
Biocontrol potential of entomopathogenic nematode, Heterorhabditis indica against pink bollworm, Pectinophora gossypiella (Saunders)(Lepidoptera: Gelechiidae)
2024-01-13 10:39:52THUBEShivajiSHINDESwatiSHAHVivekGOKTENARKHEDKARNandiniINGOLEDnyaneshwarNIKOSHEAkashTENGURIPrabhulingaTHAVKARSnehalFANDBabasahebDESHMUKHVrushaliandPRASADYenumula
Journal of Cotton Research 2023年4期
THUBE Shivaji , SHINDE Swati, SHAH Vivek, GOKTE-NARKHEDKAR Nandini, INGOLE Dnyaneshwar,NIKOSHE Akash, TENGURI Prabhulinga, THAVKAR Snehal, FAND Babasaheb, DESHMUKH Vrushali and PRASAD Yenumula
Abstract Background The emergence of pink bollworm (PBW), Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae), in cotton due to Bt resistance and concealed feeding habit has created a need for alternative, eco-friendly,and cost-effective control methods.This study aimed to evaluate the bio-efficacy and reproductive potential of two native strains of entomopathogenic nematodes (EPNs), Heterorhabditis indica, namely CICR-HI-CL and CICR-HI-MN,against PBW larvae and pupae under in-vitro conditions.
Keywords Bio-efficacy, Entomopathogenic nematodes, Multiplication rate, Heterorhabditis indica, Pectinophora gossypiella
Background
Cotton is the major fiber crop grown worldwide.India leads the production of cotton (36.5 million bales i.e., 6 205 000 tons, 1 bale = 170 kg) among the different cotton-growing countries.Cotton plays a key role in India’s economy, providing employment directly and indirectly to more than 60 million people (Cotton Corporation of India STAT 2022).India is the only country where all four cultivated species of cotton (Gossypium hirsutum,G.barbadense,G.arboreum, andG.herbaceum) are grown.Although India ranks first in total area coverage, cotton productivity is the lowest (464 kg lint per hectare) among major cotton producers of the globe.Several biotic and abiotic factors are responsible for the low productivity of cotton.Among the biotic factors, the incidence of insect pests is reported as a key limiting factor for cotton production (Nagrare et al.2022).
Among various insect pests, pink bollworm (PBW),Pectinophora gossypiellais a major threat with a record yield reduction potential up to the tune of 90%, before the use of broad-spectrum insecticides and introduction of transgenic cotton (Patil 2003).The introduction of synthetic pyrethroids in India during the 1980s marked a major milestone and played a strategic role in managing this notorious pest of cotton.However, the indiscriminate use of these chemical insecticides caused widespread ecological disruption leading to the aggravation of PBW and other secondary pest problems in the cotton ecosystem (Kranthi et al.2002; Kranthi and Russell 2009; International Cotton Advisory Committee 2010).
Subsequently, the introduction of genetically engineered transgenic cotton (Bt cotton) carrying genes encoding delta-endotoxin proteins (Bollgard I and Bollgard II) from the entomopathogenic bacteriumBacillus thuringiensisBerliner was yet another key milestone for the effective management of PBW in cotton.Initially, Bt cotton performed well and offered promising control of bollworm complex until 2009.However, later on, reports emerged on the resistance of PBW populations to Cry1Ac and their survival on Bollgard I cotton(Dhurua and Gujar 2011).The infestation of PBW on Bt cotton decreased in 2010 and 2011, but since 2014,it has become more widespread and is observed in all cotton fields, including Bollgard II.The development of evolved resistance, primarily against transgenic cotton(Bollgard I and Bollgard II), is the main reason behind the widespread infestation of PBW in Indian cotton(Fand et al.2019).Worldwide, PBW has been known to cause 2.8%–61.9% loss in seed cotton (Bhute et al.2023).Recently India witnessed the infestation of PBW ranging from 8%–92% with yield loss between 10%–30 % (Nagrare et al.2023).
In order to safeguard the cotton crop from PBW infestation, cotton growers resort to periodical, indiscriminate use of broad-spectrum insecticides.Besides affecting human health, these insecticides are also responsible for environmental pollution in the fragile agricultural ecosystem, incurring enormous setbacks to natural resources(Thube et al.2022).Until now, pesticide poisoning has claimed the lives of several farm laborers engaged in cotton fields.In India, the accidental inhalation of insecticides has caused 21 deaths in three months during 2017(The Hindu 2017).Acute pesticide poisoning is also recorded from the male and female cotton growers of India (Mancini et al.2005).In addition to the mortality of natural enemies of pests, insecticides are responsible for the pollution of natural resources and upsetting the beneficial soil microflora.The neonicotinoid group of insecticides is detrimental to pollinators and its residues are detected in stamens and maternal flowers of cotton in China (Wu et al.2022).The residues of various pesticides are detected in cotton seeds which is an important source of edible oil (Blossom and Singh 2004).
Entomopathogenic nematodes (EPNs) are considered as one of the potential biological control agents for managing insect pests in a safe and eco-friendly manner.EPNs can effectively control a wide range of soil-dwelling insect pests and are known for their selfperpetuating nature, species-specificity, and ease of mass multiplication.In contrast to chemical pesticides,EPNs do not leave any toxic residue in the environment and are considered to be a sustainable alternative for managing insect pests in agriculture.EPNs have been extensively studied and found to be effective against a wide range of soil-inhabiting insects (Begley 1990; Klein 1990).The two main families of EPNs,Steinernematidae and Heterorhabditidae, are obligate insect parasites (Poinar 1979) that carry associated bacterial symbionts (Akhurst and Boemare 1990).The infective juvenile (IJ) stage of the nematode remains in the soil until it can invade the body of a susceptible insect.After infection, the symbiotic bacteria are released into the insect hemocoel, causing septicemia and death of the insect (Kaya and Gaugler 1993).EPNs feed on the bacterial cells inside host tissues, and once the host reserves are depleted, a new generation of IJs exits the cadaver (Kung et al.1991).EPNs are widely used in insect pest management, particularly for pests that have a soil-dwelling stage in their life cycle (Kaya and Gaugler 1993).Heterorhabditishas been extensively studied for managing lepidopteran pests such asSpodoptera frugiperda(Patil et al.2022),S.litura(Gokte-Narkhedkar et al.2019), andS.littoralis(Sobhy et al.2020).Steinernema riobraveandHeterorhabditis bacteriophorahave been found to be highly pathogenic to PBW larvae (Gouge et al.1999; Shairra et al.2016).
Therefore, in the present study, we tested the virulence of two native strains ofHeterorhabditis indicaviz., CICRHI-CL and CICR-HI-MN (Shinde et al.2022), originally isolated from cotton fields, in areas where the PBW is one of the major pests.The reproductive potential of any biological control agent is marked as an important feature for their extended persistence and pathogenicity to the targeted insect pests (Blanco-Pérez et al.2017).Therefore, the reproductive potential ofH.indicawas also assessed on PBW under present investigations.
Results
Larval and pupal susceptibility of P. gossypiella to H. indica
The results indicate that both strains ofH.indicacan infect the larvae of PBW under laboratory conditions.However, the susceptibility of larvae to both strains varied significantly.The strain CICR-HI-CL was more virulent compared with CICR-HI-MN, causing higher mortality across all the larval instars evaluated in the present study.Bioassay results showed that percentage mortality increased significantly (For strain CICR-HI-MN:F8,26= 23.04;P< 0.000 1 and CICR-HI-CL:F8,26= 31.85;P< 0.000 1), with an increase in IJs dosages (Fig.1).Probit analysis of strain CICR-HI-CL indicated that theLD50values for 2nd, 3rd, and 4thinstar larvae were 8.75,6.57, and 7.14 IJs per larva, respectively.However, strain CICR-HI-MN was comparatively less virulent against 2nd, 3rd, and 4thinstar larvae, withLD50values of 21.06,24.81, and 30.22 IJs per larva, respectively.When comparing the susceptibility of various larval instars within the strain, it was observed that 2ndinstar larvae exhibited a higher susceptibility to strain CICR-HI-MN, whereas 3rdinstar larvae were more susceptible to strain CICRHI-CL (Table 1).When IJs were inoculated into the pupae of PBW using the direct application method, strain CICR-HI-CL exhibited greater virulence with a lowerLD50value (29.65 IJs per pupa) compared with strain CICR-HI-MN (73.88 IJs per pupa).One-way analysis of variance revealed significant differences in pupal mortality across the tested IJ doses for both strains (For strain CICR-HI-CL:F10,54= 154.34,P< 0.000 1; for CICR-HIMN:F10,54= 225.42,P< 0.000 1).However, the application of IJs through the soil resulted in reduced pupal mortality, indicated by higherLD50values (For strain CICR-HICL: 147.84 IJs per pupa; for strain CICR-HI-MN: 272.38 IJs per pupa).TheLD50values for various dosages for both strains are presented in Table 2.
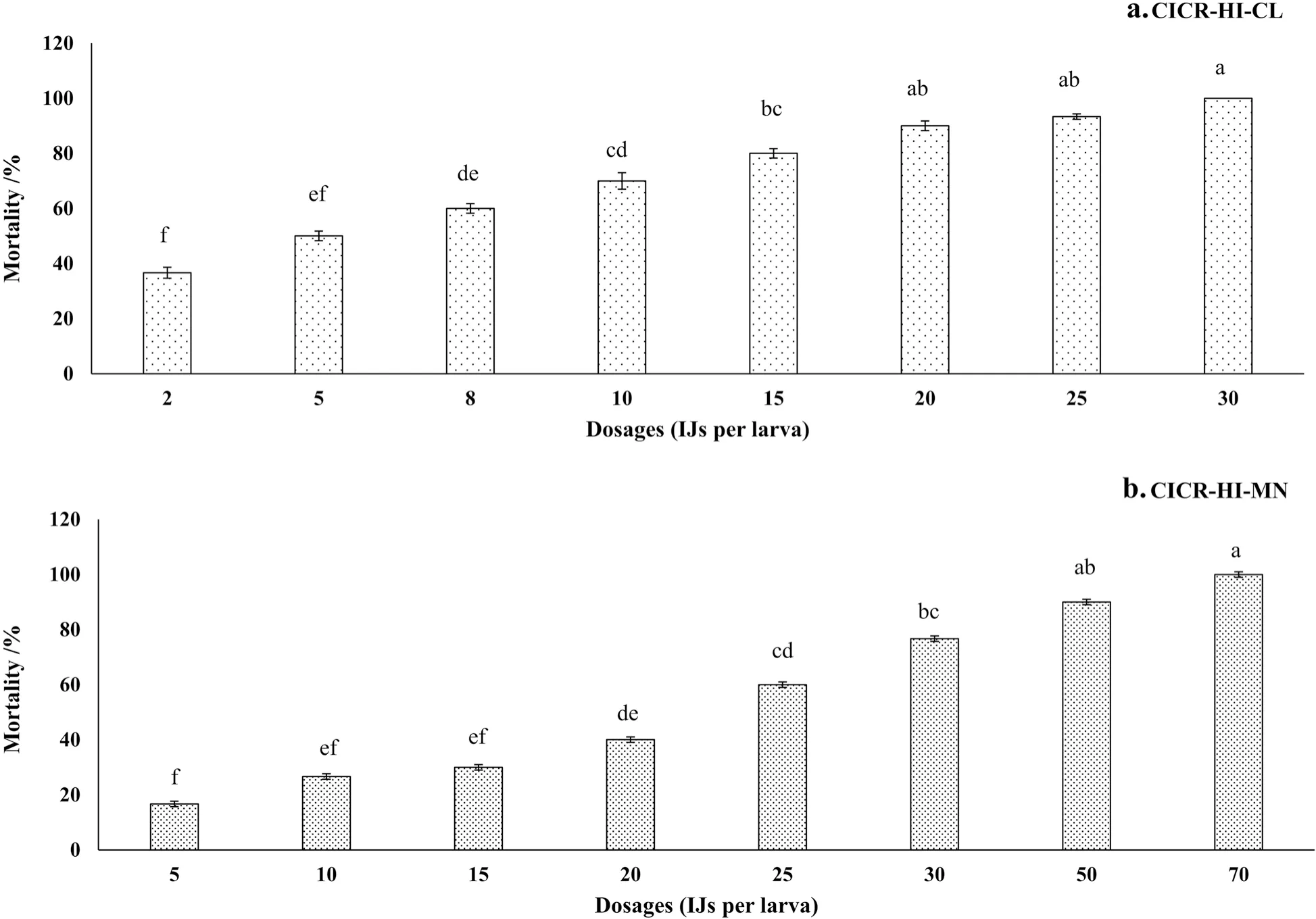
Fig.1 Percent mortality (mean ± standard error) of third instar larvae of pink bollworm, P. gossypiella, at different dosages of H. indica (a CICR-HI-CL strain; b CICR-HI-MN strain) at 72 h after treatment in laboratory conditions
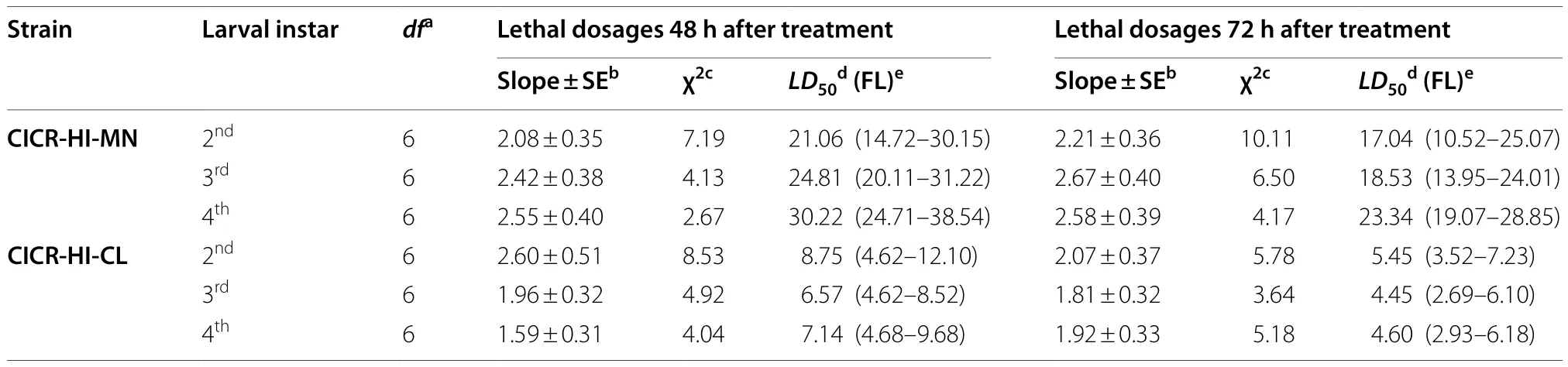
Table 1 Dose–mortality response (expressed as lethal dosages) of second, third and fourth instar larvae of Pectinophora gossypiella to Heterorhabditis indica under laboratory conditions
The PBW larvae turned brick red after 48 h of inoculation, indicating that the larval mortality was due toH.indicainfection (Fig.2b).The initial depletion of internal content of larvae resulted in a change of body color from brick red to yellowish-brown at 96 h of plating (Fig.2c).Gradually, EPN multiplication continued in the larval cadavers, leading to the complete depletion of internal content.As a result, the cadaver became translucent, and the mass of EPNs was clearly visible in the larval hemocoel under a stereo zoom-microscope (Fig.2d).However, the pupal stage invaded byH.indicacould not be differentiated by the appearance of brick red color due to the presence of a higher titre of melanin.Heterorhabditis indicainfected pupae initially turn darkish brown to black and turn translucent with a visible mass of EPNs within the body after 6–7 days of inoculation (Fig.2e).
Reproductive potential
The results of the reproductive assay revealed that both strains ofH.indicawere able to penetrate and reproduce within the hemocoel of fourth-instar larvae ofP.gossypiella.A significant difference was observed in the reproductive potential of both strains across different dosages, CICR-HI-MN (ANOVA:F= 9.83;df= 10;P< 0.005) and CICR-HI-CL (F= 5.36;df= 10;P< 0.05).However, the reproductive potential of strainCICR-HI-MN was significantly higher than that of strain CICR-HI-CL when inoculated with lower dosages (10 and 20 IJs per larva) (Fig.3).The next generation of IJs emerged nine days after inoculation and continued for fourteen days in strain CICR-HI-CL.However, in strain CICR-HI-MN, IJs emerged ten days after inoculation and continued for twelve days.A proportionate increase in the reproductive potential of both strains was observed up to the dose of 30 IJs per larva.However, a reduction in the emergence of IJs was observed for the higher inoculated dosages (40 and 50 IJs per larva).The dose of 30 IJs per larva was found to be optimal for the production of the highest number of next-generation IJs for both strains.

Table 2 Dose–mortality response (expressed as lethal dosages (LD) of Pectinophora gossypiella pupae to Heterorhabditis indica using direct and soil application methods under laboratory conditions

Fig.2 Infection of H. indica to P. gossypiella a Freshly inoculated larvae; b Brick red cadaver after 48 h of inoculation; c Yellowish brown larval cadaver after 72 h of inoculation; d Translucent larval cadaver with a visible mass of EPNs in larval hemocoel; e Translucent pupal cadaver harbouring H.indica within the body
IJs fitness assay
The IJs of both strains ofH.indicaemerged successfully from larval cadavers and demonstrated the ability to infect and kill the fresh 4thinstar larvae of the PBW.No mortality was observed in the control.The bioassay findings indicated a significant increase in percent mortality with an increase in dosages of IJs for both the strains.When IJs were inoculated into the larvae at higher doses,specifically 30, 40, and 50 IJs per larvae, the recorded mortality was 100% even after 48 h of inoculation for strain CICR-HI-CL.Additionally, the analysis of the IJs fitness assay data revealed that the strain CICR-HI-CL exhibited significantly higher virulence compared to strain CICR-HI-MN at both 48 h (F= 29.82,df= 11, 36,P< 0.001) and 72 h of inoculation (F= 54.97,df= 11, 36,P< 0.000 1).
Discussion
The effectiveness of using EPNs in integrated pest management has been well established worldwide (Kumar et al.2022).The use of local EPN strains is crucial for achieving efficient pest control since they are better suited to the local environment (Nikdel and Niknam 2015).In vitro evaluation remains a significant step in identifying the most virulent strain of EPNs that can be further deployed for field application.Reports on the biocontrol potential of EPNs against PBW are meager.As of now, only two species of the generaHeterorhabditishave been found to be effective against PBW, namelyH.indica(Shahina et al.2014) andH.bacteriophora(Gouge et al.1999; Shairra et al.2016).The present study investigated the biocontrol potential of two native strains ofH.indicaagainst PBW.Our findings revealed that both strains varied significantly concerning their virulence against larval and pupal stages of PBW.Nevertheless, both strains were lethal to PBW, and the strain CICR-HI-CL, which recorded lowerLD50values, was found to be more effective than the strain CICR-HI-MN.The virulence of any EPN species is mostly related to numerous factors, including the host insects, multiplication, and penetration (Kaya& Gaugler 1993).Both strains ofH.indicaemployed in the present study were isolated from a maize field rotated with cotton crops located at Nagpur, India(21°04′48.39″N 78°06′58.02″E).
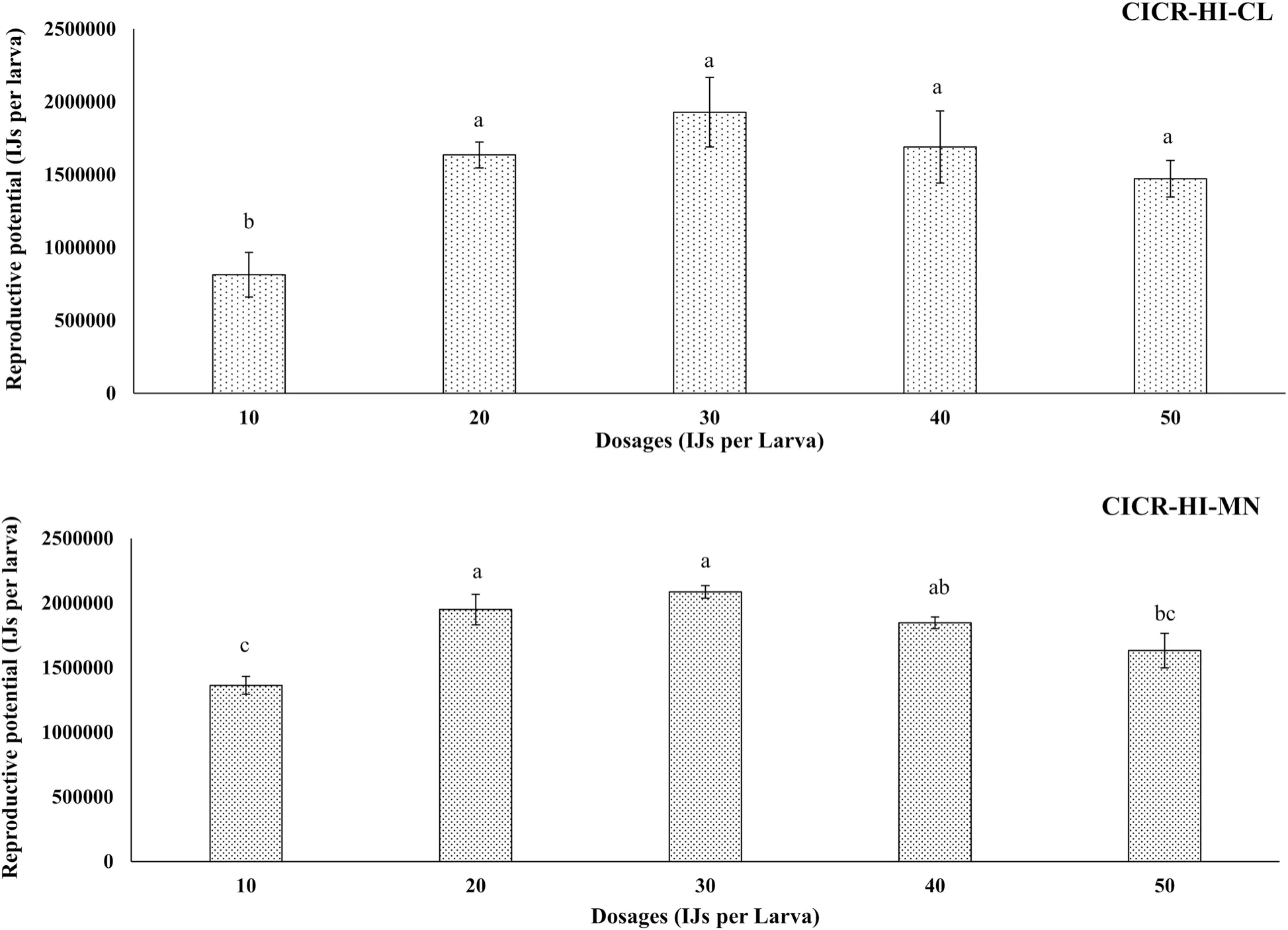
Fig.3 Reproductive potential of H. indica strains (CICR-HI-CL and CICR-HI-MN) on 4th instar larvae of P. gossypiella.Error bars indicate the standard error of mean (SEM).Different letters on the bars indicate statistically significant difference across dosages within the H.indica strain using WASP software in a completely randomized design at 0.05 level of significance
TheLD50values obtained after treating EPN strains,namely CICR-HI-CL and CICR-HI-MN, against 3rdinstar larvae of PBW at 48 h were 6.57 and 24.81 IJs per larva, respectively.However, in our previous study, the same strains demonstrated relatively higherLD50values of 21.65 and 48.91 IJs per larva at 72 h after treating fall armywormSpodoptera frugiperda(Shinde et al.2022).The substantially lowerLD50values in PBW could be attributed to the high virulence and preference of these EPN strains against PBW compared withS.frugiperda.The differences in biological attributes of larvae, such as size, weight, cuticular composition, and hemocoel, could be probable reasons for the rapid mortality of PBW compared toS.frugiperda.
The success of EPNs depends on the target life stage of the insect pest.EPNs generally prefer to feed and multiply on the larval stages of insect pests because of their sluggish movement, the presence of more natural openings like spiracles, and the provision of hemocoel as a medium for bacterial multiplication.Several studies have shown that the larval stages of lepidopteran pests are susceptible to infection by different EPNs (Salari et al.2021; Aryal et al.2022).Our results on the virulence ofH.indicato larval stages of PBW are consistent with earlier studies conducted against other lepidopteran pests (Kumar et al.2022; Shinde et al.2022; Patil et al.2022).The high virulence of strain CICR-HI-CL against 3rdinstar larvae of PBW reported in the present study is in agreement with that of earlier researchers reported againstS.frugiperda(Shinde et al.2022).
Several studies have shown that pupal stages are more resistant to EPN infection than larval stages (Hassani-Kakhki et al.2013; Kary et al.2018; Yan et al.2020; Mhatre et al.2020; Aryal et al.2022).In agreement with this,the present study indicates thatH.indicais less virulent against pupae (LD50values: CICR-HI-CL = 29.65; CICRHI-MN = 73.88) ofP.gossypiellathan against larvae (LD50values: CICR-HI-CL = 6.57; CICR-HI-MN = 24.81 IJs per larva).Our findings of higher larval mortalities and lower pupal mortalities instigated byH.indicaare in agreement with other reports, namely Henneberry et al.1995, 1996(P.gossypiella), Batalla-Carrera et al.2010 (Tuta absulata), Kary et al.2019 (Plutella xylostella).
The virulence of both strains is further reduced when applied through soil rather than by direct application(LD50values: 147.84 and 272.38 IJs per larva).This reduction in virulence could be due to the inability of all the inoculated IJs to locate pupae in the soil.Lower virulence of both strains against the pupal stage is also attributed to the formation of a protective puparium, closure of anal and oral apertures, high melanization and presence of compact hemocoel which may be more difficult for IJs to penetrate (Aryal et al.2022).Based on the present study,it can be concluded that, both the strains evaluated under present investigations are virulent against the larval and pupal stages of PBW.The 4thinstar larvae supports the high reproduction of both the strains under laboratory conditions which may increase the persistence of IJs under field conditions.
Apart from assessing the suitability of a target host insect for multiplication, the reproductive potential of EPN is also considered an important indicator of its persistence under field conditions (Blanco-Pérez et al.2017).Earlier reports on the susceptibility of PBW toH.indicadid not address the reproductive potential of this EPN,we address this issue in the present study.The reproductive potential of EPNs is proportional to its virulence against the target pest, ensuring success in perpetuation under field conditions (Patil et al.2022).However, in contrast to this, our present study reported that the strain CICR-HI-CL is more virulent, but with less reproductive potential than strain CICR-HI-MN.Our results on the reproductive potential of these strains are in agreement with the report of Shinde et al.(2022).These differences in the production of the next generation of IJs might be associated with the variation in aggressiveness, size, and/or number of penetrated IJs, mating behavior, production of oocytes and/or sperm, etc.A successful invasion of EPNs into the insect hosts and the emergence of IJs from insect cadavers is essential for the effective control of insect pests, especially pests like PBW, whose larval stages feed on developing bolls and pupate in soil.The IJs fitness assay clearly demonstrated the successful emergence of IJs from larval cadavers and their subsequent ability to infect and kill fresh 4thinstar larvae of the PBW.The absence of mortality in the control group validates the role of IJs in causing the observed mortality.The bioassay findings revealed a dose-dependent increase in percent mortality with higher dosages of IJs for both strains,underlining the importance of dosage in enhancing effi-cacy.One of the notable findings was the achievement of 100% mortality in the PBW larvae when inoculated with higher IJ doses (30, 40, and 50 IJs per larva) for strain CICR-HI-CL.This high mortality rate, even after 48 h of inoculation, highlights the potent virulence of this particular strain in effectively controlling PBW larvae.Moreover, the comparison of IJs fitness assay data between the two strains indicated that strain CICR-HICL exhibited significantly higher virulence than strain CICR-HI-MN at both 48 and 72 h post-inoculation.This difference in virulence suggests potential variations in the genetic makeup or adaptability of the strains, possibly contributing to their distinct effectiveness against PBW larvae.These findings have significant implications for pest management strategies.The identification of highly virulent strains, such as CICR-HI-CL, could inform the development of more effective biological control measures against PBW infestations, potentially reducing reliance on chemical insecticides and promoting sustainable agricultural practices.Further research and investigation into the mechanisms underlying the enhanced virulence of specific strains are essential for harnessing their full potential in integrated pest management programs.This study indicates thatH.indicais an effective biological control agent of PBW, and the high reproduction rates of this species may increase the persistence of IJs under field applications.
Conclusion
The present study has generated useful information on virulence, reproductive potential and fitness of emerged IJs of two strains of EPNH.indica(CICR-HI-CL and CICR-HI-MN) against PBW, a dreaded pest of cotton.Higher virulence and reproductive potential of EPN strains demonstrated their ability to multiply, sustain and perpetuate on larval and pupal stages of PBW.The knowledge generated will help formulate effective management strategy for PBW with the inclusion of EPNs as potential biological control candidates.The soil dwelling life stages viz., last instar hibernating larvae and pupae of PBW can be the ideal weak links to make a successful use ofH.indicafor sustainable management of PBW in the cotton ecosystem.However, before taking these EPN strains to the field for managing PBW, detailed studies investigating their biocontrol potential against PBW under field conditions are needed.
Material and methods
Insect culture
The larvae of PBW were collected from 110-day-old cotton crop (Gossypium hirsutumL.) variety Suraj(non-Bt) grown in the experimental farm of the Indian Council of Agricultural Research-Central Institute for Cotton Research (ICAR-CICR), Nagpur during 2020–2021.The field-collected larvae were reared on an artificial diet under controlled conditions in the insectary of ICAR-CICR, Nagpur at a temperature (27 °C ± 1 °C),relative humidity (65% ± 5%), and a 14 h light/10 h dark photoperiod (Shah et al.2021).A cotton twig containing squares was used as an oviposition substrate for PBW moths, and moths were provided with a plug of absorbent cotton dipped in honey solution (volume fraction of 10% prepared in distilled water) every alternate day as a moisture and energy source.Moths were allowed to pair and lay eggs on cotton twigs, which were dipped at the base in Eppendorf tubes filled with water and covered with Parafilm to keep them fresh for a longer time (Fand et al.2020).Twigs loaded with eggs were collected on tissue paper placed at the bottom of plastic jars to allow for clear visibility of neonate larvae.The newly hatched larvae were further reared on an artificial diet (Shah et al.2021).
Source of EPN
Two strains of EPN,Heterorhabditis indica, namely CICR-HI-CL and CICR-HI-MN, were procured from the EPN repository maintained at the Division of Crop Protection, ICAR-CICR, Nagpur, India.The nematodes were cultured on the last instars of the greater wax moth,Galleria mellonella, at (23 °C ± 2 °C).The infective juveniles that emerged from the larval cadavers ofG.mellonellawere collected using White traps (White 1927)and stored in sterilized water at 15 °C (Patil et al.2020).Three-day-old IJs were used in further experiments.
Laboratory bioassays
Larval bioassay
The virulence ofH.indicawas studied against 2nd, 3rd,and 4thinstar larvae ofP.gossypiella.The optimum quantity of water (diluent) required to treat a single larva of 2nd, 3rd, and 4thinstar was calibrated as 10, 20,and 30 μL, respectively.
Initially, a wide dose bioassay was carried out using dosages 2, 20, 100, 300 and 500 IJs per larva (data not shown here) to determine the final dosages of IJs to be used in bioassays against PBW larvae.Subsequently, the doses were fine-tuned with a total of eight dosages, i.e.,2, 5, 8, 10, 15, 20, 25, 30 IJs per larva for strain CICR-HICL and 5, 10, 15, 20, 25, 30, 50, 70 IJs per larva for strain CICR-HI-MN.Application of water only without IJs served as a control.
Ten larvae of each instar were placed separately in 24-well multi-cell plates (Genaxy Scientific Pvt.Ltd,India) and provided with cubes of semi-synthetic diet(0.25 to 0.40 mL) per well as a source of food (Shinde et al.2022).Each treatment (single dose) consisted of 50 larvae (n= 10 larvae per replication; five replications per treatment).For each strain, a total of 450 larvae were used to evaluate nine dosages including control.The desired dose of EPN was directly applied to the larval instar placed in multi-cell plates (Shinde et al.2022).All the plates were sealed with Parafilm to avoid moisture loss and incubated at (25 °C ± 2 °C) in a BOD incubator.Untreated control plates were maintained similarly to the treatments, except that no IJs were added to the water.Larval mortality was observed at 48 and 72 h after IJs inoculation.To confirm that larval mortality was due to nematode infection, the cadavers were dissected in Ringer’s solution under a stereomicroscope to verify the presence of nematodes.The whole experiment was repeated once with a fresh batch of IJs.
Pupal bioassay
The virulence of both strains of EPN was evaluated against 48-h-old pupae of PBW in two separate experiments.A total of ten dosages of IJs, i.e., 5, 10, 15, 20, 25,30, 40, 50, 75, 100 IJs per pupa for strain CICR-HI-CL and 5, 15, 25, 50, 75, 100, 125, 150, 175, 200 IJs per pupa for strain CICR-HI-MN, were used.
In the first experiment, the desired dosages of EPN suspension (30 μL) were directly applied to the pupae placed in multi-cell plates (Innovative Biosciences Private Limited, Nagpur, India).All the plates were sealed with parafilm to prevent moisture loss and incubated at(25 °C ± 2 °C) in a biological oxygen demand (BOD) incubator.Each treatment consisted of 50 pupae (n= 10 pupae per replication; five replications per treatment).A separate batch of pupae was maintained with distilled water without IJs for use as a control.Pupal mortality and adult emergence were recorded daily for ten days.In the second experiment, the desired dosages (50, 100, 200, 300,400, and 500 IJs per pupa) of EPN suspension (5 ml) were evenly applied to a plastic container (diameter: 12.0 cm;height: 5.0 cm) containing a mixture of sterilized sand and soil (2:1 ratio by volume).Control treatments were provided with distilled water without IJs.Ten pupae were placed at a depth of 2 cm in each container.The containers were closed with lids fitted with muslin cloth and incubated (25 °C ± 2 °C).Each treatment had 5 replicates(n= 10 pupae per replication; 50 pupae per treatment),and the assay was repeated once.Observations on the emergence of adults were recorded daily for ten days.The pupae from which adults did not emerge in both sets of experiments described above were collected, washed in distilled water, and dissected in Ringer’s solution under a stereomicroscope to verify the presence of nematodes.The adults that emerged from EPN-treated pupae were examined under a stereomicroscope for any deformities,if present.The whole experiment was repeated once with different batches of fresh EPNs.
Reproductive potential of H.indica strains on PBW
A separate assay was conducted by inoculating pure culture of both strains on fresh larvae of PBW.Reproductive potential ofH.indicastrains was studied as per the protocol outlined in Mhatre et al.(2020) and Patil et al.(2022).Five dosages (10, 20, 30, 40, and 50 IJs per pupa)were selected to assess the reproductive potential ofH.indicastrains on PBW.The nematode inoculation procedure was followed as mentioned in the larval bioassay.The 4thinstar larvae are full-grown, hibernating and soil-inhabiting larval stage of PBW (Gouge et al.1999).Hence, nematode-infected fresh cadavers of uniformsized 4thinstar larvae (Mean weight = 36.8 mg ± 0.45 mg per larva) were used in the assay.A total of 12 cadavers were selected randomly and washed with distilled water to remove nematodes adhering to the larval body.These cadavers were then transferred separately to the white trap (White 1927) and incubated at 23 °C ± 2 °C in the dark in a BOD incubator (Mhatre et al.2020).Observations on the time required for the emergence of the next generation of IJs, the total IJs emerged per larva, and the duration of emergence (the duration between the initial and last emergence of IJs) were recorded during the assay.Each cadaver was considered as a replicate; the whole experiment was repeated once, and the results of the reproduction assay were expressed as the number of IJs per larva.
IJs fitness assay
The experiment was performed using a Petri dish assay,following Kaya and Stock (1997).IJs derived from the previous experiment were used to study the fitness potential of emerged IJs from the dead cadaver of PBW to re-infect fresh larvae.Different dosages of IJs (10, 20,30, 40, 50 and 60 IJs per larva) were selected, and healthy larvae (4thinstars) of PBW were used for this assay.A Petri dish with a diameter of 150 mm was lined with double-layered Whatman filter paper No.1.Subsequently,0.5 ml of distilled water containing each concentration of nematodes was introduced into an individual Petri dish and incubated for 30 min.A single larva was then introduced into each Petri plate pre-inoculated with IJs.Six treatments, including a control, were set for each strain,with 5 replicates for each treatment.Each treatment had 4 replicates (n= 8 larvae per replication; 50 pupae per treatment).A Petri dish lined with filter paper wetted only with distilled water was used for each dose of EPN to serve as a control plate.
Statistical analysis
Experimental data, recorded on larval and pupal mortality, were subjected to probit analysis (Finney 1971) using Polo Plus software (Version 2.0, LeOra) to calculate LD50value.Fiducial limits were taken at a 95% confidence interval.The data on the mortality of PBW under laboratory bioassay, IJs fitness assay and mean reproduction potential ofH.indicawere subjected to one-factor analysis of variance (ANOVA) using SPSS software (version 22.0; SPSS, Chicago, IL, USA).When ANOVA was significant, comparisons of relevant means were made using Tukey’s significance test at the 5% level of significance.
Acknowledgements
The authors are thankful to the Indian Council of Agricultural Research, New Delhi for the grant in aid through ICAR-Central Institute for Cotton Research,Nagpur Institutional Project.Technical help received from the Technicians in carrying out experiments is duly acknowledged.The senior author extends their gratitude towards Mrs.Rohini Dhotre for her aid during the laboratory bioassays.Authors extend their sincere thanks to GT, Behere and Satish Chavan for their diligent efforts in refining and polishing the English of the manuscript.
Authors’ contributions
Thube S, Shah V., Gokte-Narkhedkar N., planned the work, designed the experimental setup, supervised the conduct of experiments, analyzed and interpreted the data, drafted the manuscript; Shinde S, Ingole D, Thube S,Thavkar S, Fand B, Prabhulinga T., and Nikoshe A, conducted laboratory experiments on bioefficacy evaluation, recorded the data, helped in data analysis and manuscript editing; Deshmukh V., planned, conducted IJs fitness assay and analysed data and Prasad YG approved the work plan, provided the research facilities/ infrastructure and monitored the work progress.All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Availability of data and materials
All the data relevant to the present study are included in the article.Any further details related to the experiments conducted can be made available by requesting the corresponding author.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
The manuscript has not been published, or submitted for publication elsewhere.
Competing interests
The authors declare that they have no conflict of interest related to the content of this article.
Author details
1Indian Council of Agricultural Research-Central Institute for Cotton Research,Nagpur, Maharashtra 440010, India.2College of Agriculture, Nagpur, Maharashtra 440001, India.3Dr.Balasaheb Sawant Konkan Krishi Vidyapeeth, Dapoli,Maharashtra 415712, India.
Received: 10 July 2023 Accepted: 4 November 2023
 Journal of Cotton Research2023年4期
Journal of Cotton Research2023年4期
- Journal of Cotton Research的其它文章
- Enhancing waterlogging tolerance in cotton through agronomic practices
- Mepiquat chloride priming confers the ability of cotton seed to tolerate salt by promoting ABA-operated GABA signaling control of the ascorbate–glutathione cycle
- Feasibility study of assessing cotton fiber maturity from near infrared hyperspectral imaging technique
- Effects of mepiquat chloride and plant population density on leaf photosynthesis and carbohydrate metabolism in upland cotton
- Risk of control failure to insecticides malathion, profenophos + cypermethrin mixture, and fipronil in boll weevil (Coleoptera:Curculionidae) populations from Bahia, Brazil
- Are yarn quality prediction tools useful in the breeding of high yielding and better fibre quality cotton (Gossypium hirsutum L.)?