
蜂糖李需冷量与需热量研究*
2024-01-22 11:09:10马玉华彭志军周俊良吴亚维
云南农业大学学报(自然科学) 2023年6期
张 敏,马玉华 ,赵 凯,彭志军,周俊良,吴亚维
(贵州省农业科学院 果树科学研究所,贵州 贵阳 551007)
蜂糖李为贵州本土李品种,因其甘甜脆爽、酸涩感低而深受市场青睐。自2016 年审定以来,其栽培面积不断扩大,至2023 年,贵州省蜂糖李种植面积达5.3×104hm2,成为贵州省内种植面积最大的果树单品。但该品种在多地存在产量不稳定和产量低的问题,研究者较多从栽培技术[1]和授粉受精[2]等角度进行探索,鲜有研究关注其需冷量和需热量。蜂糖李等落叶果树冬季需要一定的低温量以解除休眠[3],休眠后需要一定的热量以正常开花结果[4-5]。低温量得不到满足,往往引起生长发育障碍,严重影响产量[6-7],故其休眠及解除关乎结果和树体生长,是果实生产的关键因素[8]。需冷量积累不足是贵州等南方产区李坐果难、坐果率低和产量低的重要原因[9-12],冷蓄积量是广州李栽培的限制因子,也是区分适宜区的考量指标[13]。
常用的需冷量估算模型有0~7.2 ℃模型(又称冷量小时模型、Weinberger 模型,单位:冷量小时,chill hours,CH)、犹他模型(单位:冷量单元,chill units,CU)和动态模型(单位:冷量组分,chill portions,CP)[4-5,14-16]。中国多用0~7.2 ℃模型和犹他模型进行估算,在不同地区、不同品种的桃[17-18]、李[19-20]、杏[20-21]适用模型各不相同;针对李[4]和果梅[22]的研究,也有动态模型适用的报道。受年际和发育阶段等多种因素的影响,很难确定一个适用于所有地区和品种的估算模型。总体而言,中国李的需冷量低于欧洲李,倾向于更早开花[23],但也易受霜害影响[24]。需热量模型有生长度小时模型、有效积温模型、最大积温模型、温度最大值累计模型、平均温度累计模型和热量模型[25]。有效积温模型最为简单实用,在中国常用于对葡萄的研究[26-28]。生长度小时模型又可分为Anderson 模型[29]和Richardson 模型[30],前者考虑适宜温度、上限温度和下限温度,对积累的热量进行分类累加,结果更加精细准确,适用于李[4]、梨[31]和其他核果类果树[3]的研究;后者为小时温度减4.5 ℃的累加值(温度高于25 ℃、低于4.5 ℃时为0 积累),适用于李[3]和果梅[32]的研究。开花需冷量与需热量之间存在一定的消长关系[32],即冷蓄积量超过树种本身需冷量时,萌发所需热量减少(促萌时间变短)[33-34],反之,冷蓄积量达不到树种需冷量时,可通过增加萌发热量(延长促萌时间)得到弥补[35]。
目前,针对蜂糖李需冷量和需热量的研究未见报道,生产上却已经出现了引种和扩种热。多数引种未获成功,存在的问题主要包括2 种:一是向北(如河南、陕西等)扩散,冷蓄积量增加,易遭受花期低温;二是向南(如向广东、福建、云南等)扩散,冷蓄积量不足,常开花零星、挂果少。即使是在四川、云南和贵州的个别区域,盲目引种导致的低坐果率和低产量问题也已成为制约产业发展的关键问题。明确蜂糖李需冷量、需热量及果实成熟需热量对其科学引种具有重要意义,根据产业需要,课题组在前人研究的基础上,通过对离体蜂糖李枝条人工控制冷积累量、促萌(人工控制热量使其萌发)以及大田条件下蜂糖李开花需冷量和需热量、果树成熟需热量等进行研究,明确蜂糖李开花需冷量、需热量并筛选最适估算模型,分析开花需冷量与需热量的关系,计算果实成熟需热量,以期为其花期和成熟期的预测及引种布局提供科学依据。
1 材料与方法
1.1 供试材料
冷量控制试验材料于2020 年11 月24 日(此时连续3 日平均温度为12.1 ℃,根据犹他模型测算[22]冷量累计持续为负,枝条尚未达到休眠标准)采自贵州省安顺市镇宁布依族苗族自治县良田镇板袍村果园(海拔685 m,N25°36′49″,E105°50′29″),枝条带回实验室后,参照章镇等[36]的方法,用75%酒精消毒后蜡封两端,10 根枝条为1 捆,用报纸包裹后打湿,于5 ℃低温冰箱保存,其后定期取出,进行促萌试验。
田间试验材料于2021 年11 月—2022 年1 月采自贵州省农户常规管理果园,果园分别位于:紫云苗族布依族自治县火花镇破关村(海拔750 m,N25°40′42″,E106°3′27″)、黔南布依族苗族自治州惠水县断杉镇大坡村(海拔979 m,N25°50′52″,E106°30′57″)和罗甸县木引镇水井村(海拔814 m,N25°31′10″,E106°26′21″)、安顺市镇宁布依族苗族自治县六马镇弄袍村母树园(海拔907 m,N25°42′23″,E105°49′47″)和常规园(海拔890 m,N25°42′39″,E105°49′20″)。在果园放置自动数据记录器(HOBO 蓝牙温度MX2302A记录仪)收集小时温度。进入休眠后,每3~7 d 取1 次枝条,带回实验室进行促萌试验。
1.2 试验方法
1.2.1控温条件下蜂糖李离体枝条需冷量与需热量
对定期从冷藏环境中取出的蜂糖李枝条(蓄冷量不断增加)进行萌发试验,计算其萌发指数和冷量累计值。
萌发试验参考RUIZ 等[4]的方法并略作改进。剪平枝条基部,插入深约3 cm 的5%蔗糖溶液中,立即移入人工气候室培养(昼/夜温度25 ℃/18 ℃,光/暗时间16 h/8 h,光照强度2 000 lx,空气相对湿度60%~70%)。每隔3 d 换1 次蔗糖溶液,并将枝条基部剪去约2~3 mm,露出新茬,培养12 d 以积累足够热量正常萌发。热蓄积量补偿研究中,将枝条萌发时间增加至21 d,蓄冷量严重不足的增加至24 和27 d。
参考王力荣等[37]的方法,根据萌发指数确定休眠终点并略作改进。花芽分级标准:1 级,未萌动;2 级,芽鳞片张开;3 级,花蕾显露;4 级,花蕾露白;5 级,花朵开放。叶芽分级标准:1 级,未萌动;2 级,芽鳞片张开;3 级,顶尖露绿;4级,叶伸出;5 级,叶开放。当各级的芽数加权平均值等于或大于2.5 时,则采样日期为休眠终点。萌发指数(germination index,GI)计算公式为:
式中:i为花芽或叶芽萌发级别,Xi为萌芽数。需冷量为从休眠起始至休眠解除期间的小时冷量累积值,参考庄维兵等[5]和GAO 等[22]的方法,分别采用0~7.2 ℃模型、犹他模型和动态模型进行需冷量估算。
1.2.2大田蜂糖李需冷量及需热量
大田蜂糖李休眠(冷量积累)的起始日期为根据犹他模型日冷量蓄积连续3 d 为正值的日期[30],并根据李种质资源描述规范[38]确定蜂糖李盛花期、成熟期和发育时间。记录12 月1 日至3 月1 日(贵州蜂糖李冷量积累关键期)冷量积累历程,并从休眠起始后,定期(每3~7 d)对5 个果园分别取枝促萌,以估算其需冷量,估算方法同1.2.1节;根据ANDERSON 等[29]和RICHARDSON 等[30]的方法计算盛花期和果实发育期生长度小时热量(单位:GDH·℃)积累。
1.3 数据统计与分析
采用 Excel 2010 软件进行数据整理,采用SPSS 24.0 软件进行统计分析。
2 结果与分析
2.1 控温条件下蜂糖李离体枝条的需冷量与需热量
2.1.1蓄冷量对枝条萌发的影响
由表1 可知:5 ℃冷积累条件下,0~7.2 ℃模型与犹他模型估算冷量数值相同。冷积累795 h (25.8 CP,795.0 CU)时花芽通过休眠,冷积累1 154 h (36.9 CP、1 154.0 CU) 时叶芽通过休眠,表明蜂糖李叶芽需冷量远高于花芽。冷积累578 h 以内,叶芽萌发指数高于花芽;低温处理650 h 以上时,花芽萌发指数高于叶芽。就开花而言,随着低温时间的延长,花芽萌发指数增大,说明冷量积累增加,开花进程加速,且花芽萌发优势增加(图1),暗示随着冷量积累增多,开花进程中对热量的需求减少。

图1 冷量增加对蜂糖李萌芽的影响Fig.1 Effects of chilling accumulation on the bud germination of ‘Fengtang’ plum
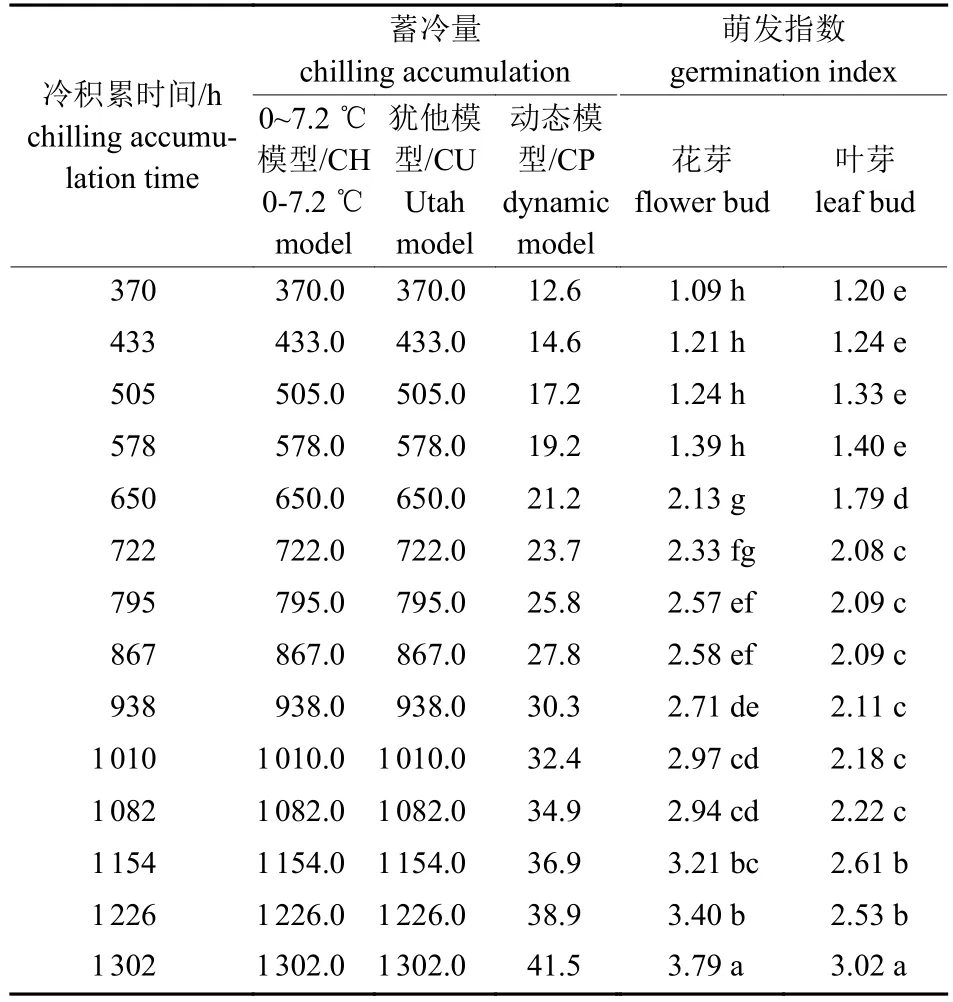
表1 蓄冷量对蜂糖李离体枝条萌发指数的影响Tab.1 Effects of chilling accumulation on the germination index of isolated ‘Fengtang’ plum branch
2.1.2蓄热量对枝条萌发的影响
由图2 可知:蓄热量对花芽萌发的影响可分3 类情况。(1)冷量严重不足,通过热补偿无法补救。如冷积累150 h 的蜂糖李枝条,保温促萌12 d(热积累量按Anderson 模型估算为6 003 GDH·℃,按Richardson 模型估算为5 668 GDH·℃)时,花芽和叶芽均无萌动;促萌时间增加到21 d (热积累量按Anderson 模型估算为10 159.7 GDH·℃,按Richardson 模型估算为9 592.0 GDH·℃)花芽萌发得到显著改善;继续积累热量至24 d (热积累量按Anderson 模型估算为11 545 GDH·℃,按Richardson模型估算为10 900 GDH·℃)和27 d (热积累量按Anderson 模型估算为12 930 GDH·℃,按Richardson 模型估算为12 208 GDH·℃)时,花芽萌发指数不再升高,仍未达到休眠解除标准。(2)冷量不足,但通过热量补偿可达到休眠解除标准。冷积累370 h 的蜂糖李枝条,热积累量增加,花芽萌发指数显著增大,保温12、21 和24 d 的花芽萌发指数差异均达到显著水平,且保温24 d 时花芽萌发指数接近休眠解除标准;冷积累433 h 及以上的蜂糖李枝条,保温促萌时间从12 d 延长到21 d,花芽萌发指数极显著增大;保温促萌12 d、冷积累505~722 h 的枝条均未达到休眠解除标准,将保温促萌时间增加至21 d 后均打破休眠。(3)需冷量得到满足后,增加热量,萌发进程加快。冷积累时间在795 h 及以上、本身达到休眠标准的枝条,增加开花热量,萌发指数也得到提升。
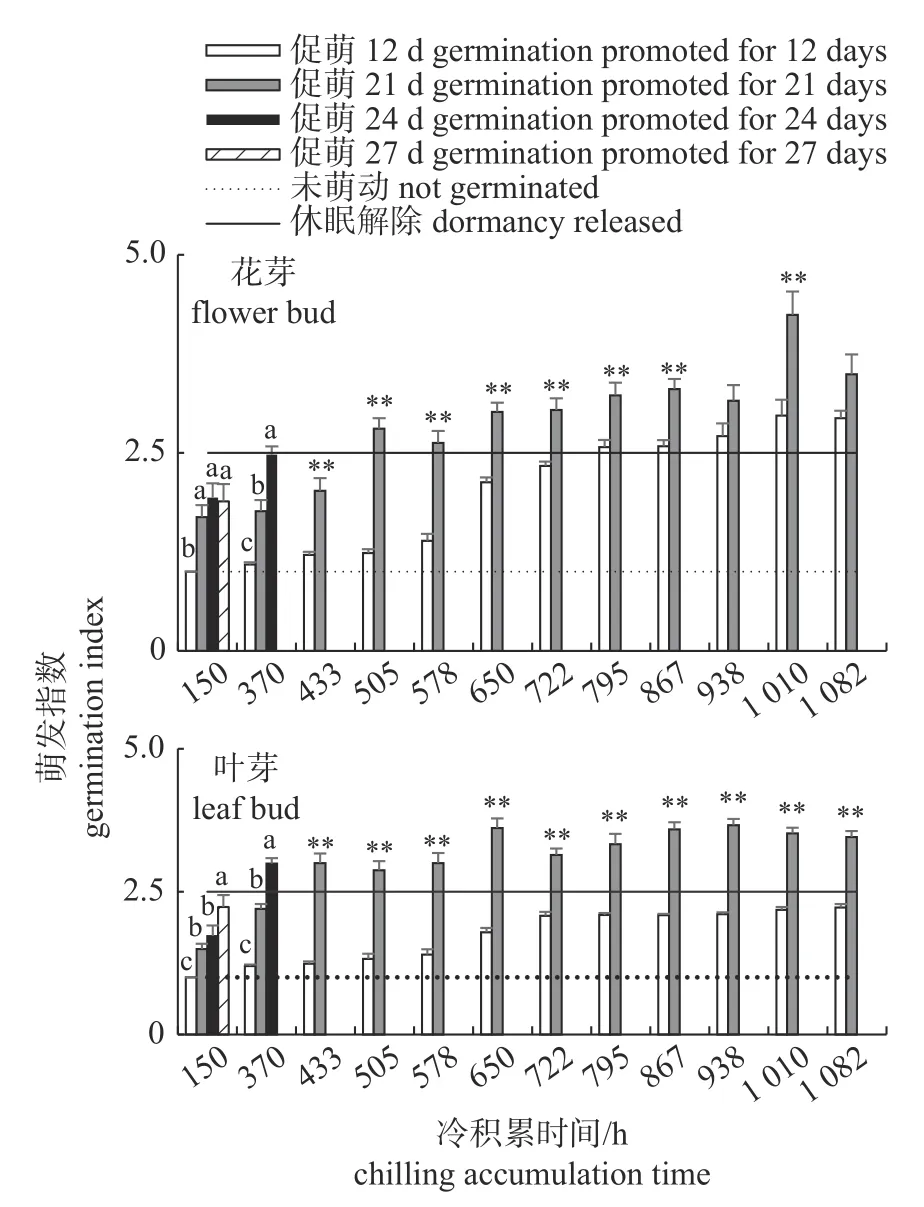
图2 热量增加对花芽和叶芽萌发指数的影响Fig.2 Effects of heat accumulation on the flower and leaf bud germination index
由图2 还可知:总体上蜂糖李叶芽需冷量高于花芽,但冷积累不足时,叶芽易通过增加热量实现休眠解除。冷积累150 h 的蜂糖李枝条增加促萌时间到27 d 时,叶芽萌发指数已接近2.5,高于花芽萌发指数;对冷积累370 h 的枝条增加热量积累,叶芽萌发指数显著增加,萌发12、21 和24 d 的叶芽萌发指数差异均达到显著水平,萌发24 d 时叶芽萌发指数超过休眠解除标准;冷积累433 h 及以上的蜂糖李枝条,萌发时间从12 d 延长到21 d,叶芽萌发指数极显著增加,且均达到休眠解除标准。
可见,在一定范围内,增加蓄热量可弥补蓄冷量的不足,促进开花,开花需冷量与需热量一定程度上存在消长关系,即:蓄冷量高,则蜂糖李开花对热需求减少;蓄热量高,则蓄冷量可相应减少;同时,蜂糖李花芽需冷量远低于叶芽需冷量,需冷量不满足时,叶芽比花芽更容易通过热量补偿实现休眠解除,如冷积累370 和433 h的枝条,补充同样热量,叶芽总是先于花芽达到休眠解除标准。
2.2 大田蜂糖李的需冷量和需热量
2.2.1大田条件下蜂糖李的主要物候期
根据大田枝条插枝情况,结合田间物候期(表2)可知:贵州主产区蜂糖李休眠期为42~54 d,平均49 d;休眠解除至开花需51~69 d,平均57 d;果实发育需109~117 d,平均114 d。2021 年11 月下旬进入休眠,2022 年3 月上旬开花,果实成熟期在6 月下旬至7 月上旬。
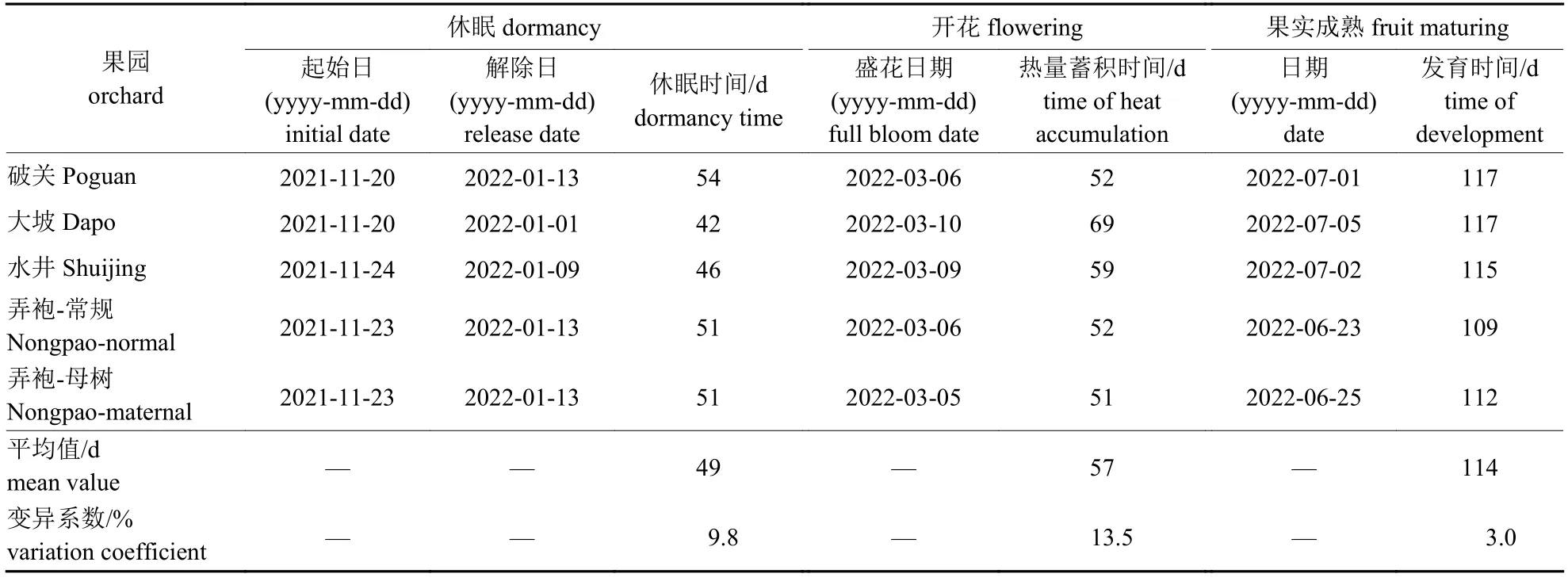
表2 不同蜂糖李果园物候期Tab.2 Phenophase of different ‘Fengtang’ plum orchards
2.2.2大田条件下蜂糖李的冷量积累
根据2021 年休眠期至2022 年3 月开花后的温度(图3),各蜂糖李果园均在12 月初就有冷量积累,但冷量积累进度各不相同(表3)。就变异系数而言,以动态模型园际间变异系数最小。5 个果园中,以大坡果园冷量积累最快,根据蜂糖李花芽通过休眠时冷积累为25.8 CP (表1),1 月1 日大坡果园蜂糖李已打破休眠;水井果园在1 月11 日前打破休眠;其余果园需冷量均在1 月21 日前满足打破休眠条件。这与田间取枝测定数据一致,表明选用符合性好的模型可直接根据田间温度进行需冷量估算以确定破眠时间。此外,各果园需冷量满足后,冷量积累仍在持续。
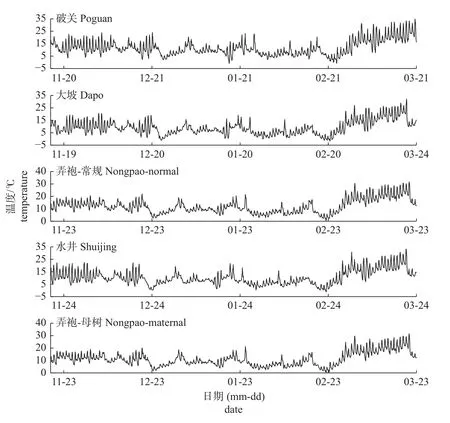
图3 果园气温Fig.3 Temperature of orchards
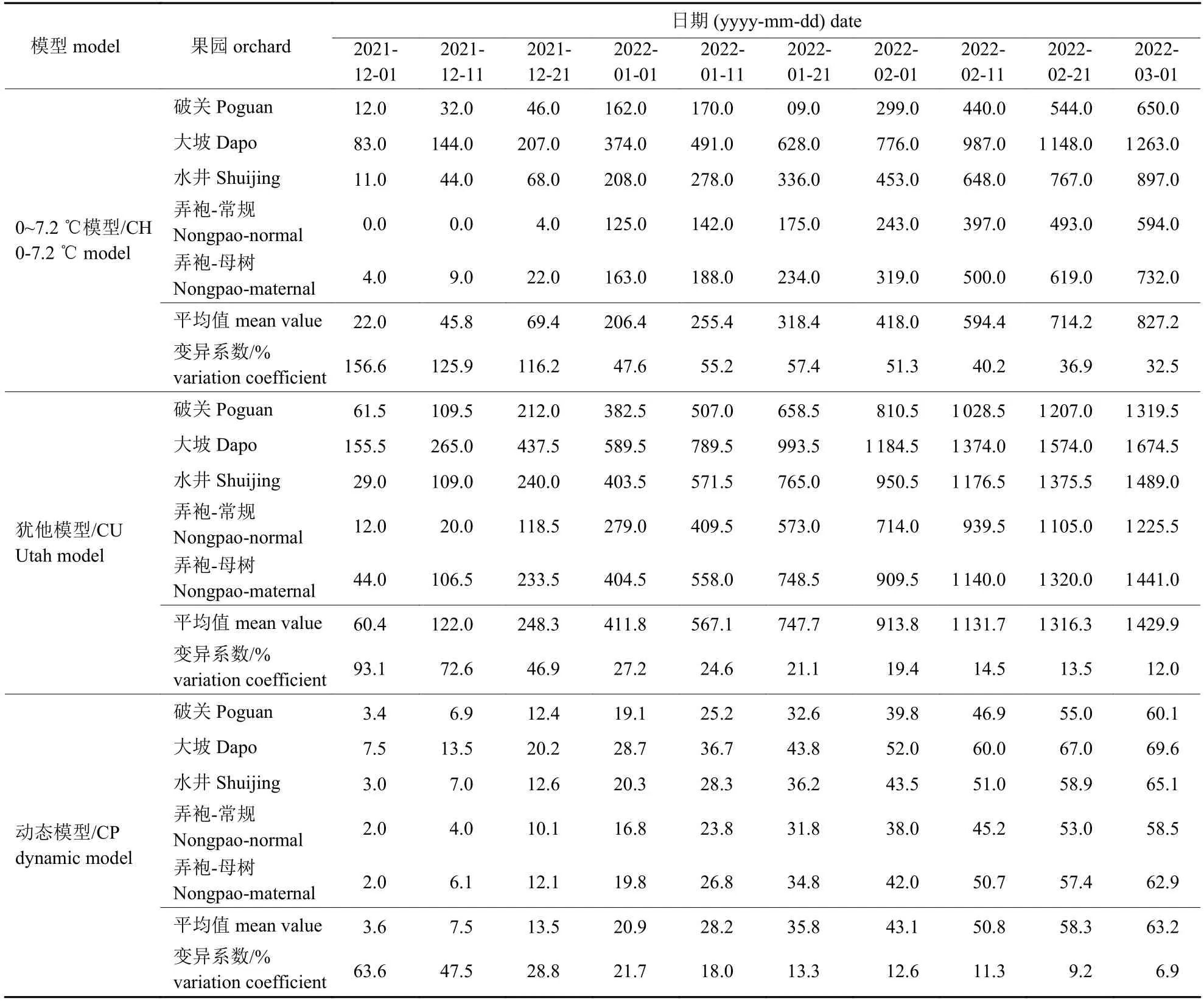
表3 2021-2022 年蜂糖李果园冷量累积Tab.3 Chilling accumulation of ‘Fengtang’ plum orchards in 2021-2022
2.2.3大田蜂糖李需冷量及其最适模型
由表4 可知:达到解除休眠标准时,按0~7.2 ℃模型估算冷量蓄积值为142.0 CH (弄袍-常规)~385.0 CH (大坡),平均为234.0 CH;按犹他模型估算冷量蓄积值为449.5 CU (弄袍-常规)~606.5 CU (弄袍-母树),平均为554.9 CU;按动态模型估算冷量蓄积值为25.8 CP (弄袍-常规)~28.9 CP (大坡),平均为27.8 CP。3 个冷量估算模型中,动态模型估算值的变异系数最小,且与插枝通过休眠数据(表1)相符,说明动态模型为蜂糖李需冷量估算的最佳模型,该模型估值以大坡果园为最高、弄袍-母树园与之接近、弄袍-常规园估值为最低。值得注意的是,弄袍-常规园与弄袍-母树园属同一行政村,在5 个果园中二者地理位置最为接近,但冷量积累差异较大,说明小气候对果园影响较大。
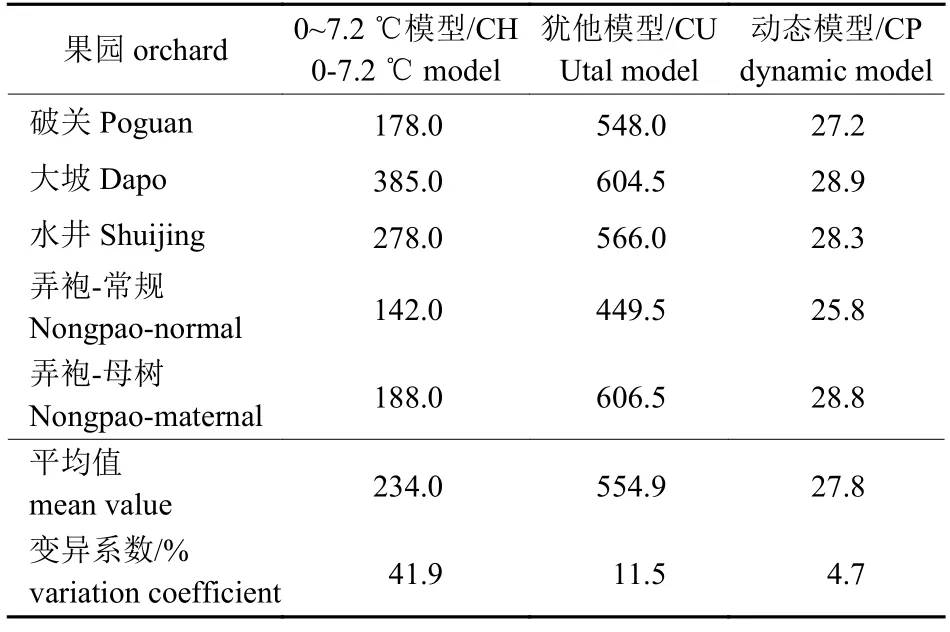
表4 蜂糖李果园通过休眠时的冷量估算Tab.4 Estimated chilling requirement of‘Fengtang’ plum orchards
2.2.4大田蜂糖李需热量及其最适模型
5 个果园蜂糖李开花需热量的估算值(表5)均以弄袍-常规园为最高、弄袍-母树园最低,Richardson 模型变异系数为8.4%;Anderson 模型变异系数为10.8%,Richardson 模型变异系数更小,且计算相对简单,更适用于蜂糖李开花需热量估算。就果实发育而言,按Richardson 模型估算的热量蓄积以破关果园最多(45 823.7 GDH·℃)、大坡果园最少(40 903.1 GDH·℃),平均需热量为42 752.5 GDH·℃;按Anderson 模型估算的热量蓄积以弄袍-母树园最多(41 892.8 GDH·℃)、大坡果园最少(40 417.1 GDH·℃),平均需热量为41 269.1 GDH·℃;Richardson 模型变异系数为4.6%,Anderson 模型变异系数为1.4%,均小于5%,说明果实成熟需热量估值相对稳定。
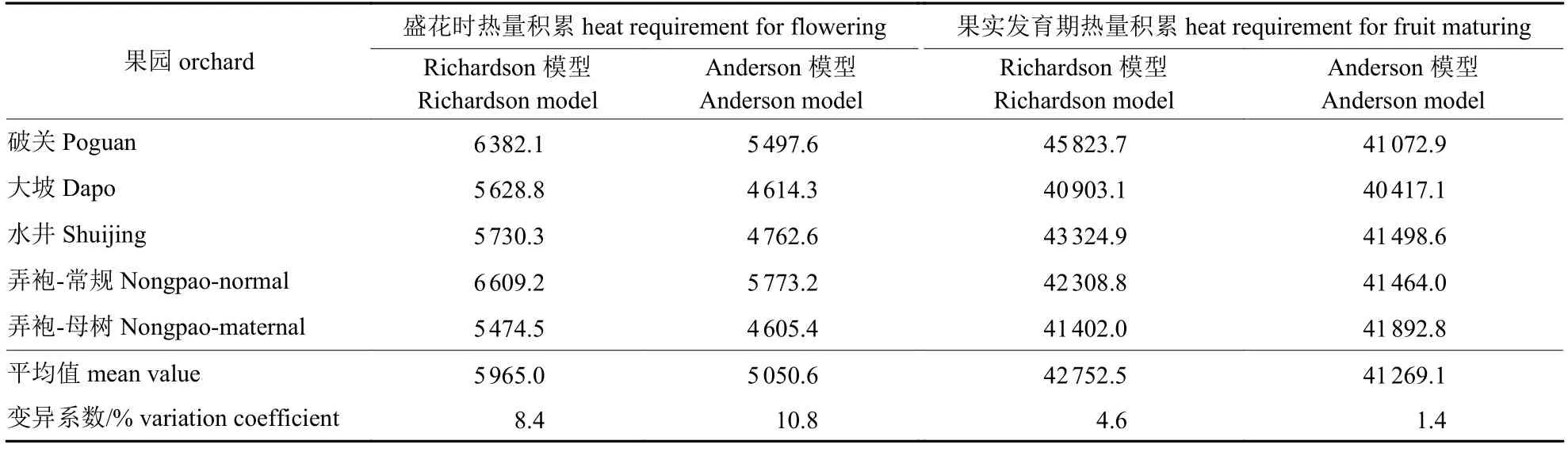
表5 蜂糖李果园热量累积的估算Tab.5 Estimated heat requirement of Fengtang plum orchards GDH·℃
2.2.5大田蜂糖李开花需冷量和需热量的相关性
由表6 可知:需冷量估算模型中,0~7.2 ℃模型与其他模型的相关性均不显著,犹他模型与动态模型呈极显著正相关、与需热量估算Anderson 模型和Richardson 模型呈显著负相关,动态模型与Anderson 模型和Richardson 模型均呈极显著负相关;需热量估算模型中,Anderson 模型和Richardson 模型呈极显著正相关。总体而言,需冷量模型估值与需热量模型估值呈负相关,这进一步验证了蜂糖李开花需冷量与需热量相互补偿的推论。
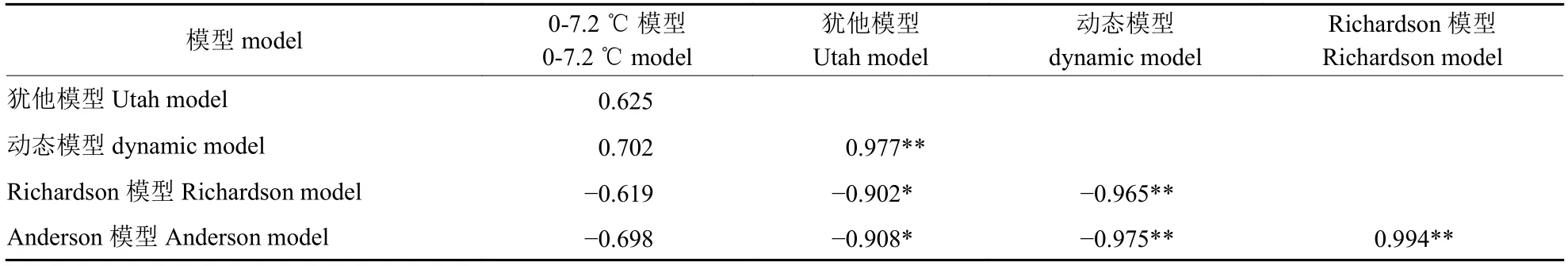
表6 蜂糖李开花需冷量和需热量的相关性Tab.6 Correlations between chilling requirements and heat requirements of ‘Fengtang’ plum flowering
3 讨论
对于包括蜂糖李在内的落叶果树,选择需冷量适宜当地的品种以确保开花结果整齐对实现产量和经济效益至关重要[4,39]。在此过程中,需冷量估算尤为关键。本研究选用国内外广泛使用的0~7.2 ℃模型、犹他模型和动态模型[3-4,14,16,22-23,35]研究蜂糖李离体枝条补充冷量、热量后的萌发状况,并对大田条件下的主产区果园需冷量进行估算,结果显示:经离体枝条控温试验,蜂糖李通过休眠时冷蓄积量测定为795.0 CH、795.0 CU和25.8 CP;田间条件下,冷量估值分别为142.0~385.0 CH、449.5~606.5 CU 和25.8~28.9 CP;3 种模型中,以动态模型的变异系数最小,且其对大田与离体控温试验估算结果符合,说明该模型估算值最接近实际。这与前人对果梅[22]和甜樱桃[39]的研究结果一致。0~7.2 ℃模型的估算值变异系数高达41.9%,说明该模型不适用于冬季偏暖的贵州蜂糖李产区,这与严娟等[40]的研究结果一致,也进一步表明筛选适宜冷量评估模型的必要性。
RUIZ 等[4]研究了11 个中国李品种的需冷量和需热量,将Pioneer 列为极低需冷量品种(22.2 CP),Red Beauty 列为低需冷量品种(34.9 CP),Black Diamond 列为中需冷量品种(42.0 CP),Golden Globe 列为高需冷量品种(55.6 CP);按犹他模型,Red Beauty 的需冷量估算为562.8 CU (3 年变化范围为500~688 CU);根据Richardson 模型,11 个品种开花需热量为5 990~9 228 GDH·℃,其中7 个品种开花需热量在7 300~8 600 GDH·℃,Red Beauty 在2011、2012 和2013 年的需热量分别为6 591、8 663 和9 099 GDH·℃。高志红等[7]对南京12 个李品种进行了研究,按犹他模型,其需冷量为810~1 245 CU,其中红美丽(引进品种Red Beauty)需冷量为930 CU。可见,同一品种的需冷量和需热量在不同地区、不同年际表现差异较大。本研究中蜂糖李需冷量为25.8~28.9 CP,开花需热量为5 474.5~6 609.2 GDH·℃,表明蜂糖李开花需冷量和需热量均较低,这可能也是蜂糖李引种难以成功、年际开花和结果表现差异较大的原因之一。向热区引种,其冷量需求难以满足;向冷区引种,其开花需冷量和需热量均可迅速达到开花要求,遭遇早春霜冻的风险极大。因此,引种蜂糖李需全面、谨慎地考虑冷量和热量条件。
本研究对蜂糖李离体枝条萌发试验结果表明:在一定条件内,同样热量下,冷量增加,开花进程加快;同样冷量下,热量增加,冷量相对不足的可达到休眠解除标准,低温量已经满足需冷量要求的枝条开花进程亦加快。贵州5 个蜂糖李果园的开花需冷量和需热量呈负相关,也印证了这一结果。可见,需冷量和需热量并非恒定量,而可能存在相互补偿机制,在一定范围内,冷量积累不够时,可以通过增加热量得到补偿。前人对甜樱桃的研究也表明:通过果园大量热量积累最高可弥补50%需冷量,并实现自然开花;冷积累量为需冷量的150%时,开花所需热量降至原来的一半[35]。谭钺等[41]对低需冷量桃的冷量控制试验结果也印证了这一观点。这可为以蜂糖李为代表的高附加值果树设施栽培下的花前温度管理提供借鉴。
本研究发现:蜂糖李花芽需冷量高于叶芽,说明其为先花后叶,但延长促萌时间,叶芽更容易通过热量补偿弥补冷量不足打破休眠,这也部分解释了在蓄冷量不足的地区,蜂糖李花叶同放甚至先叶后花的现象。花芽和叶芽需冷量和需热量之间的差异可能是其适应环境的结果,其本身树势强,长势旺,花芽先叶芽而动,增加了授粉受精成功的概率;一旦花叶同放或先叶后花,则在对树体营养的竞争中,花将处于劣势,不利于授粉受精。当前针对叶芽需冷量和需热量的研究较少,后续有必要就此开展精细研究,为在引种扩种过程中既使二者彼此补偿又确保花叶时间差提供依据。
4 结论
蜂糖李为低需冷量、低需热量品种,在贵州开花需冷量最适模型为动态模型,需冷量为25.8~28.9 CP;Richardson 模型估算的开花需热量为5 474.5~6 609.2 GDH·℃,盛花至果实成熟需热量估值40 417.1~41 892.8 GDH·℃。在一定范围内,其开花需冷量与需热量可相互补偿,并呈负相关。
猜你喜欢
中国果树(2022年1期)2022-04-20 04:57:44
小星星·阅读100分(低年级)(2021年5期)2021-08-09 23:57:39
果树实用技术与信息(2020年8期)2020-09-18 09:29:44
草原(2018年10期)2018-12-21 12:28:38
广东农业科学(2017年5期)2017-08-29 10:38:05
浙江档案(2017年1期)2017-05-16 06:40:56
国家图书馆学刊(2016年4期)2016-03-17 02:52:31
化工设计通讯(2016年10期)2016-02-10 03:04:48
小星星·阅读100分(低年级)(2015年4期)2015-05-18 07:40:27
读者(2015年5期)2015-03-04 01:04:15
