
元谋干热河谷不同植被类型对生物多样性的影响1)
2024-01-13 02:47孙泽何真敏高中腾欧朝蓉王猛李金仙孙永玉张春华
东北林业大学学报 2024年2期
孙泽 何真敏 高中腾 欧朝蓉 王猛 李金仙 孙永玉 张春华
(西南林业大学,昆明,650224)(林木资源高效生产全国重点实验室(中国林业科学研究院高原林业研究所))(云南元谋干热河谷生态系统国家定位观测研究站)
生物多样性涵盖地球上由生物和环境相互作用所形成的复杂生态系统及相关生态过程,起维持生态系统持续生产力和为人类生存提供基本条件的作用。当前,生物多样性研究是该领域备受关注的热点之一[1]。植物多样性是生物群落重要的评价指标,反映了区域内植物的物种数目、丰富度、均匀度及群落间的相互关系[2]。了解植物群落重要性对有效开发、利用、保护元谋干热河谷生态系统至关重要[3-5]。植物种类组成是群落基本特征之一,所以,分析植物种类组成是研究群落的关键[6]。作为群落的重要部分,植物种类组成影响着其生态功能[7-8],优势种更是起一定的主导作用。分析植物多样性有助于了解群落结构及动态变化[9],同时能够探究生境的复杂性,对维持生态系统平衡起重要作用[10]。
元谋干热河谷地区以其特殊的植物区系组成而闻名,该地植被通常称为“半萨王纳植被”或“河谷型萨王纳植被”[11-12]。该地区植被发育较差且组成结构单一,多为禾本科植物,植被覆盖率低。元谋干热河谷地区森林覆盖率仅为5.2%,生态环境脆弱,生态系统退化严重,区域经济、社会发展与脆弱的生态环境之间的矛盾十分突出,该现状对区域生态安全及其可持续发展能力造成严重威胁[13]。目前,针对元谋干热河谷植物多样性的研究主要在幼苗更新[14]、人工繁育[15]、生态适应性[16]、植被恢复途径与方法[17]、现有植被演变过程[18]等方面,而对于该地区不同植被类型对生物多样性影响的研究较少。本研究主要依据云南元谋干热河谷的植被调查数据,以4种植被(草地、稀树灌草丛、人工混交林、人工纯林)为对象,开展了植物群落特征研究,运用物种多样性指数全面且客观地探究了该地区不同植被物种多样性水平,便于不同植被间比较及评价,同时,揭示了不同植被对生物多样性的影响,以期为合理保护与科学利用云南元谋干热河谷生态系统提供参考。
1 研究区概况
选择云南元谋干热河谷生态系统国家定位观测研究站为试验地,该站位于金沙江支流龙川江中下游[19],为典型南亚热带季风河谷干热气候[20]。该地为河谷冲积平原,四周地势高,海拔1 200~1 700 m,年均温21.9 ℃,年均降水量616 mm,年均蒸发量达3 627 mm[21],燥红土和红壤为其主要的地带性土壤。由于该地区降水少、蒸发量大、降水蒸发比严重失衡[22]、生态环境脆弱、人类活动干扰强烈,因此生态环境退化严重。
2 研究方法
2.1 群落调查
在元谋干热河谷地区,选择4种典型植被类型(草地、稀树灌草丛、人工混交林、人工纯林),每种植被类型设置3个重复样方,每个样方面积20 m×20 m,对各样方内所有地径(d)≥1 cm、离地20 cm的木本植物进行每木检尺。样方中心设置2 m×2 m的小样方,记录灌木层植物,在小样方的右下角设置1 m×1 m的小样方,记录地被层高度(h)≤0.5 cm以下木本植物和草本植物,记录所有植物的种名、数量、株高、胸径(或地径)、盖度、坐标等信息。

表1 研究样地不同植被类型基本概况
2.2 物种组成、重要值、多样性指数及相似系数计算
参考王伯荪[23]的方法计算各物种的重要值(Iv)、相对多度(Dr)、相对显著度(Pr)和相对频度(Fr)。
物种重要值Iv=(Dr+Pr+Fr)/3;
相对多度Dr=(D/∑D)×100%;
相对显著度Pr=(P/∑P)×100%;
相对频度Fr=(F/∑F)×100%;
BA=π×(DBH/2)2。
式中:D为某个种的株数;∑D为全部种的株数;P为某个种的断面积;∑P为全部种的总断面积;F为某个种的频度;∑F为全部种的总频度;π为圆周率;BA为胸高断面积;DBH为胸径。
选取较为广泛的多样性指数和相似系数来测算群落各层次物种多样性和群落相似性[24-26]。公式如下:
Shannon-Wiener指数H=-∑(PilnPi);
Margalef指数Dmg=S-1/lnN;
Pielou指数J=H/lnS;
Jaccard相似系数SIj=j/(A+B-j)。
式中:S为样方中物种的总数;N为所有物种个体的总数;Pi为第i个物种在样方中个体数占总数的比例;A为A类物种总数,B为B类物种总数,j为共有的物种数量。
2.3 数据处理
利用Excel 2019进行数据处理,采用单因素方差分析进行不同植被生物多样性指标的差异显著性检验(P<0.05),SPSS 23.0统计分析软件(IBM,USA)进行数据分析。
3 结果与分析
3.1 物种组成
研究区共有植物27科43属52种(表2)。含2种及以上的科有8个,共24属25种,分别占总科数的29.63%、总属数的55.81%、总种数的55.56%。植物种类较多的科分别为禾本科、菊科、蝶形花科等,其中,禾本科植物最多,共有5属5种,占群落中所有植物属的11.63%、所有种的11.11%。群落中单种科植物较多,占所有科的70.37%。乔木层植物共9种,隶属6科8属,主要由含羞草科、壳斗科、使君子科等组成,代表树种为滇榄仁(Terminaliafranchetii)、锥连栎(Quercusfranchetii)、铁橡栎(Quercuscocciferoides)、余甘子(Phyllanthusemblica)、新银合欢(Leucaenaleucocephala)、赤桉(Eucalyptuscamaldulensis)等;灌木层植物共17种,隶属13科16属,主要由蝶形花科、茜草科、蔷薇科等组成,华西小石积(Osteomelesschwerinae)、沙针(Osyrislanceolata)、车桑子(Dodonaeaviscosa)、西南杭子梢(Campylotropisdelavayi)等物种广泛分布;地被层植物共有17科26属26种,主要由禾本科、菊科、龙胆科等组成。

表2 研究区主要植物名录及其重要值
3.2 重要值分析
对于重要值[27],乔木层中,稀树灌草丛以滇榄仁为优势种,物种间个体数差异明显;人工混交林以滇榄仁、赤桉、新银合欢为主;人工纯林为单一树种人工纯林。对于灌木层,稀树灌草丛优势种为华西小石积、沙针;人工混交林主要优势种为车桑子、龙须藤(Phanerachampionii)、西南杭子梢,次要优势种为清香木(Pistaciaweinmanniifolia)、白刺花(Sophoradavidii)、薄皮木(Leptodermisoblonga);人工纯林的植物种类较单一,部分物种如车桑子重要值达到86.22%,与木豆(Cajanuscajan)的重要值(13.78%)差距显著,群落结构不稳定(表2)。
3.3 植被生物多样性分析
3.3.1 不同植被物种多样性分析
由表3可知,不同植被物种多样性变化趋势基本一致。单因素方差分析可以看出,Simpson优势度指数、Shannon-Wiener多样性指数、Pielou均匀度指数在不同植被间无显著差异,Margalef丰富度指数在不同植被间有显著差异(P<0.05)。
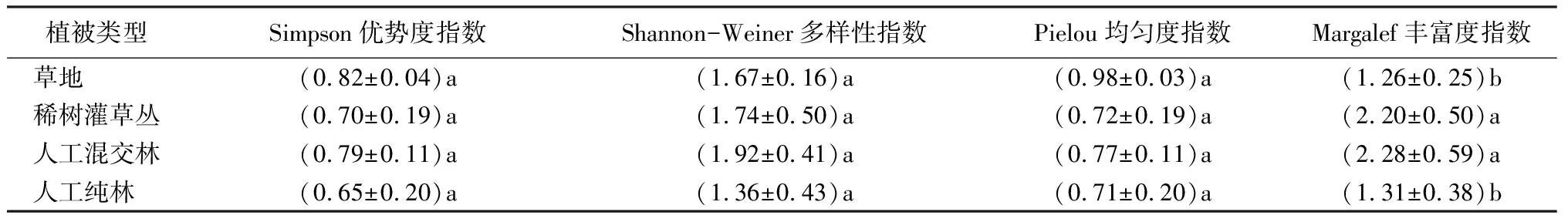
表3 不同植被物种多样性分析及差异性检验结果
不同植被类型的Simpson优势度指数从高到低依次为草地、人工混交林、稀树灌草丛、人工纯林,该现象表明,在草地的物种组成中,禾本科有显著优势地位。Shannon-Wiener多样性指数从高到低依次为人工混交林、稀树灌草丛、草地、人工纯林,这说明人工混交林具有更丰富的生物多样性,多个物种共存并发挥生态功能。Pielou均匀度指数从高到低依次为草地、人工混交林、稀树灌草丛、人工纯林,出现该现象的原因是草地在地被层中物种组成相对均匀。整体来看,人工混交林物种组成相对均匀,多个物种具有较高丰富度和重要值,无明显优势物种,易于维持群落稳定和功能。Margalef丰富度指数从高到低依次为人工混交林、稀树灌草丛、人工纯林、草地,其中,人工混交林的植物物种较多,适应性和群落稳定性均较好。
3.3.2 不同群落层次物种多样性分析
不同植被类型的群落层次物种多样性指数计算结果见表4。草地着重分析其地被层,人工纯林为单一优势种组成的纯林,故重点分析其灌木层和地被层。乔木层中,物种多样性指数均表现为人工混交林的最大,草地和人工纯林的最小,且各多样性指数差异均达到显著水平(P<0.05)。灌木层中,Simpson优势度、Shannon-Weiner多样性、Margalef丰富度指数最大的是稀树灌草丛,且稀树灌草丛的Margalef丰富度指数与其他植被类型相比有显著差异(P<0.05)。灌木层的Pielou均匀度指数由大到小依次为人工混交林、稀树灌草丛、人工纯林、草地。在地被层中,草地的Simpson优势度指数均显著大于其他植被类型(P<0.05)。除Simpson优势度指数,地被层的4个植被类型物种多样性指数差异均不显著。4种植被类型的乔灌层物种多样性指数间均存在显著差异(P<0.05)。人工混交林各群落层次的Pielou均匀度指数无显著差异。稀树灌草丛及人工纯林的乔木层Pielou均匀度指数显著低于灌木层及地被层的(P<0.05),这是由于稀树灌草丛及人工纯林乔木层的物种组成简单,单一优势种占比高,因此树种分布不均匀。

表4 不同植被类型的群落层次物种多样性分析及差异性检验结果
3.4 不同植被相似性分析
以Jaccard相似性指数计算不同植被类型在物种组成上的相似性程度。从表5可以看出,不同类型植被间物种组成存在较大差异,相似系数为0.16~0.31,相似性程度较低。具体来看,稀树灌草丛与人工混交林的相似系数最大,为0.31;草地与人工纯林的相似系数最小,为0.16,处于极不相似水平;其他植被与稀树灌草丛的相似系数为0.23~0.31,表明不同植被间相似性程度较低,处于中等不相似水平,在物种组成上存在较大差异。
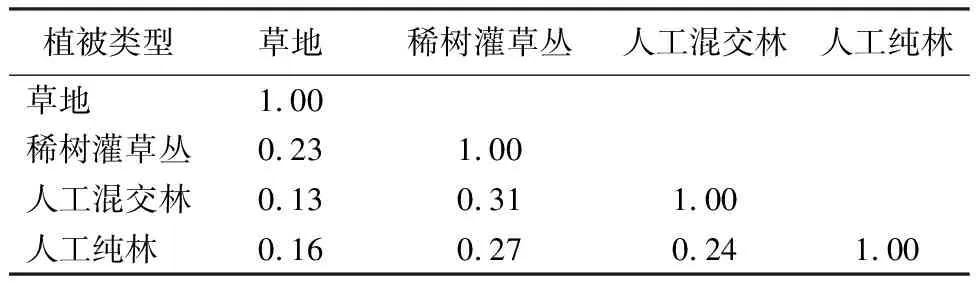
表5 不同植被类型的Jaccard相似系数
4 结论与讨论
植物群落的分布和组成能够体现群落特征[28],植物多样性可以优化生态系统功能[29-30],不同植被类型对植物群落有广泛影响[31]。本研究通过对元谋干热河谷不同植被生物多样性进行调查,对该地区不同植被物种组成和生物多样性特征进行对比。研究表明,元谋干热河谷地区植物群落主要以禾本科、菊科、蝶形花科、含羞草科、茜草科、龙胆科、蔷薇科、莎草科、壳斗科为主,具有典型的稀树灌草丛特征。地被层植物的科、属、种组成较乔木层及灌木层复杂。在4种不同植被类型的地被层中,重要值最大的科均为禾本科,扭黄茅、拟金茅、矛叶荩草(Arthraxonlanceolatus)、橘草(Cymbopogongoeringii)为其主要优势种。可以看出,禾本科在元谋干热河谷不同植被类型中均具有极强的竞争力,即使生境发生较大变化时,其优势地位仍然明显。通过对比物种组成和重要值可以观察到不同群落层次的优势科组成种类较为相似,而各科的重要值存在差异。乔木层、灌木层、地被层的物种重要值存在差异,且单种科、单种属占比均较高,证明植物演替有极大潜力。
优势种影响群落的结构且能反映群落生长状况和发展趋势,是群落重要的组成部分[32]。元谋干热河谷地区植物种类稀少、结构简单,是干热河谷中退化景观最为典型的地段之一[33]。该地区植被上层以单种乔木为优势种,混杂有余甘子和清香木等当地乡土植物,地被层以扭黄茅及拟金茅为主要优势种。在干热河谷中,禾本科占主导地位,其Simpson优势度指数为0.50~0.82。植被恢复过程中,过度的物种优势度会带来竞争加剧和生态系统功能下降的风险。因此,为维持生态系统及功能的稳定,应避免优势物种的过度扩张,保持适度优势度和其他物种存在,同时,可充分利用乡土与外来植物组合的方式,维持较高的物种多样性水平,促进物种共存和物种间的相互依赖[34-35]。
在进行生物多样性指数分析时,需要将物种组成与多样性状况相结合,以便客观评价群落多样性状况[36-37]及定量观察群落或生境中物种丰富度及均匀度的变化程度[38-39]。以重要值来测度群落多样性有良好的准确性,有助于深入了解群落结构和功能[40]。物种丰富度和多样性是衡量群落稳定性的重要指标,物种多样性越高,生态系统抗干扰能力越强,植物群落则越稳定[41]。
本研究中,4种不同植被类型物种丰富度由大到小依次为人工混交林、稀树灌草丛、人工纯林、草地。4种植被类型的Shannon-Weiner多样性指数为1.67~1.90,其指数已接近荒漠群落植物Shannon-Weiner多样性指数[42],这表明,元谋干热河谷环境恶劣,物种多样性较低。4种植被类型的乔木层及灌木层物种Shannon-Weiner多样性指数均存在不同程度的差异。稀树灌草丛和人工纯林中,乔木层物种Shannon-Weiner多样性指数显著低于灌木层及地被层,主要原因是乔木层物种组成简单,单一优势种占比高。稀树灌草丛与人工混交林间的Jaccard指数最大,两者之间植物物种最相似,群落共有种较多。人工纯林通常存在集中分布大量单一树种的问题,因此导致此植物类型的物种多样性相对较低,当生境发生变化,既出现不适宜该群落优势树种生存条件时,整个群落稳定性会受到严重破坏。相比而言,稀树灌草丛及人工混交林具有更丰富的物种组成和更广泛的生态适应性。为提高植被稳定性并促进整个生态系统的健康发展,建议采用乔木与地被层或灌木与地被层并行的恢复模式,增加不同物种的丰富度和多样性[26]。
元谋干热河谷植被生物多样性指数普遍较低,不同植被类型对生物多样性的影响有一定差异。整体来看,禾本科在物种组成中显著占优。人工混交林具有高物种多样性、均匀的物种丰度分布、潜在的生态系统稳定性的特点。草地及人工混交林整体表现为物种组成相对均匀,而相对均匀的物种组成则能够维持群落及其功能的稳定,其中,人工混交林的植物物种较多,展现出更好的适应性和群落稳定性。今后,干热河谷需发展具备耐旱、耐热、适应性强等特征的植被,并在树种选择时综合考虑当地气候、土壤条件、水资源状况、物种与生态系统的相互关系,减少单一物种的影响,增加整个群落对干热等干扰的抵抗力,进而提高群落的稳定性和适应性,使群落能更好地应对环境变化及压力。通过人工造林等植被恢复措施,可丰富干热河谷地区的物种组成,进而形成多树种、多层次、多功能、多效益的植被恢复体系。
猜你喜欢
中老年保健(2022年7期)2022-09-20
今日中国·西班牙文版(2021年4期)2021-04-09
民族音乐(2019年2期)2019-12-10
西藏农业科技(2019年1期)2019-07-25
森林工程(2018年4期)2018-08-04
现代园艺(2018年2期)2018-03-15
方圆(2018年23期)2018-01-07
民族音乐(2016年2期)2016-04-04
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
