIV与其他亚型AIV在家禽和野生鸟类间广泛传播,加上疫苗广泛接种的压力,促使流感病毒不断进化,为新型变异毒株和重组毒株的产生创造了有利条件[3-4]。H9N2亚型AIV可由家禽直接或间接传播给哺乳动物,包括人类[5-6],对公共卫生安全具有一定的威胁[7]。许多学者认为H9N2亚型AIV是造成人类流感大流行的潜在风险因素[8]。目前H9N2亚型AIV是我国鸡群中流行最为广泛的病毒亚型[2],一旦获得跨种传播的能力,很有可能对人类的生命安全造成威胁和挑战[9]。因此,要加强H9N2亚型AIV的监测和防控,对潜在的流行病学提供早期预警,以降低人类流感大流行的风险[4,10]。
为了解H9N2亚型AIV的分子变化特征及其跨物种传播的可能性,本研究对实验室保存的6株H9N2亚型AIV的8个基因片段进行序列测定和遗传进化分析,并且进一步对6株分离株与6株H9N2亚型AIV代表毒株的血凝素(hemagglutinin,HA)和神经氨酸酶(neuraminidase,NA)同源性和关键氨基酸位点进行了对比分析。
1 材料与方法
1.1 试验材料
在河南、山东、四川等地区采集肺组织病料中分离到6株AIV,分别为A/chicken/Henan/HN1/2011(H9N2),A/chicken/Henan/HN2/2011(H9N2),A/chicken/Shandong/SD-B40/2014(H9N2),A/chicken/Sichuan/SC-B44/2014(H9N2),A/chicken/Shandong/SD-3297/2016(H9N2),A/chicken/Shandong/SD-3220/2016(H9N2),由本实验室保存。
9~11日龄的SPF鸡胚,购自北京勃林格殷格翰维通生物技术有限公司;病毒RNA提取试剂盒,购自Qiagen公司;LATaq预混酶、反转录酶和pMD-18T载体,购自TaKaRa公司;胶回收试剂盒,购自Omega公司;DL2000 DNA Marker,购自全式金公司。

M. DL2000 DNA Marker;1. SD-B40毒株;2. SC-B44毒株;3. HN1毒株;4. HN2毒株;5. SD-3297毒株;6. SD-3220毒株。
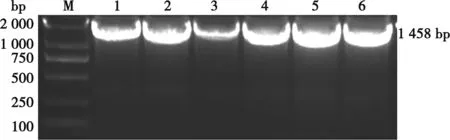
M. DL2000 DNA Marker;1. SD-B40毒株;2. SC-B44毒株;3. HN1毒株;4. HN2毒株;5. SD-3297毒株;6. SD-3220毒株。
1.2 病毒扩增
6株病毒分别接种至5枚9~11日龄SPF鸡胚的尿囊腔中大量增殖,除去24 h内死亡的鸡胚,在37 ℃孵化箱中孵育72 h后将鸡胚放入4 ℃冰箱过夜,用针管吸出鸡胚尿囊液。对鸡胚中吸出的尿囊液进行血凝检测,将血凝阳性的尿囊液保存于-80 ℃冰箱,用于后续病毒基因组的提取,进一步分析毒株的基因序列。
1.3 病毒RNA的提取及反转录
分别提取6株病毒的总RNA,将提取好的病毒RNA反转录为cDNA,放入冰箱保存,温度为-20 ℃。
1.4 8个基因片段PCR扩增及序列测定
分别以6株病毒的cDNA为模板,使用流感病毒通用引物分别对6株病毒的8个基因片段进行PCR扩增。PCR扩增反应体系为:LATaq预混酶25 μL,上游引物2 μL,下游引物2 μL,cDNA 2 μL,ddH2O 19 μL。PCR扩增反应程序为:预变性94 ℃ 3 min;变性94 ℃ 30 s,退火53 ℃ 30 s,延伸72 ℃ 2 min 30 s,35个循环;终延伸72 ℃ 10 min。PCR产物经1%琼脂糖凝胶电泳鉴定,切下与目的基因片段大小一致的片段,胶回收纯化。将含有8个基因片段的PCR产物分别连接到pMD-18T载体上,转化进入大肠杆菌DH5α感受态细胞,送至上海擎科生物科技有限公司进行测序。
1.5 HA及NA同源性分析
利用DNAStar中的SeqMan软件对测序得到的各基因片段序列进行拼接,再利用EditSeq软件对结果进行整理,获得6株病毒完整的HA和NA基因序列,并翻译为氨基酸序列。从NCBI数据库中下载各谱系代表毒株的核苷酸和氨基酸序列,6株代表毒株分别为A/Duck/Hong Kong/Y280/97(H9N2),A/Duck/Hong Kong/Y439/97(H9N2),A/quail/Hong Kong/G1/1997(H9N2),A/Chicken/Hong Kong/G9/97(H9N2),A/Chicken/Beijing/1/94(H9N2),A/Chicken/ShangHAi/F/98(H9N2)。利用MegAlign 7.1软件分析6株分离株与6株代表毒株HA和NA序列的同源性。
1.6 8个基因片段遗传进化分析及系统进化树的构建
从NCBI上下载19株代表毒株的8个基因片段序列,同时将6株病毒的8个基因片段序列提交到NCBI数据库上的BLAST模块进行比对,将同源性最高的序列作为参考序列,将分离到的6株病毒与19株代表毒株的8个基因片段序列利用MEGA 7.0软件分别进行遗传进化分析,并完成系统进化树的构建。
1.7 HA和NA蛋白关键氨基酸位点分析
从NCBI数据库下载H9N2亚型AIV的6株代表毒株(同1.5)HA和NA蛋白的氨基酸序列,将分离到的6株病毒与6株代表毒株的HA和NA蛋白氨基酸序列一起导入MegAlign 7.1软件,比较分析关键氨基酸位点。
2 结果与分析
2.1 病毒扩增
将实验室保存的6株病毒接种于SPF鸡胚,收取尿囊液后通过血凝检测鉴定病毒的分离情况,收取的6株病毒鸡胚尿囊液均可使1%鸡红细胞凝集,说明6株病毒均获得扩增。后续对这6株病毒进行病毒RNA提取、RT-PCR扩增及序列测定。
将6株病毒分别接种至SPF鸡胚的尿囊腔中大量增殖,用针管吸出鸡胚尿囊液,进行血凝试验。6株病毒鸡胚尿囊液均有较高的血凝活性,其中SD-B40和HN2毒株为1∶512;HN1和SD-3220毒株为1∶1 024;SC-B44和SD-3297毒株为1∶2 048,血凝活性最高。具体结果见表1。

表1 分离株详细信息
2.2 病毒的8个基因片段扩增及序列测定
提取6株分离株的RNA,将RNA反转录为cDNA,通过PCR对6株病毒的8个基因片段进行PCR扩增,所有扩增的片段大小均与预期相符,如图1为HA基因片段,图2为NA基因片段,均连接到pMD-18T载体上,送至公司测序。
2.3 HA和NA同源性分析
将分离到的6株病毒分别与NCBI数据库下载的6株代表毒株的HA和NA序列进行同源性对比分析,结果发现,SD-3220、SD-3297、SD-B40、HN1、SC-B44毒株的HA核苷酸和氨基酸序列均与Y280毒株同源性最高,HN1分别为93.9%和95%,SD-3220为89.9%和91.2%,SD-3297为89.9%和91.2%,SD-B40为90.7%和93%,SC-B44为94.6%和96.4%;而HN2毒株核苷酸和氨基酸序列与G9毒株同源性最高,为95.7%和95.1%。6株病毒与Y439毒株的HA核苷酸和氨基酸序列同源性最低,分别为80.6%~84.5%和84.8%~88.5%,说明6株病毒与Y439谱系遗传距离较远。
NA序列同源性分析结果显示,SD-B40、SC-B44、HN1、SD-3297和SD-3220毒株NA核苷酸序列与Y280毒株同源性最高,分别为91%、94.5%、94.7%、90.5%和91.5%,HN2毒株与F/98毒株同源性最高。SC-B44、HN1、SD-3297和SD-3220毒株NA氨基酸序列与Y280和F/98毒株同为最高,分别为96.4%、96.4%、91.4%和92.3%;SD-B40毒株与Y280毒株同源性最高,为93.2%;HN1毒株与F/98毒株同源性最高,为97.6%。
2.4 8个基因片段遗传进化分析及系统进化树的构建
对6株病毒及其同源性较高的毒株和各谱系代表毒株进行遗传进化分析,同时构建了8个基因的系统发育进化树。结果显示(如图3、图4),这6株病毒均属于欧亚系,其HA基因属于Y280/G9-like谱系,NA基因均属于Y280-like谱系;6株病毒的PB1、PA、NP和NS基因均属于F/98-like谱系;在M基因和PB2基因来源上有些差异,SC-B44、HN1和HN2毒株的M基因和PB2基因均属于F/98-like谱系,而SD-B40、SD-3297和SD-3220毒株的M基因和PB2基因均属于G1-like谱系。因此,SD-B40、SD-3297和SD-3220毒株的8个基因来源符合G57基因型(S基因型)。

▲表示本研究分离株。下同。

图4 NP(A)、PA(B)、PB1(C)和PB2(D)基因片段系统进化树
2.5 HA和NA蛋白关键氨基酸位点分析
2.5.1 HA蛋白裂解位点及受体结合关键位点分析
将6株病毒的核苷酸序列翻译为氨基酸序列,与6个代表毒株HA氨基酸关键位点进行对比分析,结果见表2。HN1、SC-B44毒株的HA蛋白裂解位点为PARSSR↓GL,SD-3220、SD-3297、SD-B40毒株的裂解位点为PSRSSR↓GL,与早期各谱系代表毒株相比,第2位发生了由A变为S的突变,只有HN2毒株裂解位点为PAKSSR↓GL,第3位发生了由R变为K的突变,6株病毒的裂解位点均为非连续性碱性氨基酸,符合低致病性毒株特点。SD-3220、SD-3297、SD-B40毒株受体结合位点左沿为NGLMGR,HN1和SC-B44毒株受体结合左沿为NGQQGR,HN2毒株受体结合左沿为NGLQGR,与早期代表毒株相比,SD-3220、SD-3297、SD-B40、HN2毒株发生了Q226L突变,可能使得这些毒株对人型α-2,6唾液酸受体更具有亲和性。228位均为G,未发生突变。SD-3220、SD-3297、SD-B40毒株受体结合位点右沿为GTSTA,HN1、HN2和SC-B44毒株受体结合位点右沿为GTSKA,与早期代表毒株相比,SD-3220、SD-3297、SD-B40毒株发生了K/A141T突变。

表2 HA蛋白裂解位点、受体结合位点左沿和受体结合位点右沿分析
HA蛋白受体结合关键位点分析结果显示(如表3),在本试验中的6株病毒的HA蛋白受体结合关键位点第153、183、194、220位较为保守,均为WNLR;第190和226位变动较大,其中SD-3220、SD-3297和SD-B40毒株发生了A190T突变,HN1和SC-B44毒株发生了A190V突变。6株病毒的155位均为T,这可能会使病毒获得与人型α-2,6唾液酸受体结合的能力。

表3 HA蛋白受体结合关键位点分析
2.5.2 HA蛋白潜在糖基化位点分析
将分离的6株病毒与代表毒株的HA蛋白潜在糖基化位点进行比较分析,结果显示(如表4),6株病毒的第21、105、133、290、484、543位点较为保守,6株病毒均有该位点。HN1和SC-B44毒株共有8个潜在糖基化位点,SD-3220、SD-3297、SD-B40毒株共有7个潜在糖基化位点,HN2毒株HA的潜在糖基化位点仅有6个。相对于代表毒株,HN1和SC-B44毒株发生了137位由S、T或D变为N的突变,增加了这一糖基化位点;SD-3220、SD-3297、SD-B40毒株发生了P307S突变,增加了这一糖基化位点;SD-3220、SD-3297、SD-B40毒株发生了T212I位突变,缺失了这一糖基化位点;HN2毒株发生了S299R突变,缺失了这一糖基化位点。
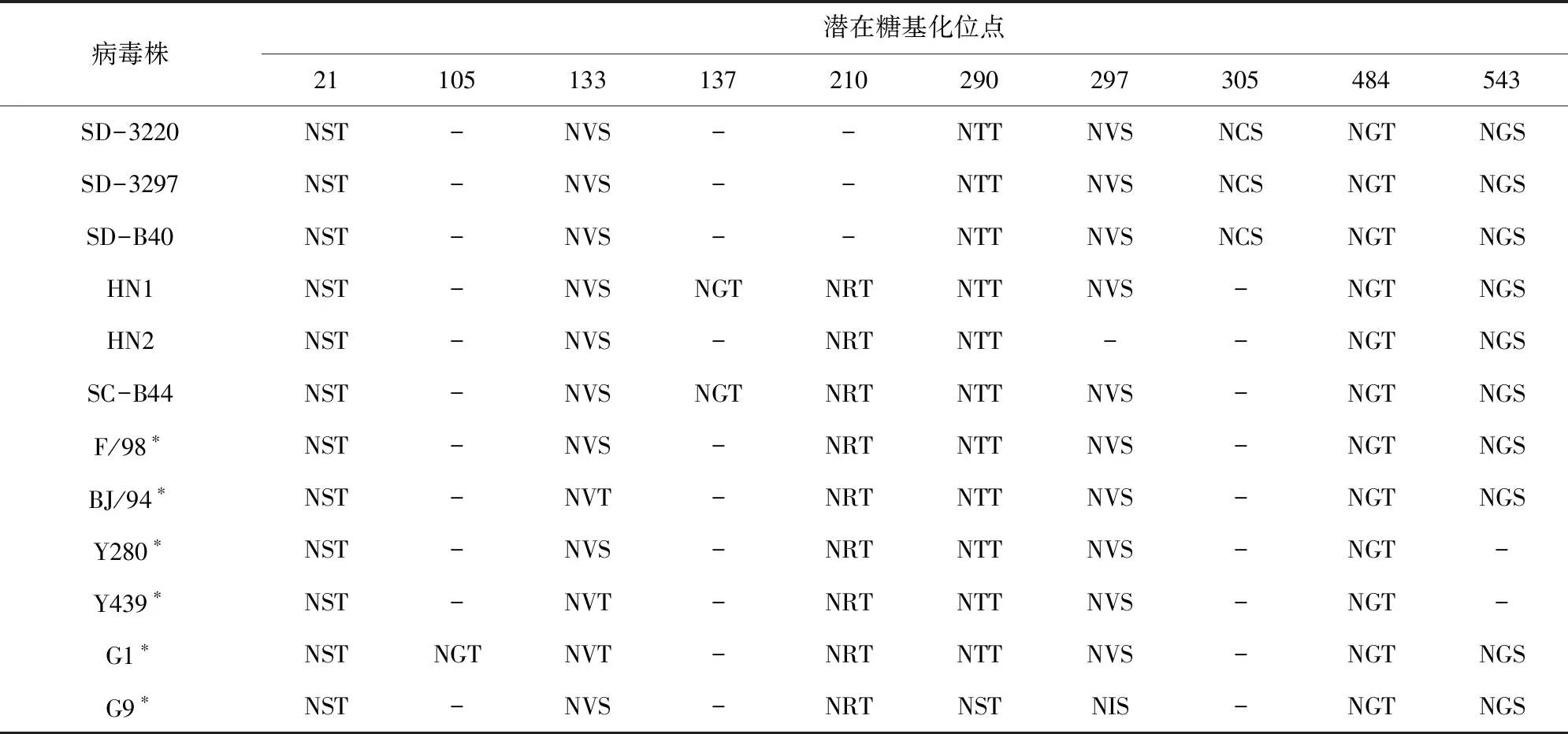
表4 HA蛋白潜在糖基化位点分析
2.5.3 NA蛋白关键氨基酸位点分析
对H9N2亚型AIV的6株代表毒株和6株分离株的NA蛋白关键氨基酸位点及潜在糖基化位点进行了对比分析,结果发现(如表5),6株分离株均有62~64位氨基酸的缺失,在6株代表毒株中,Y280和F/98毒株有62~64位氨基酸的缺失。这2个毒株所代表的谱系在我国近20年的流行中占主要地位。6株分离株与代表毒株相比,部分NA糖基化位点较为保守,SD-B40、SD-3297和SD-3220增加了368这一糖基化位点;除了HN2毒株,其余毒株失去了402这一糖基化位点。NA蛋白的红细胞结合位点分析结果显示,6株病毒的431~433位均为PQE;368和369位氨基酸变化较大,SD-B40发生了D401G突变。

表5 NA蛋白关键氨基酸位点分析
3 讨论
H9N2亚型AIV自1992年在我国广东省首次分离后,成为目前我国禽类中分布最广泛且流行率最高的AIV亚型[11-12]。H9N2亚型AIV容易将内部基因重新组合到其他亚型的流感病毒中,成为变异病毒基因组的供体[13]。新型重组毒株可能在家禽与家禽之间、家禽与哺乳动物之间、哺乳动物与哺乳动物之间进行传播,对家禽业的健康发展和公众健康安全造成极大威胁[14-15]。近年来,我国人感染H9N2亚型AIV的病例逐渐增多,血清学监测显示,人类对H9N2亚型AIV的易感程度高于H5和H7亚型AIV。因此,加强对H9N2亚型AIV的监测具有重要意义。
在我国H9N2亚型AIV流行的谱系分为几个阶段,在2000年之前主要为BJ/94-like谱系,随后逐渐被F/98-like谱系取代,从2010年开始F/98-like谱系逐渐被Y280-like谱系取代,直到现在我国普遍流行的谱系仍然为Y280-like。本研究中的6株病毒均属于欧亚系,其HA和NA基因均属于Y280/G9-like谱系,现有的人类感染的H9N2亚型AIV数据显示,Y280/G9-like谱系是大多数人类病例的来源[16]。6株病毒的PB1、PA、NP和NS基因均属于F/98-like谱系,其中SD-B40、SD-3297和SD-3220毒株的M基因和PB2基因均属于G1-like谱系,因此,SD-B40、SD-3297和SD-3220毒株的8个基因来源符合G57基因型。该基因型在2007年出现并逐渐在鸡群中稳定下来,成为当前H9N2亚型AIV流行的最主要基因型[17]。该基因型的HA和NA基因均来源于BJ/94-like谱系(Y280-like谱系)[18],内部基因PB1、PA、NP和NS以F/98-like谱系作为骨架,PB2基因和M基因由F/98-like谱系转变为G1-like谱系。而SC-B44、HN1和HN2毒株的M基因和PB2基因来源于F/98-like谱系,这可能与这些毒株分离年份较早有关。
HA蛋白介导流感病毒附着在细胞唾液酸受体上并与宿主细胞膜融合[11],HA裂解位点氨基酸序列不同,可影响病毒的组织趋向性、扩散性、致病性。HPAIV的HA裂解位点存在连续4个以上碱性氨基酸(-RRRR-),可被宿主不同组织细胞中广泛存在的多种蛋白酶裂解,引起宿主全身感染[19-20]。LPAIV的HA裂解位点一般只有单一的碱性氨基酸(-R-)[21],只有特异蛋白酶才能识别出这种结构的HA蛋白并使之发生裂解,所以病毒的增殖仅限于呼吸道和消化道等[20]。本试验所分离的6株病毒的HA裂解位点均为非连续性碱性氨基酸,符合LPAIV毒株特点。
当前H9N2亚型AIV的HA蛋白已获得优先与人型α-2,6唾液酸受体结合的能力[14],其中Q226L和G228突变发挥了关键作用,是跨物种传播的先决条件[1,22-23]。在2016—2020年我国流行的H9N2亚型AIV中,99.5%毒株HA蛋白226位为L,表明当前我国流行毒株普遍增加了病毒跨宿主感染哺乳动物的风险[24]。本研究中SD-3220、SD-3297、SD-B40、HN2毒株的HA蛋白发生了Q226L突变,推测可能具有感染人的潜在风险,但6株病毒的228位并未发生突变。有研究表明,A190V突变可以增强病毒对宿主细胞膜的吸附能力及在小鼠体内的复制能力[10]。HA蛋白的A190V突变不影响病毒与受体结合特异性,但会增强与小鼠和人类肺组织细胞受体结合的亲和力,190V对人类具有高度亲和力,190T次之,190A最低。同时,HA蛋白的A190V突变也是H9N2亚型AIV跨越物种屏障感染哺乳动物的重要决定因素之一[10,25]。在本研究中,SD-3220、SD-3297和SD-B40的190位为T。SC-B44和HN1毒株的HA蛋白发生了A190V突变,病毒与小鼠和人肺组织细胞受体结合的亲和力较强,存在引起流感大流行的风险。
HA蛋白的糖基化对防止病毒蛋白被水解、阻碍宿主抗体识别、维持蛋白三维结构的稳定性等方面发挥着重要作用[5,21,26]。本研究中HN1和SC-B44毒株的HA蛋白发生了S/D137N突变,增加了这一糖基化位点,可能导致毒力增强及抗原变异[27]。本研究中所有毒株均有133位糖基化位点,可以增强病毒对SPF鸡的毒力、组织脏器复制能力、病理损伤及体外排毒[28],SD-3220、SD-3297、SD-B40毒株拥有305位糖基化位点,但210位糖基化位点缺失。HA蛋白305位和210位糖基化位点的增加可以提高病毒与宿主受体的结合能力[29]。HA蛋白第210位糖基化位点的缺失降低了病毒结合鸡红细胞的能力,提高了病毒HA蛋白的热稳定性及病毒对鸡胚的致死性[30]。
NA蛋白茎部的氨基酸缺失可扩大H9N2亚型AIV的宿主范围,对小鼠更易感,还可以影响病毒的毒力[31]。本研究中的6株病毒的NA均有62~64位氨基酸的缺失,这也可能是我国H9N2亚型AIV在我国广泛流行传播的原因之一。NA茎部缺失可能是H9N2亚型AIV适应性突变其中的一种。SD-B40、SD-3297和SD-3220增加了368这一糖基化位点。除了HN2毒株,其余毒株失去了402这一糖基化位点,2个糖基化位点对病毒的影响需要进一步研究。根据红细胞结合位点分析,6株病毒的431~433位点较为保守,而368和369位氨基酸变化较大。在亚洲分离的H9N2亚型AIV的红细胞结合位点一直处于特定的选择压力下,因此会产生累积突变,从而提高了病毒的适应性[25]。
随着对H9N2亚型AIV研究的深入,发现了H9N2亚型AIV的宿主范围不断扩大,其具有内部基因较高的相容性,从而致使其在哺乳动物中的致病性和传播性都在不断增强[23]。本研究的6株H9N2亚型AIV均为LPAIV,其HA和NA蛋白发生了哺乳动物适应性突变,增强了对哺乳动物甚至对人的易感性。因此,我们需要对H9N2亚型AIV的跨物种传播的能力保持充分警惕,并采取更多预防措施来防止H9N2亚型AIV的传播,减少对家禽及人类的威胁。
猜你喜欢
艺术品鉴(2022年16期)2022-07-09
科学大观园(2022年2期)2022-01-23
现代畜牧科技(2021年6期)2021-07-16
河池学院学报(2021年1期)2021-07-10
景德镇陶瓷(2021年1期)2021-03-24
东方考古(2017年0期)2017-07-11
动物医学进展(2015年10期)2015-12-07
医学研究杂志(2015年12期)2015-06-10
特产研究(2014年4期)2014-04-10
郑州大学学报(理学版)(2014年3期)2014-03-01
