
陆地棉耐镉胁迫的遗传与QTL定位分析
2024-01-01 00:00:00钟镇涛赵天伦陈进红祝水金
棉花学报 2024年3期
关键词:棉花



收稿日期:2021-04-30" " "第一作者简介:钟镇涛(1995―),男,硕士研究生,21816016@zju.edu.cn。 * 通信作者:shjzhu@zju.edu.cn
基金项目:国家重点研发计划(2016YFD0101400);国家棉花产业技术体系(CARS-18-25);江苏省现代作物生产协同创新中心
摘要:【目的】棉花是1种较耐重金属镉的作物,培育与种植耐镉胁迫棉花品种对于镉污染土壤的修复具有重要意义。本文旨在研究陆地棉耐镉性的遗传并对相关数量性状位点 (quantitative trait locus, QTL)进行定位分析。【方法】以陆地棉种质HS46和MARCABUCAG8US-1-88为亲本构建的188个重组自交系(recombinant inbred line, RIL)为材料,调查RIL群体在镉胁迫条件下的发芽率、发芽势,以及苗期叶片的叶绿素含量、株高和干/鲜物质质量等表型数据,并计算各家系的镉胁迫系数;采用完备区间作图法对RIL群体的各性状的镉胁迫系数及镉胁迫综合系数进行QTL定位和遗传分析。【结果】MARCABUCAG8US-1-88为耐镉材料,HS46则为镉敏感材料,188个RIL家系的耐镉性相关性状基本服从正态分布,包含强耐镉型材料23份、耐镉型材料49份、中等耐镉型材料67份、镉敏感型材料49份。连锁分析共鉴定到28个与镉胁迫系数相关的QTL,分布于14条染色体上,单个位点解释1.58%~8.41%的表型变异,其中根鲜物质质量镉胁迫系数位点qRFW-13-1在2个环境下均被检测到,分别解释7.66%和7.71%的表型变异。【结论】棉花耐镉性为多基因控制的数量遗传,受环境影响较大,定位到的耐镉QTL qRFW-13-1为稳定QTL,该结果对于棉花耐镉分子育种具有一定的应用价值。
关键词:棉花;镉胁迫;QTL定位
QTL mapping and genetic analysis of cadmium stress tolerance in upland cotton
Zhong Zhentao, Zhao Tianlun, Chen Jinhong, Zhu Shuijin*
(College of Agriculture and Biotechnology/Zhejiang Key Laboratory of Crop Germplasm, Zhejiang University, Hangzhou 310058, China)
Abstract: [Objective] Cotton is a relatively tolerant crop to cadmium (Cd) stress. It is of great significance to breeding and extending the Cd tolerant cotton cultivars for remediation of Cd contaminated soil. This study aims to analysis the inheritance of cotton tolerance to Cd stress and to map the quantitative trait locus (QTL) related to Cd stress. [Methods] In this study, 188 recombinant inbred lines (RIL) called HM188 derived from an intraspecific hybrid between HS46 and MARCABUCAG8US-
1-88 were used to study the phenotypic data of germination rate, germination potential, chlorophyll content, plant height, dry and fresh weight at seedling stage under Cd stress, and the Cd stress coefficient of each material was calculated as well. QTL mapping was conducted for each trait and Cd stress coefficient of HM188 population by using complete interval mapping method. [Results] According to the performance and Cd stress coefficient of each trait under Cd stress, MARCABUCAG8US-
1-88 was the Cd tolerant parent, while HS46 was a sensitive one. The Cd tolerance of 188 RIL was basically normal distribution, including 23 strong Cd tolerant lines, 49 Cd tolerant line, 67 medium tolerant lines, and 49 sensitive lines. A total of 28 QTL for Cd stress coefficient was identified, which were distributed on 14 chromosomes with explaining of variation of 1.58%~8.41%. Among them, qRFW-13-1 related to root fresh weight was detected in both environments, which explained variation of 7.66% and 7.71%, respectively. [Conclusion] Cd tolerance in upland cotton is a quantitative trait controlled by multiple genes, which was greatly affected by the environment. However, qRFW-13-1 for Cd tolerance may be a stable QTL, which has some application values in molecular breeding for Cd tolerant cotton.
Keywords: cotton; cadmium stress; QTL mapping
随着全球人口剧增,工农业生产以及日常生活废弃物对土壤产生了严重的污染。2014年我国原国土资源部和环境保护部联合发布的《全国土壤污染状况调查公报》显示,我国土壤环境状况总体不乐观,有19.4%的受调查耕地被污染,影响耕地环境质量的主要污染物是重金属,其中镉(cadmium,Cd)为首要污染物,在超过40%的受污染耕地中发现重金属Cd超标[1]。高浓度的Cd会对土壤生物产生毒害作用,并很容易转移到植物中,最终进入食物链[2]。摄入过多Cd会严重损害人体健康,导致癌症、骨质疏松和其他疾病[3]。因此,对Cd污染土壤的修复刻不容缓。目前针对重金属污染土壤的修复技术包括物理修复、化学修复、生物修复和农业生态修复技术等,生物修复法中的植物修复技术因成本低,可修复大面积污染土壤且具有一定绿化美化作用而备受关注。目前全世界已发现500多种富集重金属的植物,其中部分植物表现出对Cd的选择性吸收,如菠菜、烟草、向日葵等[4]。孙红艳[5]在大麦单双倍体(double haploid, DH)群体鉴定到11个与Cd含量相关的数量性状位点(quantitative trait locus, QTL),并研究了ZIP3、ZIP5、ZIP7和ZIP8这4个基因对大麦Cd转运与积累的调控与影响。潘晨阳等[6]在水稻重组自交系(recombinant inbred line, RIL)群体中鉴定到了7个Cd积累相关QTL,并筛选出LOCOs02g50240、LOCOs02g52780、LOCOs09g31200、LOCOs09g35030和LOCOs09g-
37949为耐Cd基因。
棉花是我国重要的经济作物。研究表明中低浓度Cd处理对棉花生长发育影响较小,棉花对土壤Cd具有良好的富集能力,棉纤维为非食用或饲用产品且Cd吸附量较低,棉花是理想的Cd污染耕地的修复作物之一[7-11]。植棉修复重金属污染耕地具有经济效益高、对土壤和环境友好、适用于重金属复合污染等优势[10]。然而,土壤中Cd质量浓度≥20 mg·kg-1时会严重抑制棉花种子萌发和幼苗生长发育,显著降低单株铃数、铃重、单铃种子数、籽棉产量和皮棉产量[12]。目前对于棉花耐Cd性和Cd积累的研究较少,尚未见与棉花耐Cd性和Cd积累相关的QTL报道,对于棉花耐Cd的遗传表现了解甚少,强耐Cd、高Cd富集棉花品种不足以满足Cd污染耕地修复的需要。本研究以陆地棉(Gossypium hirsutum)RIL群体为材料,研究Cd胁迫条件下棉花发芽和苗期生长情况,分析棉花苗期耐Cd性的遗传特点,并对棉花耐Cd性进行QTL定位,为耐Cd品种选育提供遗传资源和材料资源。
1 材料与方法
1.1 试验材料
以综合性状优良的陆地棉品种HS46和对多种逆境都有抗性的陆地棉种质系MARCABUCAG8US-1-88(简称MAR),以及两者杂交获得的188个RIL群体(简称HM188)为供试材料[13]。RIL通过单籽粒传法自交8代构建完成,自交繁殖保存。试验用种为2018年冬种于海南三亚收获的南繁自交种子,由浙江大学农学院棉花基因工程实验室提供。
1.2 试验方法
发芽试验:各材料脱绒种子晒干。发芽前用75%的酒精消毒,用无菌水冲洗干净后浸种12 h。挑选50粒种子均匀平铺到垫有双层滤纸的培养皿中,每皿加入600 μmol·L-1 CdCl2溶液20 mL进行Cd胁迫处理,以蒸馏水作对照,4次重复,放置于28 ℃、光暗周期为14 h/10 h的植物培养室中发芽,记录每天发芽种子数。
苗期试验:试验设2个环境,分别于2019年6-7月在浙江省杭州市浙江大学华家池校区实验站和2020年7-8月在浙江大学紫金港校区控温光生长室进行。采用裂区随机试验设计,主区为Cd胁迫处理(Cd),以非胁迫处理(CK)为对照;副区为RIL群体和双亲,对照为中棉所49。按试验设计,硫酸脱绒的饱满种子分别播于50(10×5)孔穴盘内,每个株系种植1行(10孔),每孔播2粒饱满的脱绒种子,加营养土至与孔口平。每穴盘中间1行播中棉所49,用以校正穴盘之间的试验误差。待幼苗1片真叶完全展开后,将处理组(Cd)用600 μmol·L-1 CdCl2溶液浇灌至饱和,保持10 d;对照组(CK)用水浇灌至饱和,保持10 d。处理10 d后,各处理每个株系随机取10株幼苗,分别测定其叶片叶绿素含量(以soil and plant analyzer development即SPAD值反映叶片叶绿素相对含量)、株高(plant height, PH)、根鲜物质质量(root fresh weight, RFW)、根干物质质量(root dry weight, RDW)、地上部鲜物质质量(shoot fresh weight, SFW)和地上部干物质质量(shoot dry weight, SDW),取平均值,为1次重复。试验设2次重复。2020年利用叶绿素分析仪(SPAD-502Plus,日本)测定幼苗1、2、3片真叶的SPAD值,取平均值为该株材料的SPAD值。
1.3 性状统计
利用Excel 2014和SPSS 20.0软件对亲本及RIL群体的表型数据进行统计分析和聚类分析,运用 Kolmogorov-Smirnov(D检验)和 Shapiro-
Wilk(W检验)进行正态分布检验。参照朱协飞等[14]的方法计算Cd胁迫系数:
Cd胁迫系数=×100%
参照岳松青等[15]的方法,计算各性状Cd胁迫系数的隶属函数值Xμ:
Xμ=;
式中,X为供试材料某一性状的Cd胁迫系数,Xmax为供试材料中某一性状Cd胁迫系数的最大值,Xmin为供试材料中某一性状Cd胁迫系数的最小值。将每个材料各性状的隶属函数值相加后取平均值,作为该品种的Cd胁迫综合系数,综合系数越小,该材料的耐Cd性越强。
1.4 连锁图谱与QTL定位
本研究所用RIL群体的连锁图谱由Li等[16]构建。利用QTL IciMapping 4.2软件中BIP模块,采用完备区间作图法(inclusive composite interval mapping,ICIM)分别对叶片SPAD值、株高、根干/鲜物质质量和地上部干/鲜物质质量进行QTL定位,参数设置:步长为1 厘摩(centiMorgan, cM),PIN为0.000 1,以对数优势比(logarithm of odd score,LOD)≥2.5作为阈值判断QTL存在。QTL命名规则[17]:q+性状英文名称缩写+染色体序号+QTL编号。
2 结果与分析
2.1 HM188群体及双亲的发芽情况及苗期耐Cd表现
2个环境下的HM188及其亲本在Cd胁迫条件下的发芽情况、苗期生长情况及对应的Cd胁迫系数统计结果见表1~3。
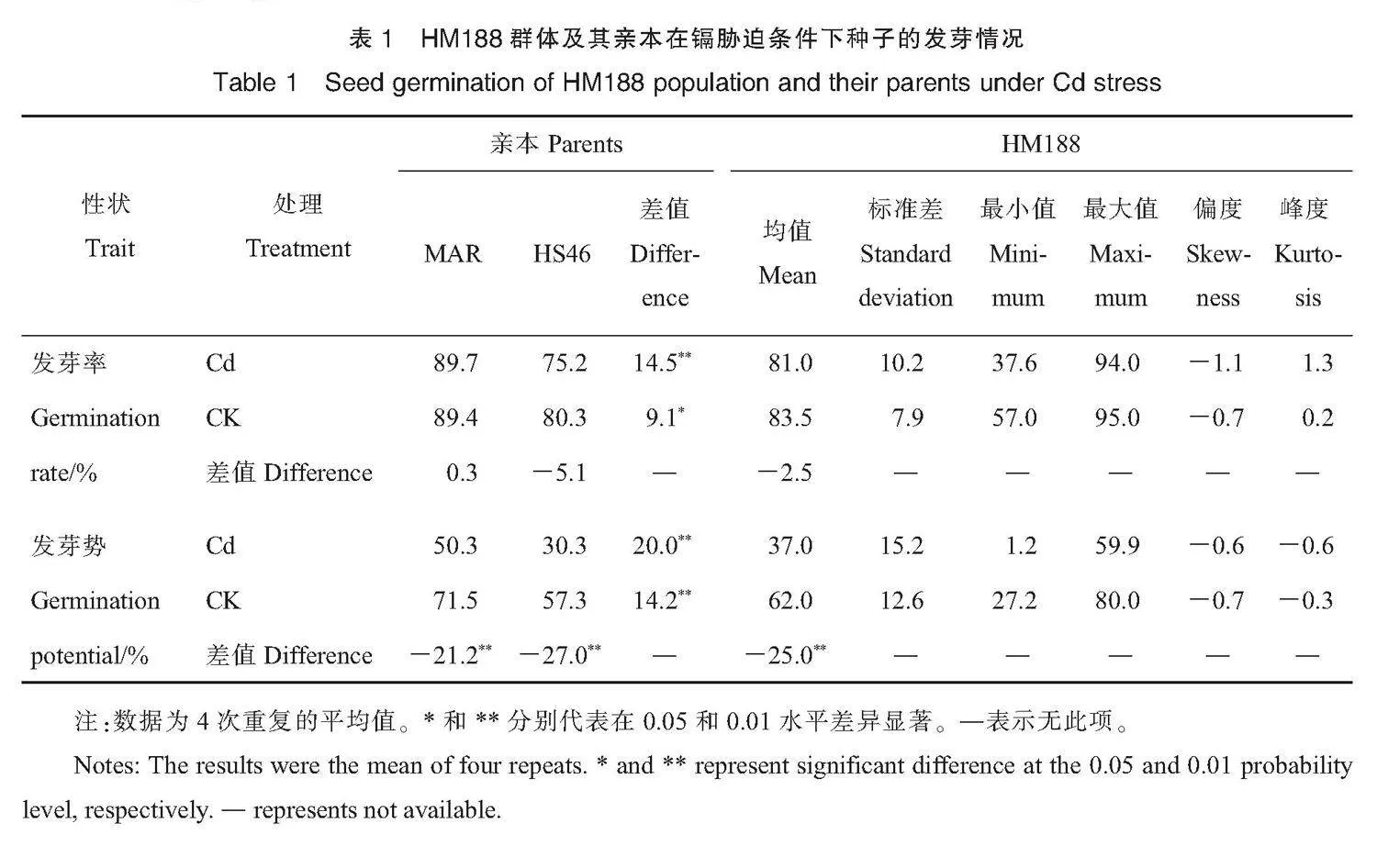
从表1来看,在对照和Cd胁迫条件下,亲本MAR的发芽率、发芽势均显著高于HS46,且MAR发芽率和发芽势的Cd胁迫系数显著低于HS46(表3),说明2个亲本的耐Cd性存在显著差异,MAR较HS46耐Cd。在Cd胁迫条件下,HM188群体的平均发芽势显著低于对照,发芽率与对照无显著差异。在对照条件下,HM188的发芽率为57.0%~95.0%,表现出超亲分离现象。在Cd胁迫下,HM188的发芽率为37.6%~94.0%,变异幅度更大。在Cd胁迫条件下,HM188群体的发芽势为1.2%~59.9%,表现出超亲分离现象。这些结果表明,在Cd胁迫下棉花种子能发芽,但种子活力显著下降。
胁迫系数结果(表3)表明,HM188群体的发芽势在Cd胁迫条件下平均下降了41.18%,发芽势的Cd胁迫系数(-18.80%~97.50%)呈连续分布,偏度与峰度绝对值均小于1,基本符合正态分布,且表现出超亲分离。说明Cd胁迫下的发芽势可作为棉花耐Cd性指标之一。
在正常和Cd胁迫处理条件下,亲本MAR和HS46间的SPAD值均无显著差异,但在Cd胁迫条件下MAR和HS46的SPAD值较对照分别下降了17.5%和28.3%。HM188群体的SPAD平均值在Cd胁迫条件下显著低于无胁迫的对照。胁迫系数结果表明,亲本MAR叶片SPAD值的Cd胁迫系数显著低于HS46,HM188群体的叶片SPAD值在Cd胁迫条件下平均下降了23.68%,SPAD值的Cd胁迫系数呈连续分布(18.52%~29.54%),且表现出超亲分离,Cd胁迫系数分布的偏度与峰度绝对值均小于1,符合正态分布。尽管只有1年的测定数据,但也能够说明Cd胁迫影响植株叶片的SPAD值并影响光合作用。
在正常条件下,亲本MAR的株高显著低于HS46,而Cd胁迫条件下MAR的株高显著高于HS46(表2),尽管2个环境的结果不尽一致,但趋势相同(结果未显示)。亲本MAR的株高Cd胁迫系数显著低于HS46(表3)。Cd胁迫条件下HM188群体的株高为8.1~11.0 cm,平均为9.8 cm,显著低于CK,平均下降了30.50%。亲本MAR和HS46的株高Cd胁迫系数分别为22.97%和37.23%;HM188的株高Cd胁迫系数呈连续分布,变幅为21.47%~40.97%,具有超亲表现,Cd胁迫系数分布的偏度与峰度绝对值均小于1,符合正态分布。Cd胁迫下的株高可作为棉花耐Cd性指标之一。
在正常条件下,亲本MAR的根鲜物质质量和根干物质质量均显著低于HS46,而在Cd胁迫条件下2个亲本的根鲜物质质量和根干物质质量均无显著差异(表2)。在Cd胁迫条件下,HM188群体的根鲜物质质量和根干物质质量分别为0.462~0.823 g和0.032~0.052 g,平均分别为0.635 g和0.037 g,比CK分别显著降低29.99%和35.09%(表2)。从表3来看,亲本MAR根鲜物质质量、根干物质质量的Cd胁迫系数显著低于HS46;在Cd胁迫条件下HM188的根鲜物质质量和干物质质量的Cd胁迫系数呈连续分布,分别为4.88%~44.44%和12.02%~49.15%,均表现出超亲现象,偏度与峰度绝对值均小于1,符合正态分布。Cd胁迫下的根鲜物质质量和干物质质量可作为棉花耐Cd性指标之一。
CK条件下,亲本MAR的地上部鲜物质质量和干物质质量均显著低于HS46,在Cd胁迫条件下,2个亲本的地上部鲜物质质量和地上部干物质质量均无显著差异(表2)。Cd胁迫条件下,HM188群体的地上部鲜物质质量和干物质质量变幅分别为1.421~2.097 g和0.161~0.245 g,平均分别为1.766 g和0.203 g,比CK显著降低32.08%和36.36%(表2)。胁迫系数(表3)结果表明,亲本MAR地上部鲜物质质量和干物质质量的Cd胁迫系数均显著低于HS46,地上部鲜物质质量和干物质质量的Cd胁迫系数呈连续分布,并有超亲表现,分布的偏度与峰度绝对值均小于1,符合正态分布。Cd胁迫下的地上部鲜物质质量和干物质质量可作为棉花耐Cd性指标之一。
Cd胁迫条件下HM188群体的发芽势、叶片SPAD值、株高、根干/鲜物质质量、地上部干/鲜物质质量比CK显著降低。亲本MAR的Cd胁迫综合系数显著低于HS46,表现出良好的耐Cd性。
2.2 HM188群体耐Cd性的聚类分析和遗传力分析
采用最长距离法对各材料的Cd胁迫综合系数进行聚类,结果见图1。聚类分析表明,HM188群体和双亲被划分为4个类群:Ⅰ类为强耐Cd型材料,包含亲本MAR、RIL-6、RIL-8和RIL-14等24份材料,占供试材料的12.6%;Ⅱ类为耐Cd型材料,包含RIL-5、RIL-7和RIL-13等49份材料,占供试材料的25.8%;III类为中等耐Cd型材料,包含RIL-1、RIL-2和RIL-4等67份材料,占供试材料的35.3%;IV类为Cd敏感型材料,包含亲本HS46、RIL-9、RIL-10和RIL-32等50份材料,占供试材料的26.3%。
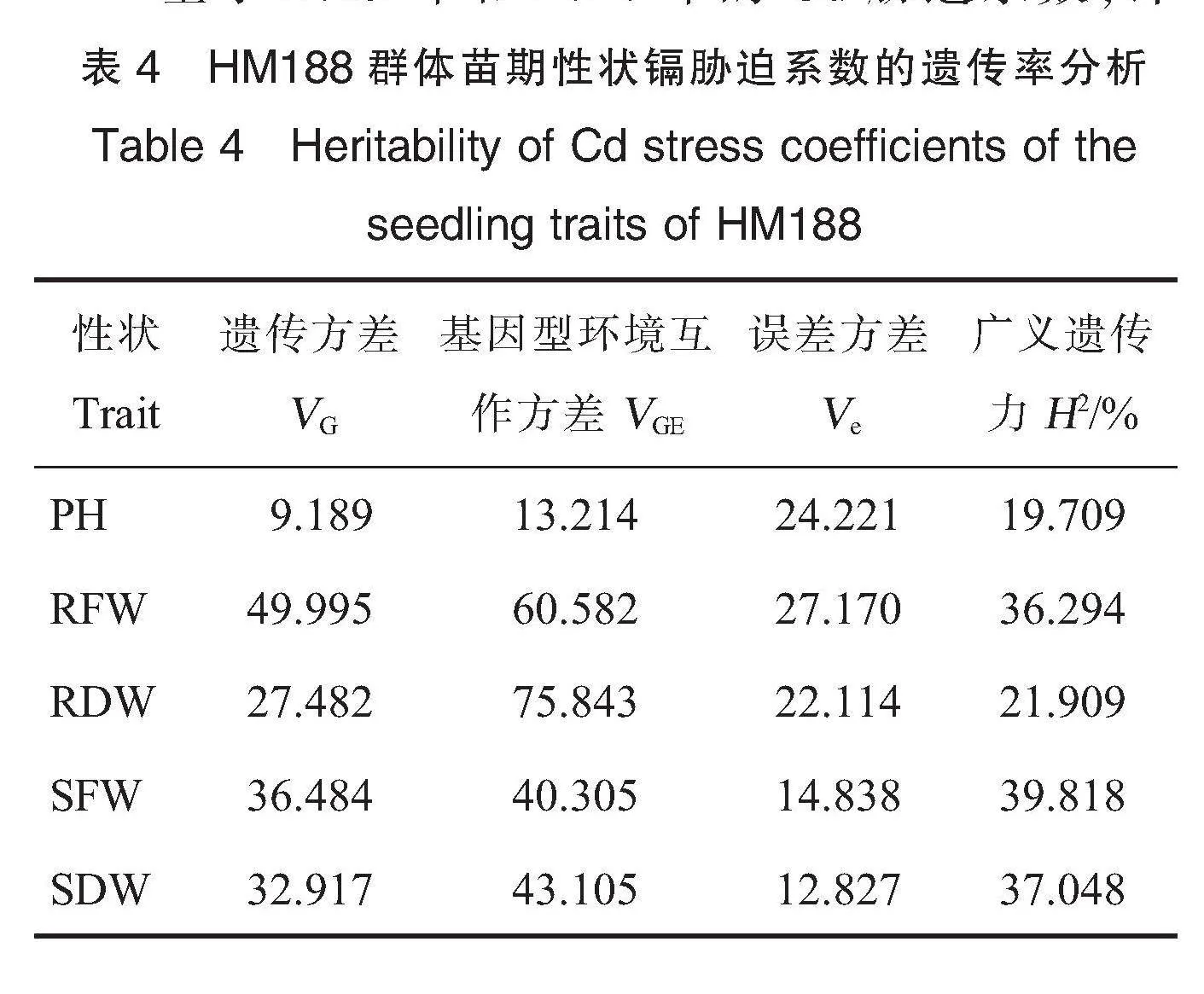
基于2019年和2020年的Cd胁迫系数,计算株高、根鲜物质质量、根干物质质量、地上部鲜物质质量和地上部干物质质量Cd胁迫系数的广义遗传率(H2),分别为19.709%、36.294%、21.909%、39.818%和37.048 %(表4),其中地上部鲜物质质量Cd胁迫系数的广义遗传率相对最高,地上部干物质质量的次之,株高的最低。可见陆地棉耐Cd性的遗传率较低,受环境影响较大,遗传改良较为困难。
2.3 Cd胁迫系数的QTL定位
HM188群体的发芽率、发芽势、叶片SPAD值、株高、根干/鲜物质质量、地上部干/鲜物质质量这8个性状的Cd胁迫系数及Cd胁迫综合系数均呈连续性分布,偏度与峰度绝对值均小于1,基本服从正态分布,适合用于QTL定位(表3)。
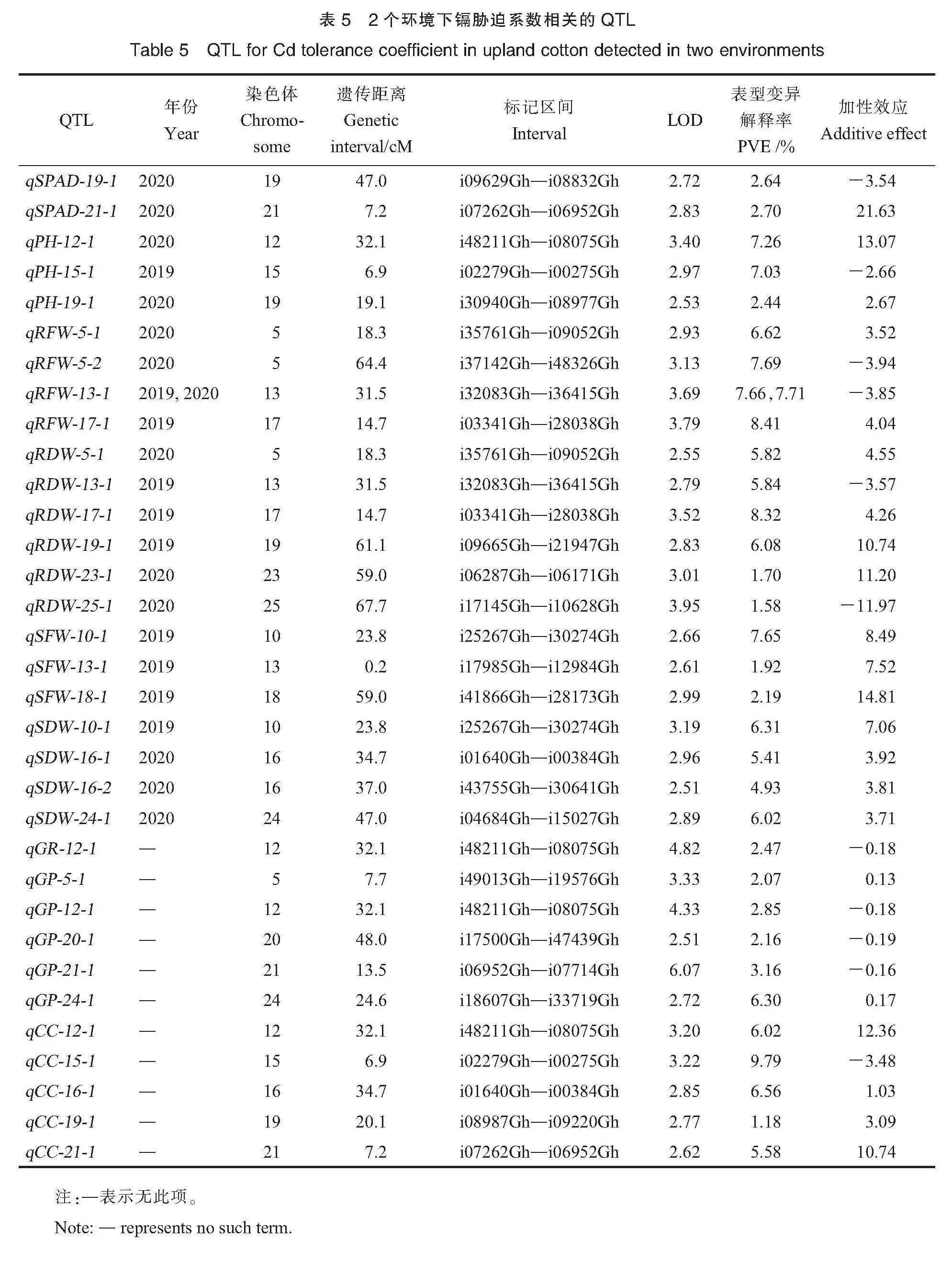
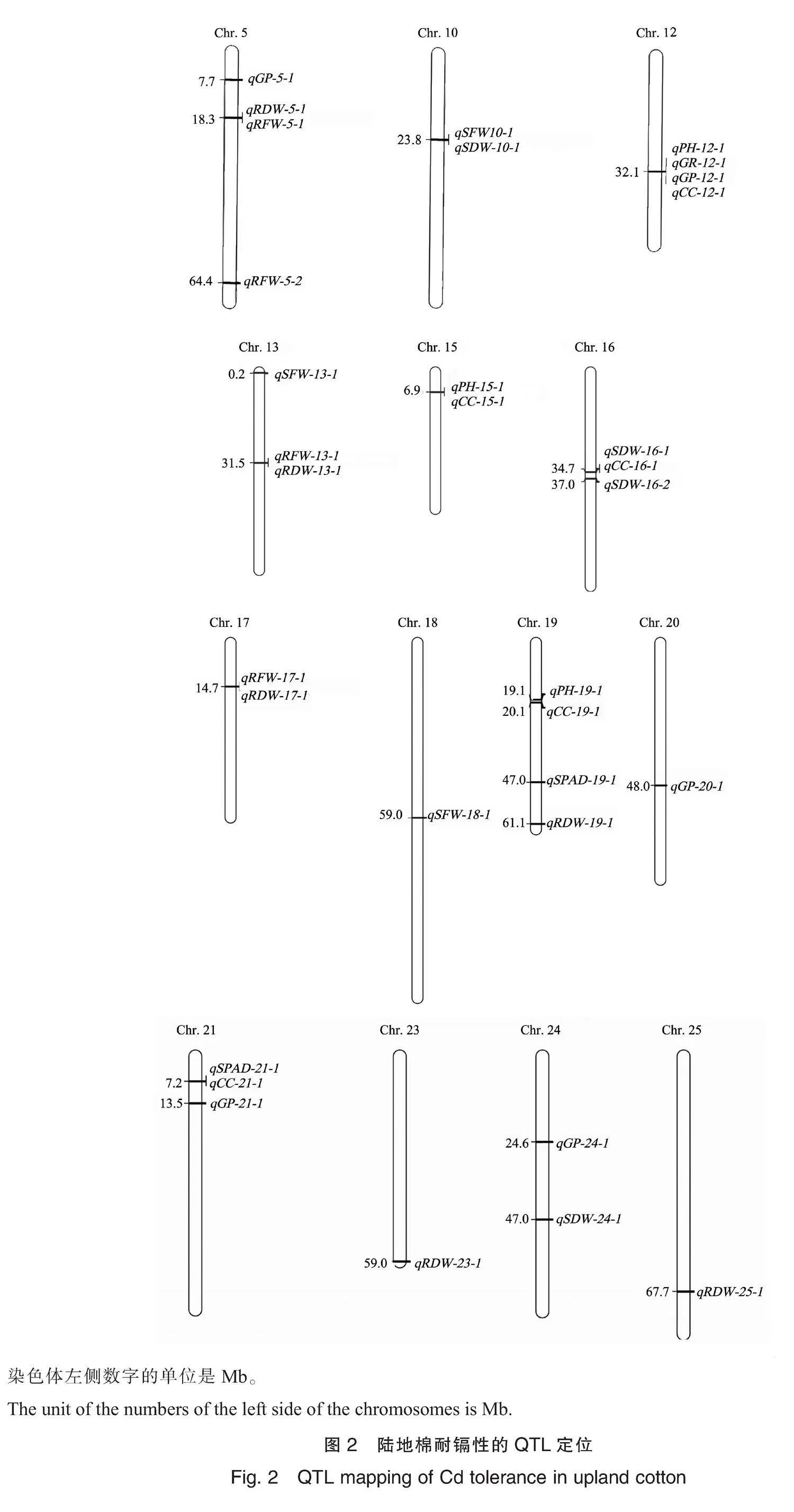
通过QTL定位,在2个环境下共检测到28个与这些性状的Cd胁迫系数相关的QTL,5个与Cd胁迫综合系数相关的QTL,分布在5、10、12、13、15、16、17、18、19、20、21、23、24和25号染色体上(表5、图2)。
叶片SPAD值:共检测到2个与叶片SPAD值 Cd胁迫系数相关的QTL,分布在19和21号染色体上,解释2.64%~2.70%的表型变异,qSPAD-19-1的有利等位变异来自HS46,qSPAD-
21-1的有利等位变异来自MAR。
株高:共检测到3个与株高Cd胁迫系数相关的QTL,分布在12、15和19号染色体上,解释2.44%~7.26%的表型变异,qPH-15-1的有利等位变异来自HS46,qPH-12-1和qPH-19-1的有利等位变异来自MAR。
根鲜物质质量:共检测到4个与根鲜物质质量Cd胁迫系数相关的QTL,5号染色体上2个、13和17号染色体上各1个,解释6.62%~8.41%的表型变异。其中qRFW-13-1在2019和2020年2个环境下均被检测到,分别解释7.66%和7.71%的表型变异。
根干物质质量:共检测到6个与根干物质质量Cd胁迫系数相关的QTL,5、13、17、19、23和25号染色体上各1个,解释了1.58%~8.32%的表型变异。其中qRDW-5-1与根鲜物质质量Cd胁迫系数位点qRFW-5-1为同一位点,有利等位变异来自MAR,qRDW-13-1与根鲜物质质量Cd胁迫系数位点qRFW-13-1为同一位点,有利等位变异来自HS46,qRDW-17-1与根鲜物质质量Cd胁迫系数位点qRFW-17-1为同一位点,有利等位变异来自MAR。
地上部鲜物质质量:共检测到3个与地上部鲜物质质量Cd胁迫系数相关的QTL,10、13和18号染色体上各1个,解释1.92%~7.65%的表型变异,有利等位变异均来自MAR。
地上部干物质质量:共检测到4个与地上部干物质质量Cd胁迫系数相关的QTL,16号染色体上2个,10和24号染色体上各1个,解释4.93%~6.31%的表型变异,4个QTL的有利等位变异均来自MAR。其中qSDW-10-1和地上部鲜物质质量Cd胁迫系数位点qSFW-10-1为同一位点。
共检测到1个与发芽率Cd胁迫系数相关的QTL,位于12号染色体,解释2.47%的表型变异。共检测到5个与发芽势Cd胁迫系数相关的QTL,5、12、20、21和24号染色体上各1个,解释2.07%~6.30%的表型变异。其中,qGP-12-1和发芽率Cd胁迫系数位点qGR-12-1、株高Cd胁迫系数位点qPH-12-1为同一位点,其有利等位变异来自HS46。
Cd胁迫综合系数(comprehensive coefficient, CC):共检测到5个与Cd胁迫综合系数相关的QTL,分别位于12、15、16、19和21号染色体上,解释1.18%~9.79%的表型变异。其中,qCC-12-1和株高Cd胁迫系数位点qPH-12-1、发芽率Cd胁迫系数位点qGR-12-1、发芽势Cd胁迫系数位点qGP-12-1为同一位点,qCC-15-1和株高Cd胁迫系数位点qPH-15-1为同一位点,qCC-16-1和地上部干物质质量Cd胁迫系数位点qSDW-16-1为同一位点,qCC-21-1和叶片SPAD值Cd胁迫系数位点qSPAD-21-1为同一位点。
3 讨论
本研究构建HM188群体的亲本为陆地棉HS46和MARCABUCAGSUS-l-88,两者亲缘关系较远, HS46是高产优质陆地棉品种, MAR是多抗的陆地棉种质资源。2个亲本在农艺性状、纤维品质和抗病性等方面都存在显著的差异,但目前缺少关于该群体对低温、高温、干旱、涝渍和重金属等非生物胁迫的耐性/抗性的研究。
前人已对HM188群体的抗病性、种子品质、纤维品质、产量等方面开展了遗传效应分析和QTL定位等工作。任爱霞等[17]调查了该群体枯萎病和黄萎病的田间发病情况,比较抗病、非抗病株系的比例,得出枯萎病抗性与黄萎病抗性分别为数量性状和质量性状。栾启福等[13]利用包括基因型与环境互作在内的加性模型分析该群体,证实枯萎病抗病能力主要受加性效应影响。孔广超等[19]调查了该群体在4个环境下的产量及纤维品质性状的表型数据,通过连锁分析检测到109个与籽棉产量、皮棉产量、衣分、铃重、籽指和衣指相关的QTL。秦利[20]将该群体株系间随机组配,构建了1套包括376个组合的永久F2群体,检测到控制籽指、仁指、种仁率、蛋白质含量、油分含量和棉酚含量相关的加性QTL 18个、上位性QTL 12个,解释性状2.42%~23.04%的表型变异。Li等[21]将2个亲本HS46和MAR分别作为父本与HM188群体回交创建了2个陆地棉回交F1群体,定位到205个与产量性状相关的QTL、195个与纤维品质性状相关的QTL、138个与产量性状杂种优势相关的QTL和65个与纤维品质性状杂种优势相关的QTL。
耐Cd性苗期鉴定因环境问题而受到限制。本研究苗期试验采用穴盘种植,通过在每个穴盘上种植1行中棉所49作为数据校准依据,减少穴盘间差异导致的误差。Cd胁迫综合系数计算采用隶属函数法,由于目前尚无文献明确得出棉花各表型性状的权重,参照王准等[22]的方法,采用客观赋权法计算各性状的权重进行综合评价,为棉花耐重金属胁迫鉴定提供借鉴。采用该方法对HM188群体的苗期耐Cd性进行鉴定和QTL定位,其中qRFW-13-1位点在2019和2020年共2个环境下均被检测到,qPH-12-1/qGR-12-1/qGP-
12-1、qPH-15-1、qSDW-16-1、qSPAD-21-1这4个位点在性状Cd胁迫系数转换为Cd胁迫综合系数后同样可以定位到,可以为陆地棉耐Cd相关候选基因的筛选和克隆提供参考。
4 结论
通过对HM188群体及双亲的发芽率、发芽势以及5个苗期性状的鉴定,发现HM188群体及双亲的耐Cd性变异丰富,其中强耐Cd型材料24份、耐Cd型材料49份、中等耐Cd型材料67份、Cd敏感型材料50份,为棉花耐Cd性的遗传研究和品种选育提供了宝贵的种质材料。通过连锁分析检测到28个与陆地棉苗期表型性状Cd胁迫系数相关的QTL和5个与Cd胁迫综合系数相关的QTL,填补了陆地棉耐Cd相关遗传定位研究的空白,可为棉花耐Cd分子育种提供参考。但本研究仅针对陆地棉种子萌发期和苗期进行胁迫研究,对于Cd胁迫对陆地棉产量、纤维品质以及各组织Cd吸收量的研究需要在后续的试验中进行补充。
参考文献:
[1]中华人民共和国环境保护部. 全国土壤污染状况调查公报[EB/OL]. (2012-04-17) [2021-04-25]. http://www.gov.cn/foot/
site1/20140417/782bcb88840814ba158d01.pdf.
Ministry of Environmental Protection of the People's Republic of China. Bulletin of national soil pollution survey[EB/OL]. (2012-
04-17) [2021-04-25]. http://www.gov.cn/foot/site1/20140417/
782bcb88840814ba158d01.pdf.
[2] Li Y S, Tang H, Hu Y X, et al. Enrofloxacin at environmentally relevant concentrations enhances uptake and toxicity of cadmium in the earthworm Eisenia fetida in farm soils[J/OL]. Journal of Hazardous Materials, 2016, 308: 312-320[2021-04-25]. https://
doi.org/10.1016/j.jhazmat.2016.01.057.
[3] Moynihan M, Peterson K E, Cantoral A, et al. Dietary predictors of urinary cadmium among pregnant women and children [J/OL]. Science of The Total Environment, 2017, 575: 1255-
1262[2021-04-25]. https://doi.org/10.1016/j.scitotenv.2016.09.
204.
[4] 卢红玲, 肖光辉, 刘青山, 等. 土壤Cd污染现状及其治理措施研究进展[J/OL]. 南方农业学报, 2014, 45(11): 1986-1993[2021-04-25]. https://dio.org/10.3969/j.issn.2095-1191.2014.11.
1986.
Lu Hongling, Xiao Guanghui, Liu Qingshan, et al. Advances in soil Cd pollution and solution measures[J/OL]. Journal of Southern Agriculture, 2014, 45(11): 1986-1993[2021-04-25]. https://dio.org/10.3969/j.issn.2095-1191.2014.11.1986.
[5] 孙红艳. 大麦耐Cd及Cd积累相关QTL定位与候选基因筛选及调控研究[D]. 杭州: 浙江大学, 2012.
Sun Hongyan. Identification of QTLs and candidate genes associated with cadmium tolerance and accumulation in barley and its regulation[D]. Hangzhou: Zhejiang University, 2012.
[6] 潘晨阳, 叶涵斐, 周维永, 等. 水稻籽粒Cd积累QTL定位及候选基因分析[J/OL]. 植物学报, 2021, 56(1): 25-32[2021-04-25]. https://doi.org/10.11983/CBB20148.
Pan Chenyang, Ye Hanfei, Zhou Weiyong, et al. QTL mapping of candidate genes involved in Cd accumulation in rice grain [J/OL]. Chinese Bulletin of Botany, 2021, 56(1): 25-32[2021-
04-25]. https://doi.org/10.11983/CBB20148.
[7] Kirungu J N, Magwanga R O, Shiraku M L, et al. Genome-wide identification and analysis of the CNGC gene family in upland cotton under multiple stress conditions[J/OL]. Journal of Cotton Research, 2023, 6: 17[2024-04-25]. https://doi.org/10.1186/
s42397-023-00152-z.
[8] 郭利双, 陈浩东, 贺云新, 等. Cd高积累棉花品种水培筛选结果初报[J/OL]. 中国棉花, 2015, 42(10): 14-16[2021-04-25]. https://
doi.org/10.11963/issn.1000-632X.201510005.
Guo Lishuang, Chen Haodong, He Yunxin, et al. Preliminary results on cotton cultivars screening for high accumulation of cadmium[J/OL]. China Cotton, 2015, 42(10): 14-16[2021-04-
25]. https://doi.org/10.11963/issn.1000-632X.201510005.
[9] 李玲, 陈进红, 何秋伶, 等. 3个陆地棉种质系重金属Cd的积累、转运和富集特性分析[J/OL]. 棉花学报, 2012, 24(6): 535-
540[2021-04-25]. https://doi.org/10.3969/j.issn.1002-7807.2012.
06.010.
Li Ling, Chen Jinhong, He Qiuling, et al. Accumulation, transportation, and bioconcentration of cadmium in three upland cotton plants under cadmium stress[J/OL]. Cotton Science, 2012, 24(6): 535-540[2021-04-25]. https://doi.org/10.3969/j.issn.1002-
7807.2012.06.010.
[10] 樊霆, 叶文玲, 陈海燕, 等. 农田土壤重金属污染状况及修复技术研究[J]. 生态环境学报, 2013, 22(10): 1727-1736.
Fan Ting, Ye Wenling, Chen Haiyan, et al. Review on contamination and remediation technology of heavy metal in agricultural soil[J]. Ecology and Environmental Sciences, 2013, 22(10): 1727-1736.
[11] 秦普丰, 铁柏清, 周细红, 等. 铅与Cd对棉花和水稻萌发及生长的影响[J/OL]. 湖南农业大学学报, 2000, 26(3): 205-207[2021-04-25]. https://doi.org/10.3321/j.issn:1007-1032.2000.
03.013.
Qin Pufeng, Tie Baiqing, Zhou Xihong, et al. Effects of cadmium and lead in soil on the germination and growth of rice and cotton[J/OL]. Journal of Hunan Agricultural University, 2000, 26(3): 205-207[2021-04-25]. https://doi.org/10.3321/j.issn:1007-
1032.2000.03.013.
[12] 刘连涛, 陈静, 孙红春, 等. Cd胁迫对棉花幼苗生长效应及不同器官Cd积累的影响[J/OL]. 棉花学报, 2014, 26(5): 466-
470[2021-04-25]. https://doi.org/10.3969/j.issn.1002-7807.2014.
05.012.
Liu Liantao, Chen Jing, Sun Hongchun, et al. Effects of cadmium stress on growth and cadmium accumulation in cotton (Gossy-pium hirsutum L.) seedlings[J/OL]. Cotton Science, 2014, 26(5): 466-470[2021-04-25]. https://doi.org/10.3969/j.issn.1002-7807.
2014.05.012.
[13] 栾启福, 祝水金. 陆地棉重组近交系HM188及其性状表现[J]. 棉花学报, 2003, 15(4): 252-253.
Luan Qifu, Zhu Shuijin. Introduction of a recombinant inbred line of upland cotton (G. hirsutum L.) HM188[J]. Cotton Science, 2003, 15(4): 252-253.
[14] 朱协飞, 司占峰. 棉花导入系耐盐性鉴定及耐盐基因QTL定位[J/OL]. 棉花学报, 2019, 31(1): 23-30[2021-04-25]. https://
doi.org/10.11963/1002-7807.zxfzxf.20181226.
Zhu Xiefei, Si Zhanfeng. Evaluation and QTL mapping of tolerance to salinity using interspecific introgression lines from Gossypium barbadense in G. hirsutum[J/OL]. Cotton Science, 2019, 31(1): 23-30[2021-04-25]. https://doi.org/10.11963/1002-
7807.zxfzxf.20181226.
[15] 岳松青, 曹辉, 荀咪, 等. 基于指标综合分析的苹果砧木耐Cd性评价[J]. 植物生理学报, 2019, 55(5): 649-656.
Yue Songqing, Cao Hui, Xun Mi, et al. Evaluation of cadmium tolerance for apple rootstocks based on index comprehensive analysis[J]. Plant Physiology Journal, 2019, 55 (5): 649-656.
[16] Li C, Dong Y T, Zhao T L, et al. Genome-wide SNP linkage mapping and QTL analysis for fiber quality and yield traits in the upland cotton recombinant inbred lines population[J/OL]. Frontiers in Plant Science, 2016, 7: 1356[2021-04-25]. https://
doi.org/10.3389/fpls.2016.01356.
[17] McCouch S R, Chen X L, Panaud O, et al. Microsatellite marker development, mapping and applications in rice genetics and breeding[J]. Plant Molecular Biology, 1997, 35(1/2): 89-99.
[18] 任爱霞, 胡家恕, 祝水金, 等. 棉花黄萎病抗性与过氧化物酶同工酶分析[J]. 棉花学报, 2002, 14(5): 273-276.
Ren Aixia, Hu Jiasu, Zhu Shuijin, et al. Analysis of peroxidase isozymes and resistance to Verticillium dahliae in cotton[J]. Cotton Science, 2002, 14(5): 273-276.
[19] 孔广超, 秦利, 徐海明, 等. 棉花IF2群体构建及其在纤维品质遗传和杂种优势研究中的应用[J]. 作物学报, 2010, 36(6): 940-944.
Kong Guangchao, Qin Li, Xu Haiming, et al. Construction of IF2 population and its application in studies on genetic effects and heterosis for fiber quality in upland cotton (G. hirsutum L.)[J]. Acta Agronomica Sinica, 2010, 36(6): 940-944.
[20] 秦利. 陆地棉种子品质性状遗传及其QTL定位研究[D]. 杭州: 浙江大学, 2009.
Qin Li. Inheritance of seed quality traits and their QTL mapping in upland cotton (G. hirsutum L.)[D]. Hangzhou: Zhejiang University, 2009.
[21] Li C, Yu H R, Li C, et al. QTL Mapping and heterosis analysis for fiber quality traits across multiple genetic populations and environments in upland cotton[J/OL]. Frontiers in Plant Science, 2018, 9: 1364[2021-04-25]. https://doi.org/ 10.3389/fpls.2018.
01364.
[22] 王准, 张恒恒, 董强, 等. 棉花耐低氮和氮敏感种质筛选及验证[J/OL]. 棉花学报, 2020, 32(6): 538-551[2021-04-25]. https://doi.org/10.11963/1002-7807.wzsmz.20201023.
Wang Zhun, Zhang Hengheng, Dong Qiang, et al. Screening and verification of low nitrogen tolerant and nitrogen sensitive cotton germplasm[J/OL]. Cotton Science, 2020, 32(6): 538-551[2021-04-25]. https://doi.org/10.11963/1002-7807.wzsmz.20201023.
(责任编辑:王国鑫" " 责任校对:王小璐)" ●
猜你喜欢
作文周刊·小学一年级版(2023年36期)2023-09-14 14:32:41
少儿科学周刊·儿童版(2022年10期)2022-06-27 14:06:58
少儿科学周刊·儿童版(2022年10期)2022-06-27 02:09:44
小读者(2020年4期)2020-06-16 03:33:54
学苑创造·A版(2017年9期)2017-09-25 17:47:18
小溪流(画刊)(2017年1期)2017-03-16 16:07:03
小布老虎(2016年8期)2016-12-01 05:46:26
红领巾·萌芽(2015年7期)2015-09-10 07:22:44
红领巾·萌芽(2015年6期)2015-08-14 21:32:14
小天使·一年级语数英综合(2014年7期)2014-06-26 14:42:38
