
菌酶协同发酵对鱼废弃物中氨基酸组成及含量的影响
2024-01-26 03:59张治军曾建华陈庭勇潘孝忠
湖南农业科学 2023年12期
雷 菲,唐 丽,张治军,曾建华,陈庭勇,潘孝忠
(海南省农业科学院农业环境与土壤研究所,农业农村部海南耕地保育科学观测实验站,海南省耕地保育重点实验室,海南 海口 571100)
鱼废弃物包括鱼头、鱼尾、鱼皮、鳞片、骨头、内脏、死亡和损坏的鱼等固体废弃物以及由血液和粘液等组成的液体废物[1]。据统计,全球每年产生的鱼废弃物超过2 000 万t[2],约20%被深加工成肽、鱼油和胶原蛋白等物质,其余大部分被倾倒、焚烧或作为废弃物排入江海[3],造成了大量的资源浪费和环境风险。然而,鱼废弃物含有丰富的粗蛋白、粗脂肪和矿质营养元素,可作为原料加工成肥料或饲料,不仅可避免资源的浪费,而且可以大大提升鱼废弃物的利用价值[4]。
目前,鱼废弃物加工方式主要包括酶解和发酵。其中,酶解常用的酶有木瓜蛋白酶、中性蛋白酶、碱性蛋白酶、风味蛋白酶等[5-6]。孙静[7]采用动物蛋白酶和碱性蛋白酶酶解鲣鱼加工废弃物得到鱼蛋白降解液,浓缩后添加耐镉菌株粉末制成富集镉的鱼蛋白有机液肥;陈杰[8]在罗非鱼下脚料中添加复合酶BJ 制成氨基酸液体肥。而目前常用的发酵技术有单菌发酵技术和混菌发酵技术。研究发现,采用戊糖片球菌发酵鱼糜,可抑制腐败菌和致病菌的生长,还能提高鱼糜中的低分子肽、非蛋白氮和游离氨基酸的含量[9-10];吴燕燕等[11]用戊糖片球菌、清酒乳杆菌以及肉葡萄球菌混合发酵鳜鱼,可缩短发酵时间,改善鳜鱼发酵的品质;刘书来等[12]用植物乳杆菌和产朊假丝酵母分段发酵鲭鱼加工废弃物,进一步提升了鲭鱼加工废弃物的游离氨基酸含量。随着技术的发展,学者们提出了菌酶协同发酵的方式。该方式可将发酵与酶解有机结合,有效缩短发酵时间,提升发酵效率[13]。例如:柯玉原[14]用木瓜蛋白酶和产朊假丝酵母联合发酵鲭鱼加工废弃物,发酵产物中的氨基态氮含量较不加酶提升了19.51%。
为提高鱼废弃物发酵产物中游离氨基酸的含量,研究以鲭鱼废弃物为原料,以团队前期研究筛选得到的最佳菌种(戊糖片球菌)为第一阶段发酵菌种,将酵母菌和产朊假丝酵母分别与木瓜蛋白酶和风味蛋白酶联合进行第二阶段发酵,通过比较分析发酵产物中游离氨基酸的变化,筛选出最佳的发酵方式,为鱼废弃物发酵提供技术支撑。
1 材料与方法
1.1 试验材料
试验所用鲭鱼废弃物购于海南省文昌市;戊糖片球菌购于威明海思(山东)生物工程有限公司,菌活力≥1.0×1010CFU/g;产朊假丝酵母菌购于陕西柏科生物科技有限公司,菌活力≥2.0×1010CFU/g;酵母菌购于陕西柏科生物科技有限公司,菌活力≥2.0×1010CFU/g;木瓜蛋白酶购于陕西柏科生物科技有限公司,酶活力1.0×106U/g;风味蛋白酶购于陕西柏科生物科技有限公司,酶活力1.0×106U/g。葡萄糖购于广东光华科技股份有限公司,分析纯。
1.2 试验方法
1.2.1 试验设计 试验设7 个发酵处理,分别为:CK,只添加戊糖片球菌进行单菌发酵;FJ,添加戊糖片球菌、酵母菌进行分段发酵;FJM:添加戊糖片球菌、酵母菌和木瓜蛋白酶进行菌酶协同分段发酵;FJF,添加戊糖片球菌、酵母菌和风味蛋白酶进行菌酶协同分段发酵;FC,添加戊糖片球菌、产朊假丝酵母菌进行分段发酵;FCM,添加戊糖片球菌、产朊假丝酵母菌和木瓜蛋白酶进行菌酶协同分段发酵;FCF,添加戊糖片球菌、产朊假丝酵母菌和风味蛋白酶进行菌酶协同分段发酵。每处理重复4 次。
1.2.2 发酵工艺 将鱼废弃物洗净切碎后用打浆机碎成鱼糜状,将30 g 鱼糜和10 mL 去离子水加入三角瓶中,75℃巴氏灭菌20 min,制成发酵基质。发酵分为2 个阶段,第一阶段为无氧发酵,在发酵培养基中加入戊糖片球菌0.4 g 和高温灭菌的200 g/L葡萄糖溶液10 mL,在恒温摇床30℃无氧发酵36 h;第二个阶段为有氧发酵,加入酵母菌0.4 g 或者产朊假丝酵母0.4 g,木瓜蛋白酶0.03 g 或者风味蛋白酶0.03 g,在恒温摇床30℃有氧发酵66 h。发酵结束后取出样品,置于-20℃冰箱冷藏备用。
1.2.3 测定项目及方法 试验中游离型氨基酸种类和含量采用超高效液相色谱和高分辨质谱法测定。(1)样品检测前处理。将10 mL 样本稀释80 倍,吸取100 mL 到离心管中,加入2 倍体积乙腈,涡旋混匀,-20℃静置30 min;12 000 r/min 离心10 min,取上清;加入等体积的0.2 mol/L 盐酸,涡旋混匀,室温条件下提取1 h;12 000 r/min 离心10 min,取上清;取10 µL 样品到瓶中,加入70 µL AccQ·Tag Ultra Borate 缓冲液和20 µL AccQ·Tag 试剂,再将反应混合物在55℃加热10 min,冷却后上机检测。(2)色谱条件。色谱柱为Waters BEH C18(50 mm×2.1 mm,1.7 µm);流动相A 为超纯水(含0.1%甲酸),流动相B 为乙腈(含0.1%甲酸),进行梯度洗脱(0 min 乙腈5%,5.5 min 乙腈10%,7.5 min 乙腈25%,8 min 乙腈60%,8.5 min 乙腈5%,13 min 乙腈5%);流速0.5 mL/min;柱温55℃;进样量1 µL。(3)质谱分析。色谱分离后采用美国Thermo 公司的Q Exactive 高分辨质谱检测系统进行质谱分析和数据采集,利用Trace Finder 软件处理质谱数据,以外标法定量。
1.3 数据处理
采用Microsoft office Excel 2007 软件对原始数据进行整理计算,采用SPSS 27.0 软件对数据进行统计分析,采用Origin 2021 和Microsoft office Excel 2007 软件绘制图表。
2 结果与分析
2.1 不同发酵方式对鱼废弃物中游离氨基酸种类及含量的影响
由表1 可知,从各处理发酵产物中均检测出21 种游离氨基酸, 其中谷氨酸族6 种,天冬氨酸族6 种,丙酮酸族3 种,丝氨酸族2 种,组氨酸族1 种,芳香族3 种。鱼废弃物经过不同酶、菌剂发酵后,各组的游离氨基酸含量存在一定的差异。与CK 相比,FC 的组氨酸、4-羟基-L-脯氨酸和天冬氨酸含量显著上升,分别增加了14.60%、50.81%和237.21%;FJ 的天冬酰胺、谷氨酰胺、丝氨酸、甘氨酸、天冬氨酸、谷氨酸、苏氨酸和丙氨酸含量均显著上升,分别增加了8 051.65%、2 016.39%、2 196.82%、38.86%、280.71%、99.36%、2 391.08%和52.90%;FCF 的甘氨酸、天冬氨酸和丙氨酸也显著上升,分别增加了32.45%、321.94%和53.18%;FCM 有8 种氨基酸较CK 显著上升,增加范围为19.66%~407.70%,其中天冬氨酸的上升幅度最大;FJF 有15 种氨基酸较CK 显著上升,增加范围为19.77%~9 843.60%,其中天冬酰胺的上升幅度最大;FJM 有16 种氨基酸较CK 显著上升,增加范围为18.79%~8 950.21%,天冬酰胺的上升幅度最大。

表1 不同发酵方式鱼废弃物中游离氨基酸的种类及含量 (µg/mL)
2.2 不同发酵方式对鱼废弃物中游离氨基酸总量和相对含量的影响
由图1 可知,与CK 相比,其他处理发酵产物中的游离氨基酸总量均显著增加,幅度为12.84%~62.50%,其中FJM 的游离氨基酸总量最高,为23 451.59 µg/mL;其次为FJF,游离氨基酸总量为20 673.52 µg/mL;第三为FCM,游离氨基酸总量为18 990.34 µg/mL。不同发酵方式对鱼废弃物发酵产物中游离氨基酸的相对含量也有一定影响,由图2可知,CK、FC、FJ 和FCF 中游离氨基酸相对含量最高为组氨酸,FCM、FJF 和FJM 中游离氨基酸相对含量最高为赖氨酸,组氨酸和赖氨酸占各组总氨基酸的比例为24.37%~32.62%。与CK 相比,FC 和FCF 的丙氨酸相对含量增加幅度最大,分别为1.52%和2.67%;FJ、FJF 和FJM 的谷氨酰胺相对含量增加幅度最大,分别为4.57%、4.73%和3.51%;FCM的赖氨酸相对含量增加幅度最高,为3.97%。
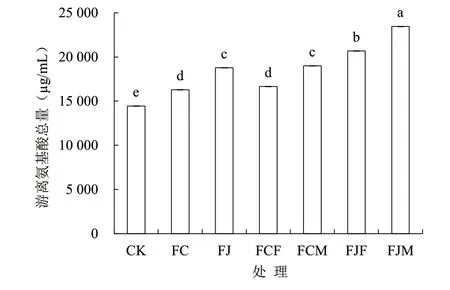
图1 不同发酵方式的鱼废弃物游离氨基酸总量
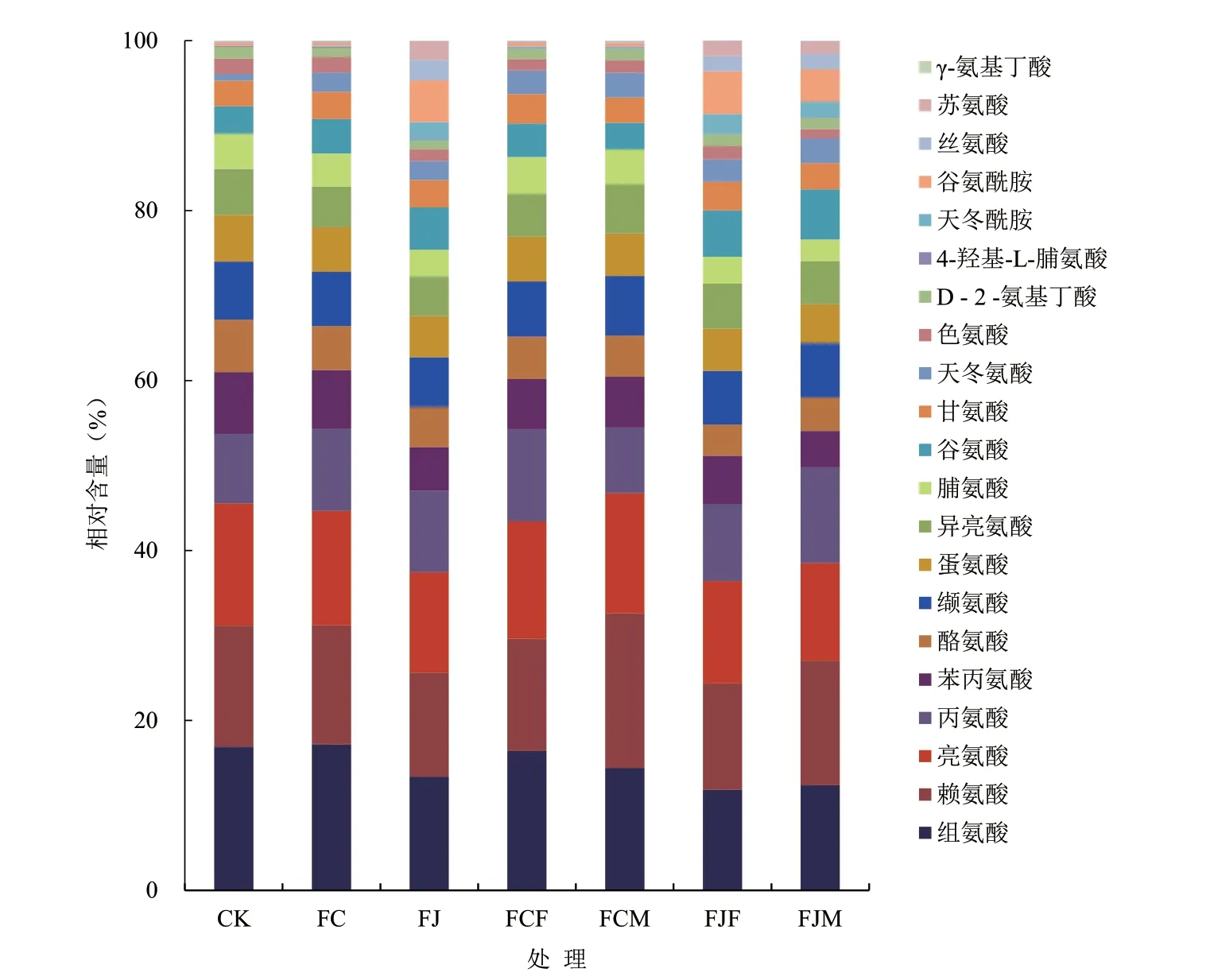
图2 不同发酵方式的鱼废弃物游离氨基酸相对含量
2.3 样品游离氨基酸的主成分分析
利用SPSS 27.0 软件先对数据进行标准化处理,再进行主成分分析,得到表2 和表3。由表2 和表3可知,前4 个主成分的初始特征值均大于1,累积贡献率为94.05%,因此选择前4 个主成分进行后续分析。第一主成分的方差贡献率为51.81%,主要影响因子为甘氨酸、谷氨酸、蛋氨酸、缬氨酸和异亮氨酸(载荷系数≥0.92);第二主成分的方差贡献率为23.18%,主要影响因子为组氨酸、 -氨基丁酸和脯氨酸(载荷系数≥0.72);第三主成分的方差贡献率为13.39%,主要影响因子为苯丙氨酸和色氨酸(载荷系数≥0.83);第四主成分的方差贡献率为5.66%,主要影响因子为组氨酸和4-羟基-L-脯氨酸(载荷系数≥0.48)。
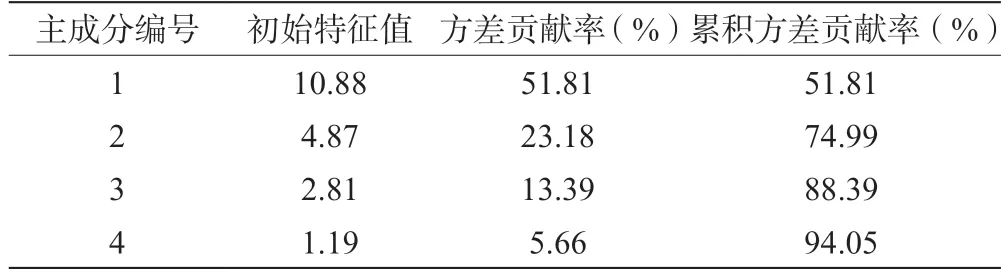
表2 主成分的特征值及方差贡献率
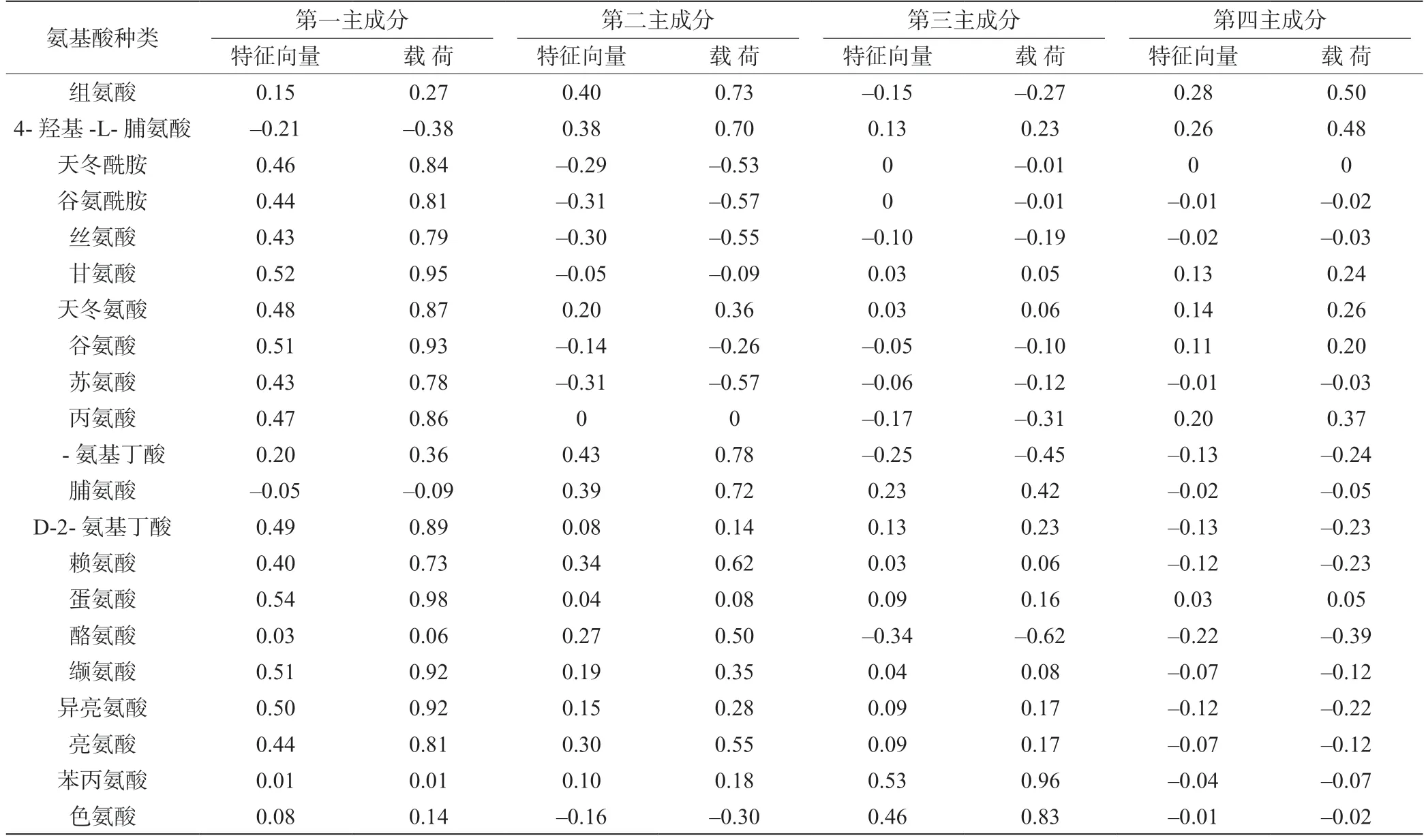
表3 主成分特征向量及载荷分析
以每个主成分对应的方差贡献率与4 个成分累计贡献率的比值为权重系数,建立鱼废弃物发酵液游离氨基酸分类评价模型F= 0.55F1+0.25F2+0.14F3+0.06F4,计算各处理鱼废弃物发酵液的综合得分,以反应各样本经不同酶和菌发酵对样品产生游离氨基酸的贡献。由表4 所示,CK 的综合得分为-4.57,FC、FJ、FCF、FCM、FJF 和FJM 的综合得分均高于CK,说明经过不同酶和菌发酵处理后,鱼废弃物的游离氨基酸含量均得到了提升。FJM 的综合得分最高,为5.36,说明添加戊糖片球菌、酵母菌和风味蛋白酶进行菌酶协同分段发酵对鱼废弃物中游离氨基酸的形成最有利。

表4 不同发酵方式样品综合得分及排序
2.4 聚类分析
为了进一步分析不同发酵方式对鱼废弃物发酵产物中氨基酸含量的影响,利用SPSS 27.0 软件对数据进行了聚类分析,结果如图3 所示。当欧式距离大于5、小于10 时,所有样品可聚为4 类,第一类为CK、FC 和FCF;第二类为FCM;第三类为FJ、FJF;第四类为FJM,说明同一类样品中的氨基酸含量最为相似。当欧式距离大于15、小于20 时,所有样品可聚为2 大类,其中CK、FC、FCF 和FCM归为第一大类,FJ、FJF 和FJM 归为第二大类,说明同一大类样品中的氨基酸含量具有一定的相似性。当欧式距离为25 左右时,所有样品才归为一类,说明第一大类和第二大类样品的氨基酸含量相似度较低。由此可知,与CK(戊糖片球菌单菌发酵)相比,分段发酵以及在分段发酵基础上增加酶进行菌酶协同发酵对鱼废弃物发酵产物中氨基酸含量都产生了一定的影响,其中2 次发酵菌种的选择对氨基酸含量影响较大,这与主成分分析的结果基本一致。
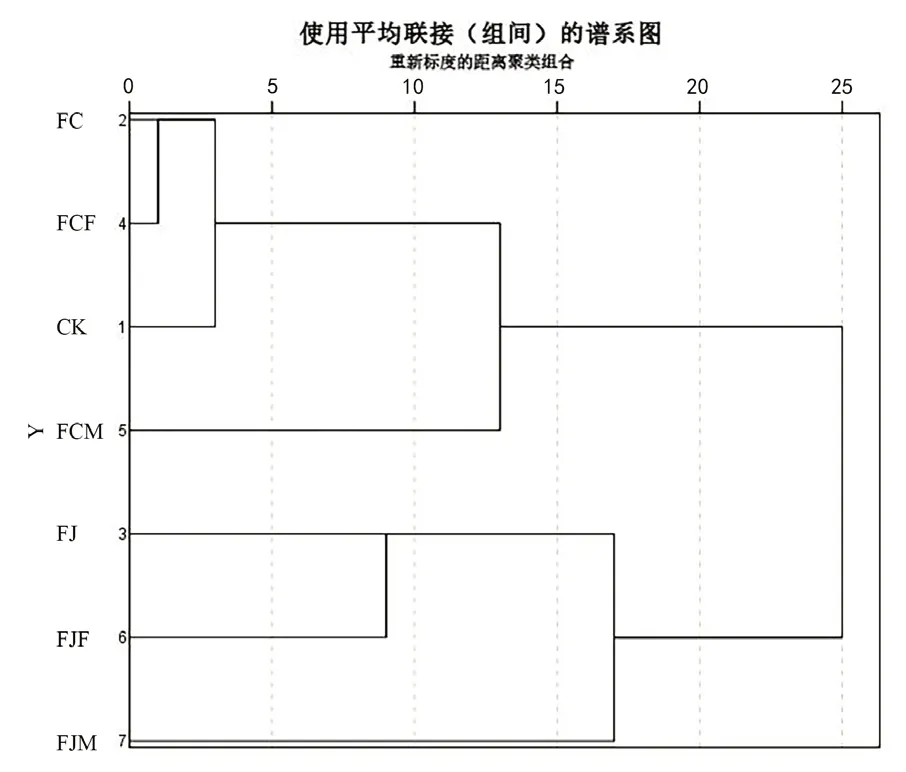
图3 不同发酵方式的鱼废弃物聚类分析树状图
3 小结与讨论
试验结果表明,采用分段发酵和菌酶协同发酵均能显著提高鱼废弃物发酵产物中的游离氨基酸含量, 与CK 比,FC、FJ、FCF、FCM、FJF 和FJM处理的发酵产物中有3~16 种氨基酸含量显著上升,总氨基酸含量增加了12.84%~62.50%,其中FJM 的总氨基酸含量最高。通过主成分分析和聚类分析发现,添加戊糖片球菌、酵母菌和风味蛋白酶进行菌酶协同分段发酵对鱼废弃物中游离氨基酸的形成最为有利。
采用微生物分段发酵能提高营养物质转化及次生代谢产物积累量[12,15-16]。刘书来等[12]用植物乳杆菌和产朊假丝酵母进行分段发酵,第二阶段发酵产物中氨基态氮含量较第一阶段提高了43.33%。试验结果表明,与戊糖片球菌单菌发酵相比,戊糖片球菌与产朊假丝酵母分段发酵产物中游离氨基酸总量增加了12.84%,戊糖片球菌与酵母菌分段发酵产物中游离氨基酸总量增加了30.05%。不同酵母产蛋白酶的能力不同,对蛋白分解的能力也不同,有研究发现酿酒酵母对氨基酸废液的同化效果高于产朊假丝酵母[17-18]。试验结果显示,戊糖片球菌与酵母菌分段发酵产物中的天冬酰胺、谷氨酰胺、丝氨酸、谷氨酸和苏氨酸含量均较戊糖片球菌与产朊假丝酵母分段发酵显著上升,游离氨基酸总量增加了15.25%。
菌酶协同可以共同促进底物酵解,提高蛋白消化率[17]。史路路等[19]用淀粉酶、糖化酶和酵母菌协同发酵玉米蛋白粉,发酵后总氨基酸含量增加了18.88%。笔者在分段发酵的基础上,分别添加了风味蛋白酶和木瓜蛋白酶进行协同发酵,发现游离氨基酸总量均进一步得到了提升。鱼废弃物中的蛋白质在蛋白酶的作用下可酶解成氨基酸、小分子肽等物质,但不同蛋白酶的酶解效果不同。关平彦等[20]发现风味蛋白酶酶解金枪鱼下脚料的效果优于胰蛋白酶和中性蛋白酶。该试验中,与FC 处理相比,FCF 和FCM 处理发酵产物中的游离氨基酸总量分别增加了2.23%和16.61%;与FJ 处理相比,FJF 和FJM 处理发酵产物中的游离氨基酸总量分别增加了10.15%和24.95%,说明此试验中木瓜蛋白酶的酶解效果优于风味蛋白酶。
猜你喜欢
军事文摘·科学少年(2021年1期)2021-02-04
国际呼吸杂志(2019年1期)2019-01-28
现代检验医学杂志(2016年2期)2016-11-14
环境科技(2016年3期)2016-11-08
兽医导刊(2016年12期)2016-05-17
阅读与作文(小学低年级版)(2015年8期)2015-05-30
化学工业与工程(2015年1期)2015-02-10
食品工业科技(2014年9期)2014-03-11
食品科学(2013年19期)2013-03-11
化学工业与工程(2012年6期)2012-02-10
