
4种长苞铁杉群落木质残体生物量及碳储量特征
2024-01-01 00:00:00倪舒静肖石红曹彦梁瀞游巍斌何东进
林业科学研究 2024年3期


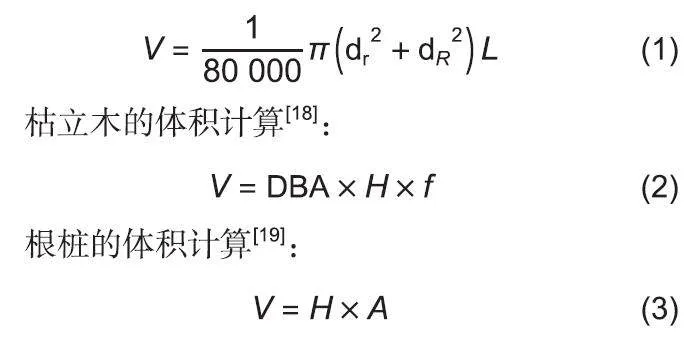
摘要:[目的]揭示天宝岩国家级自然保护区内长苞铁杉群落木质残体在碳循环中的基础作用,为长苞铁杉群落保护与森林可持续经营提供依据。[方法]以天宝岩国家级自然保护区4种长苞铁杉群落(长苞铁杉纯林、长苞铁杉+阔叶树混交林、长苞铁杉+猴头杜鹃混交林和长苞铁杉+毛竹混交林)为研究对象,对林内不同树种、不同存在形式(细木质残体、倒木、枯立木和树桩)开展动态调查(2012年、2014年、2016年),估算木质残体生物量和碳储量并分析其动态变化。[结果]表明:(1)4种群落内不同树种木质残体均是长苞铁杉的生物量占比最大(31.57%~77.77%);(2)4种长苞铁杉群落同一林型不同年份相同存在形式的木质残体类型生物量差异均不显著(pgt;0.05),同一林型不同年份的木质残体类型均是倒木生物量最高(长苞铁杉纯林:5.82~9.48t·hm-2;长苞铁杉+阔叶树混交林:10.90~17.83t·hm-2;长苞铁杉+猴头杜鹃混交林:7.20~10.37 t·hm-2;长苞铁杉+毛竹混交林:6.11~9.77 t·hm-2);3次调查结果中长苞铁杉纯林枯立木生物量与其他3种群落枯立木生物量均差异显著(p<0.05);(3)4种长苞铁杉群落均是长苞铁杉的碳储量最多,且随着年份的增长,其碳储量总体呈现增长趋势(长苞铁杉纯林:3.64~4.62 t·hm-2;长苞铁杉+阔叶树混交林:4.67~4.71 t·hm-2;长苞铁杉+猴头杜鹃混交林:2.29~2.53 t·hm-2;长苞铁杉+毛竹混交林:3.10~4.79 t·hm-2);(4)4种长苞铁杉群落木质残体中均是倒木的碳储量最高,除长苞铁杉+毛竹混交林倒木碳储量是先减少后增加外,其他3种长苞铁杉群落倒木的碳储量均呈现上升趋势,且4种群落中,长苞铁杉+阔叶树混交林的倒木碳储量最高,为4.91~7.68 t·hm-2;3次调查结果中,长苞铁杉纯林枯立木碳储量与其他3种混交林枯立木碳储量均具有显著差异(p<0.05)。[结论]p4种长苞铁杉群落中,不同形式木质残体中均是倒木的生物量和碳储量最高;不同树种木质残体均是长苞铁杉的生物量和碳储量最高。长苞铁杉纯林内枯立木生物量、碳储量与其他群落内枯立木差异显著(p<0.05)。
关键词:天宝岩国家级自然保护区;长苞铁杉群落;木质残体;生物量;碳储量
中图分类号:S718.5 文献标识码:A 文章编号:1001-1498(2024)03-0013-11
木质残体(Woody debris,WD)主要指森林生态系统中倒木、枯立木、根桩和大枯枝等粗木质残体(Coarse woody debris,CWD),以及小枝和碎根残片等细木质残体(Fine woody debris,FWD)。它们在森林生态系统的生物地球化学循环中起到关键作用,并对全球的碳平衡和碳循环有着重要影响,是生态系统中重要的碳库和养分库。
长苞铁杉(Tsuga longibracteata Cheng)林是天宝岩国家级自然保护区森林的主体,拥有维持区域生态系统平衡、涵养水源和保持水土等重要生态作用,对古生态、古气候具有重要的研究意义。而目前长苞铁杉的分布区不断缩小,集群规模不断下降,林内多为成熟或过熟个体,种群处于衰退期,天然更新困难,且多生长于山脊岩石裸露区,一旦遭到破坏,极难恢复与更新。目前国内学者对长苞铁杉群落木质残体的研究主要有:肖石红等对长苞铁杉林粗死木质残体现存量进行了研究;穆振北等对天宝岩长苞铁杉林木质残体径级和腐烂等级的研究表明,木质残体的物理性质是影响养分含量变化的关键因素;何东进等系统探究了长苞铁杉林倒木的结构、倒木的分解者、倒木的生态功能及倒木的环境效应等特征。这些研究为全面了解长苞铁杉群落及其木质残体的生态功能提供了不可或缺的数据,但对长苞铁杉群落木质残体生物量、碳储量方面却少有研究。
鉴于长苞铁杉林的重要价值及其木质残体对长苞铁杉群落的重要影响,本研究以天宝岩国家级自然保护区内的4种长苞铁杉群落为研究对象,探究其林内不同形式木质残体的生物量和碳储量动态变化,旨在了解其在群落养分循环与碳循环中的重要作用,为长苞铁杉群落保护与发展提供科学参考。
1 研究区概况和研究方法
1.1 研究区概况
福建天宝岩国家级自然保护区(25°50'51"~26°1'20"N,117°28'3"~117°35'28"E)地处福建省永安市境内,为中亚热带季风气候区,年平均降水量约为2 039 mm,年平均气温为15℃,气候温暖湿润,无霜期长,约290 d,天宝岩为保护区内最高峰。保护区内植物物种多样性十分丰富,包括长苞铁杉、马尾松(Pinus massonianaLamb)、青冈(Quercus glauca Thunb)等主要乔木树种;以及猴头杜鹃(Rhododendronsimiarum Hance)、柃木(Eurya japonicaThunb)等主要灌木树种。保护区内的长苞铁杉林以及猴头杜鹃林是中国南方其他地方少见或没有的,具有很高的研究价值。尤其是珍稀濒危植物长苞铁杉林分布面积高达186.7 hm2,其中,原生性纯林面积达20 hm2,其纯林面积为我国之最,目前已列入省级重点保护植物,具有极高的保护价值和研究价值。
1.2 研究方法
1.2.1 样地设置与木质残体调查
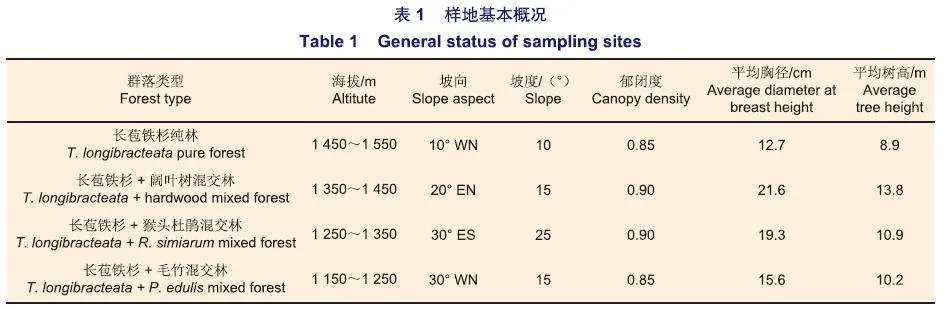
选取长苞铁杉纯林、长苞铁杉+阔叶树混交林、长苞铁杉+猴头杜鹃混交林和长苞铁杉+毛竹混交林4个海拔梯度的4种群落类型作为研究对象,在4种群落类型中选择地形相对一致、林相相对整齐的具有一定代表性的地段作为典型样地,每种群落类型均设置3个20m×30m的固定样地,样地总面积为7 200m2(表1)。于2012年10月、2014年8月和2016年9月先后3次对固定样地中木质残体进行调查,具体方法如下:对样地内所有木质残体鉴定树种(本研究将高度腐烂而无法鉴别的树种定义为未知树种)并对其进行生物量及碳储量的研究、逐株测量胸径、大头直径(或基径)、小头直径、中央直径和长度(或高度)、鉴定腐烂等级,同时标记序号。采用Harmon等在1996年制定的新标准以及我国学者闫恩荣等总结的分类标准,木质残体被分为两类:细木质残体(FWD)和粗木质残体(CWD),其中将1.0 cm≤直径<10 cm的木质残体定义为FWD,直径(通常指粗头部分)≥10 cm的死木质物为CWD。在一级分类的基础上,根据CWD在群落中所表现的状态和长度可进一步分为倒木(log)、枯立木(snag)、根桩(stump)、大枯枝(largebranch)。倒木是指倾斜度gt;45°,大头直径≥10cm,长度≥1m的死木质体;而倾斜度≤45°的则归为枯立木;与枯立木具有类似特征的,但长度小于1m则定义为根桩;而大头直径<10 cm,长度gt;1m的不含主干的死亡木则称为大枯枝。
1.2.2 木质残体生物量与碳储量计算
在每个样地中,以对角线的方式设置3个5mx5m的小样方,并且将小样方内的细木质残体称量后取3份混合样品,带回实验室置于85℃烘箱内烘干至质量恒定后根据含水率推算出细木质残体生物量。每次取样所设置的小样方均不重复。参考闫恩荣等粗木质残体的腐烂等级划分标准,对腐烂等级较轻(Ⅰ、Ⅱ和Ⅲ)的CWD,用钢锯截取5 cm厚的圆盘,采用排水法测定粗木质残体的体积,再将圆盘置于85℃烘箱内烘干至质量恒定;对于高腐烂等级(Ⅳ和Ⅴ)粗木质残体则直接用小刀取部分样品装满已知容积的铝盒,称其鲜质量,并带回实验室烘干,最后通过计算质量体积比求得木质残体的密度。最后计算木质残体的密度乘以对应的体积求得CWD的生物量。木质残体体积计算公式如下:
倒木的体积计算:
式中,V为体积/m3;dr为小头直径/cm;dR为大头直径/cm;L为倒木的长度Im;DBA为胸高断面积/m2;H为枯立木或根桩高度/m;f为形数(取0.43);A为根桩中端断面积/m2。
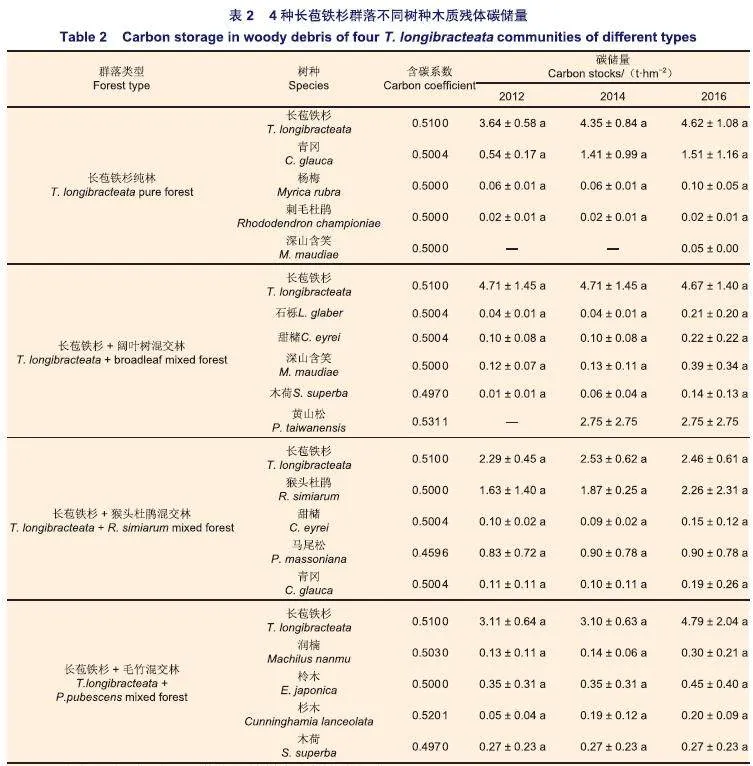
传统计算木质残体碳储量的方法是以生物量为基础,乘上相应的碳浓度(即含碳系数或含碳率)从而得到其碳储量。国内大多数研究采用0.5或者0. 45作为平均含碳率来推算森林的碳储量,由于不同树种,含碳系数也有所差异,若统一采用固定含碳率推算木质残体的碳储量会导致精度降低。因此,本研究参考《中国森林植被生物量和碳储量计算方法》及国家林业局印发的《造林项目碳汇计量监测指南》里各树种的含碳率,对于未进行碳含量检测的树种,参考相似树种的碳含量或采用平均值为0.5的碳含量来代替。本研究中不同树种木质残体采用的含碳率如表2所示。不同存在形式木质残体的含碳率基于本课题组前期调查测定结果。
2 结果和分析
2.1 4种长苞铁杉群落不同树种木质残体生物量特征
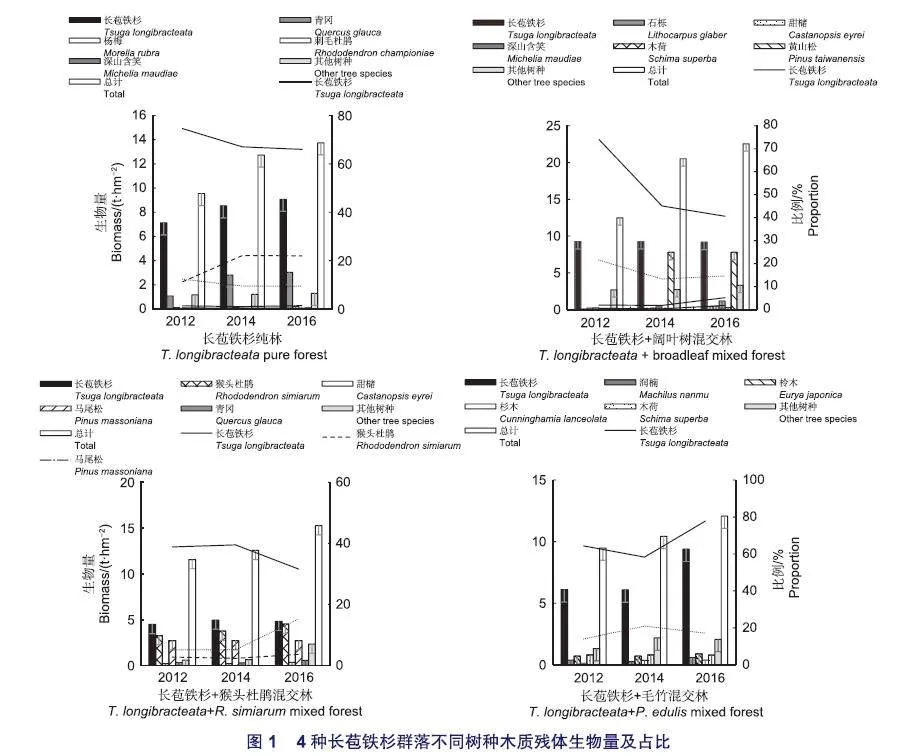
由图1可知,不同群落类型木质残体的树种组成不同,每种群落类型样地内的木质残体由11~16种植物组成,其中长苞铁杉的生物量均占比最大。长苞铁杉纯林中,长苞铁杉木质残体的生物最大,3个年份的占比均超过总生物量的65%,但随着调查年限的增加,其生物量比例有所下降,但青冈木质残体生物量有增加的趋势,其他树种木质残体生物量及比例变化较为稳定(图1);长苞铁杉+阔叶树混交林中,长苞铁杉木质残体的生物量变化不大,但其比例从2012年的74.06%降低到2016年的40.61%,其比例变化大的原因主要为此群落类型样地内黄山松(Pinus taiwanensis Hayata.)木质残体增加了7.77 t·hm-2,超过总生物量的1/4,同时,石栎(Lithocarpus glaber(Thunb.) Nakai.)、甜槠(Castanopsis eyrei(Champ.ex Benth.)Tutch.)、深山含笑(Michelia maudiae Dunn.)和木荷(Schimasuperba Gardn. et Champ.)的木质残体生物量也有所增加,使得长苞铁杉木质残体的比例下降(图1);长苞铁杉+猴头杜鹃混交林中,长苞铁杉木质残体占比最大,其次是猴头杜鹃和马尾松,三者比例之和超过80%,但马尾松木质残体生物量变化不大,比例逐年下降,这可能与群落内其他树种木质残体增多有关(图1);长苞铁杉+毛竹混交林中,长苞铁杉木质残体的比例最大,超过55%,在2016年的调查中,长苞铁杉的生物量较2014年增加3.31 t·hm-2,其比例增加19.46%(图1)。
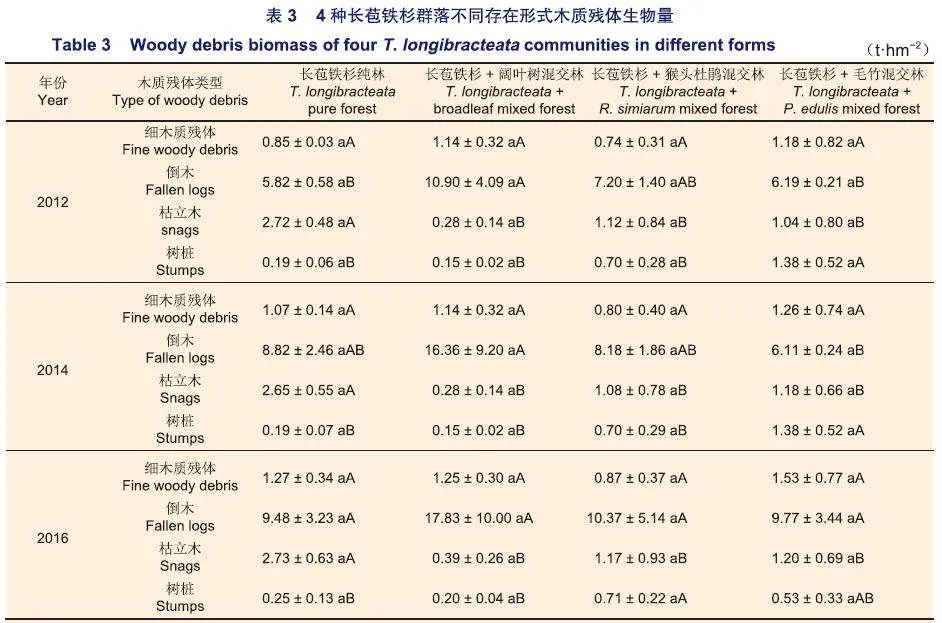
2.2 4种长苞铁杉群落不同存在形式木质残体生物量特征
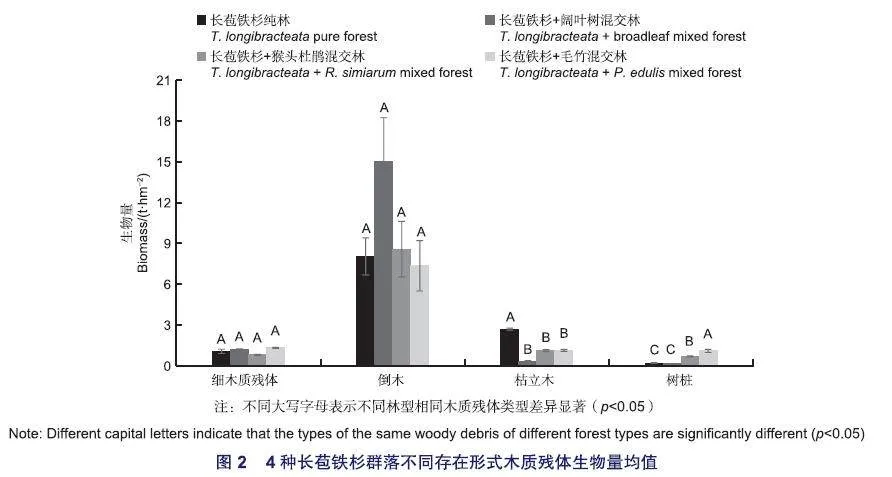
福建天宝岩国家级自然保护区4种长苞铁杉群落同一林型中相同木质残体类型各年间差异均不显著(pgt;0.05)(表3);不同木质残体类型生物量年际动态变化总体趋势为波动上升,2016年生物量最高;同一林型中倒木生物量最高(表3)。长苞铁杉纯林与长苞铁杉+猴头杜鹃混交林各组分生物量均是倒木gt;枯立木gt;细木质残体gt;树桩;长苞铁杉+阔叶树混交林则是倒木gt;细木质残体gt;枯立木gt;树桩;而长苞铁杉+毛竹混交林在2012年与2014年的调查中生物量是倒木gt;树桩gt;细木质残体gt;枯立木,而在2016年的调查中则是倒木gt;细木质残体gt;枯立木gt;树桩(表3)。长苞铁杉+毛竹混交林中由于受到的人为干扰较大,因此其木质残体的组成也存在一定的波动,而其他群落类型基本不受人为干扰,其木质残体生物量变化主要是由自然灾害、病虫害等因素引起的。
同一年份不同长苞铁杉群落中,细木质残体生物量各群落间差异均不显著(pgt;0.05)(表3);2012年长苞铁杉+阔叶树混交林与长苞铁杉纯林、长苞铁杉+毛竹混交林倒木生物量差异显著(p<0.05),长苞铁杉纯林枯立木生物量与长苞铁杉+阔叶树混交林差异极显著(p<0.01),与其他群落差异显著(p<0.05),长苞铁杉+毛竹混交林中树桩生物量与长苞铁杉纯林、长苞铁杉+阔叶树混交林差异极显著(p<0.01),与长苞铁杉+猴头杜鹃混交林差异显著(p<0.05)(表3);2014年长苞铁杉+阔叶树混交林与长苞铁杉+毛竹混交林倒木生物量差异显著(p<0.05),长苞铁杉纯林枯立木生物量与长苞铁杉+阔叶树混交林差异极显著(p<0.01),与其他两种群落差异显著(p<0.05),长苞铁杉+毛竹混交林中树桩生物量与长苞铁杉纯林、长苞铁杉+阔叶树混交林差异极显著(p<0.01),与长苞铁杉+猴头杜鹃混交林差异显著(p<0.05)(表3);2016年各群落倒木生物量差异均不显著(pgt;0.05),长苞铁杉纯林枯立木生物量与长苞铁杉+阔叶树混交林差异极显著(p<0.01),与其他两种群落差异显著(p<0.05),长苞铁杉+猴头杜鹃混交林树桩生物量与长苞铁杉纯林、长苞铁杉+阔叶树混交林差异显著(p<0.05)(表3)。
p从平均值来看,4种长苞铁杉群落类型中细木质残体和倒木差异不显著(pgt;0.05),长苞铁杉纯林枯立木生物量与其他群落差异显著(p<0.05),长苞铁杉纯林、长苞铁杉+阔叶树混交林与其他群落树桩生物量差异显著(p<0.05),长苞铁杉+猴头杜鹃混交林与长苞铁杉+毛竹混交林树桩生物量差异显著(p<0.05)(图2)。
2.3 4种长苞铁杉群落不同树种木质残体碳储量特征
4种长苞铁杉群落同一林型不同树种(除深山含笑与黄山松)各年份的碳储量差异均不显著(pgt;0.05)(表2);4种长苞铁杉群落均是长苞铁杉的碳储量最高,且随着年份的增长,其碳储量总体呈现增长趋势(表2)。长苞铁杉纯林中,长苞铁杉木质残体的碳储量最高,青冈木质残体碳储量呈增加的趋势,其他树种木质残体碳储量变化较为稳定;长苞铁杉+阔叶树混交林中,长苞铁杉木质残体的碳储量虽有增长但变化不大,2014年该群落内出现黄山松木质残体,其碳储量仅次于长苞铁杉;长苞铁杉+猴头杜鹃混交林中,长苞铁杉木质残体碳储量最高,其次是猴头杜鹃,其他树种木质残体碳储量变化不大;长苞铁杉+毛竹混交林中,长苞铁杉木质残体碳储量远高于其他树种,2016年,长苞铁杉的碳储量较2014年增加1.69 t·hm-2,其他树种木质残体碳储量变化较为稳定(表2)。
2.4 4种长苞铁杉群落不同存在形式木质残体碳储量特征
福建天宝岩4种长苞铁杉群落木质残体中均是倒木的碳储量最高,除长苞铁杉+毛竹混交林倒木碳储量是先减少后增加外,其他3种长苞铁杉群落倒木的碳储量均呈现上升趋势,且4种群落中,长苞铁杉+阔叶树混交林的倒木碳储量最高,在4.91~7.68 t·hm-2之间(表4);4种长苞铁杉群落细木质残体的碳储量总体均呈现增长趋势,其中长苞铁杉+毛竹混交林碳储量最高,在0.55~0.73 t·hm-2之间,长苞铁杉+猴头杜鹃混交林最少(表4);长苞铁杉纯林的枯立木碳储量最高,长苞铁杉+阔叶树混交林最少(表4);除长苞铁杉+毛竹混交林,其他3种长苞铁杉群落不同木质残体类型中树桩的碳储量最少,其中长苞铁杉+阔叶树混交林树桩的碳储量最少,其次是长苞铁杉纯林(表4);长苞铁杉纯林与长苞铁杉+猴头杜鹃混交林中各木质残体类型碳储量均呈现出倒木gt;枯立木gt;细木质残体gt;树桩(表4)。
同一年份不同长苞铁杉群落中,细木质残体碳储量各群落间差异均不显著(pgt;0.05)(表4);2012年长苞铁杉+阔叶树混交林倒木碳储量与长苞铁杉纯林、长苞铁杉+毛竹混交林显著差异(p<0.05),长苞铁杉纯林枯立木碳储量与长苞铁杉+阔叶树混交林差异极显著(p<0.01),与其余混交林枯立木碳储量均显著差异(p<0.05),长苞铁杉+毛竹混交林树桩碳储量与长苞铁杉纯林、长苞铁杉+阔叶树混交林差异极显著(p<0.01),与长苞铁杉+猴头杜鹃混交林差异显著(p<0.05)(表4);2014年长苞铁杉+阔叶树混交林倒木碳储量与长苞铁杉+毛竹混交林显著差异(p<0.05),长苞铁杉纯林枯立木碳储量与其他3种混交林显著极差异(p<0.01),长苞铁杉+毛竹混交林树桩碳储量与长苞铁杉纯林、长苞铁杉+阔叶树混交林差异极显著(p<0.01),与长苞铁杉+猴头杜鹃混交林差异显著(p<0.05)(表4);2016年长苞铁杉纯林与长苞铁杉+阔叶树混交林枯立木碳储量显著差异(p<0.05),与长苞铁杉+猴头杜鹃混交林、长苞铁杉+毛竹混交林差异极显著(p<0.01),长苞铁杉+猴头杜鹃混交林与长苞铁杉+阔叶树混交林树桩碳储量差异显著(p<0.05),各林型间倒木碳储量差异均不显著(pgt;0.05)(表4)。
从平均值来看,4种长苞铁杉群落类型中细木质残体和倒木碳储量差异不显著(pgt;0.05),长苞铁杉纯林枯立木碳储量与其他群落差异显著(p<0.05),长苞铁杉纯林、长苞铁杉+阔叶树混交林与其余混交林树桩碳储量差异显著(p<0.05),长苞铁杉+猴头杜鹃混交林与长苞铁杉+毛竹混交林树桩碳储量差异显著(p<0.05)(图3)。
3 讨论
森林生态系统中,木质残体的形成是一个极其复杂的过程,森林群落类型、树种组成、树种竞争、林龄、自然灾害以及人为干扰等都会使其生物量发生变化。福建天宝岩国家级自然保护区内不同树种木质残体生物量总体表现为逐年上升,总生物量为9.48~22.55 t·hm-2,与我国长白山针阔混交林中的CWD生物量(7.9~16.2 t·hm-2)相差不大,但高于同处亚热带的广州针叶林、针阔混交林和常绿阔叶林,远低于美国Olympic国家公园针叶林(537 t·hm-2)。纵观全球各种群落类型的木质残体生物量,天然针叶林的木质残体生物量范围为30~200 t·hm-2,世界针阔混交林木质残体生物量范围为5~50 t·hm-2,可见,天宝岩国家级自然保护区长苞铁杉林木质残体生物量远低于世界天然针叶林平均水平,但处于世界针阔混交林的范围内。产生这种差异受群落类型、所属地域、海拔和气候带等多种因素影响;另外,木质残体调查的直径界定标准不统一也是其原因之一。
木质残体生物量主要受输入量和分解量的双重作用,其中占主导作用的是输入量,天宝岩木质残体生物量以CWD占主体地位,且在CWD中生物量以枯立木与倒木为主,这与大部分研究保持一致。其中倒木的生物量含量明显高于其他类型的木质残体,这可能与群落内倒木的数量有关,这与何持卓研究岭南山地典型森林群落木质残体数量特征的结果是一致的。在3次调查中,长苞铁杉纯林中枯立木生物量均显著高于另外3种群落类型,这主要是由于长苞铁杉纯林中乔木树种基本以长苞铁杉为主,但其平均胸径远低于其他群落类型长苞铁杉,而且该群落类型乔木层树种密度较大,使得群落内长苞铁杉种内竞争加剧,一些植物在竞争中死亡,从而增加枯立木的生物量。2012年和2014年长苞铁杉+毛竹混交林中根桩生物量显著高于另外3种群落类型,这是因为长苞铁杉+毛竹混交林中人为干扰较严重,人为砍伐加大了根桩的生物量。
国内外大多数学者多采用0.45或0.5作为林分的平均含碳率进行森林生态系统碳储量的研究,这可能会产生较大的系统误差,增加了研究结果的不确定性。我国学者对不同森林类型树种作了一些测定,如唐旭利等对鼎湖山南亚热带季风常绿阔叶林的树种和器官进行了测定;闫平等对红松(Pinus koraiensis Sieb. et Zucc.)、色木槭(Acer mono Maxim.)、椴树(Tilia tuanSzyszyl.)和山杨(Populus davidiana Dode.)各器官的含碳率进行了研究。因此在本研究中对不同树种木质残体采用了与其相同或相近树种含碳率,以提高研究的准确性,增加了研究结果的可信度。目前,对于森林生态系统的碳平衡研究主要侧重于土壤和活立木的碳储量,而对木质残体的碳储量研究相对较少。但据估计,木质残体碳储量占森林地表碳储量的14%~17%,是植被碳库与土壤碳库之间重要的纽带。本研究中,4种长苞铁杉群落木质残体中均是倒木的碳储量最高,这与赵嘉诚和李海奎研究广东省森林死木碳库特征结果一致,而吉林省死木碳库中倒木碳储量小于枯立木碳储量,这可能与吉林省气候条件有关。
木质残体生物量与碳储量受气候、海拔、腐烂等级、树种、人为原因等各种因素的影响,本研究中仅研究了不同树种和不同形式的木质残体,并未考虑到不同腐烂等级木质残体生物量与碳储量特征,这是本研究的不足之处,在今后的研究应注意这一点。
4 结论
福建天宝岩4种长苞铁杉群落不同树种木质残体均是长苞铁杉生物量占比最大;在长苞铁杉+阔叶树混交林的3次调查中,长苞铁杉木质残体的生物量变化不大,但其比例降低了33.45%,其主要原因为此群落类型样地内黄山松木质残体增加了7.77 t·hm-2,占比超过总生物量的1/4。同一林型中不同形式木质残体倒木生物量最高,且在3次调查中长苞铁杉+阔叶树混交林内倒木生物量均大于其他群落内的倒木生物量,在10.90~17.83t·hm-2之间;长苞铁杉纯林内枯立木生物量与其他群落内枯立木生物量差异显著。4种长苞铁杉群落内不同树种木质残体中均是长苞铁杉的碳储量最多,且随着年份的增长,其碳储量总体呈现增长趋势。不同形式木质残体中均是倒木的碳储量最高,其中长苞铁杉+阔叶树混交林倒木碳储量高于其他3种群落,在4.91~7.68 t·hm-2之间;3次调查中,长苞铁杉纯林的枯立木碳储量与其他3种混交林差异显著。
(责任编辑:崔贝)
基金项目:国家自然科学基金资助项目(31370624)、福建省地质测绘院横向课题(KH220035A)
猜你喜欢
今日农业(2020年19期)2020-12-14 14:16:52
安徽农学通报(2020年7期)2020-05-26 01:58:28
甘肃科技(2020年21期)2020-04-13 00:33:46
安徽农学通报(2017年1期)2017-02-15 18:21:10
大经贸(2016年11期)2017-01-06 21:39:07
现代园艺(2016年17期)2016-10-17 06:54:29
电脑知识与技术(2016年5期)2016-04-14 11:15:42
中国科技信息(2015年2期)2015-11-16 08:18:32
绿色科技(2015年6期)2015-08-05 18:47:03
湖北农业科学(2014年3期)2014-07-21 10:48:15
