
长梗柳带叶插穗不定根形成中内源激素含量变化
2023-12-21 03:09王中煊熊远芳陈世品冯丽贞
生物学杂志 2023年6期
蔡 琦,龚 娜,王中煊,熊远芳,陈世品,冯丽贞
(福建农林大学 林学院,福州 350002)
LED植物光源系统具有绿色环保、操作简便、智能调控等优点,能大大提高植物培育效率[1],既可生成多种波长的单色光源,与植物光合和光形态建成的光谱相一致,还可将不同波长LED光组合成为复合光,高效促进植物的形态建成以及对生长发育的调控[2]。红光和蓝光是植物吸收、利用最多且最重要的有效光源[3]。相关研究表明单色红光处理能显著促进IAA、GA和ABA 的合成,有利于提高植物的生根率、不定根数以及平均根长[4-6]。
长梗柳 (Salixdunnii) 为杨柳科 (Salicaceae) 柳属 (Salix)植物,是我国华东和华南地区溪流、江河、湖泊等水系旁的护岸植物[7]。长梗柳雌雄异株,种子园建设技术尚不成熟,扦插是其主要繁殖手段。为进一步提高其扦插生根效率和质量,以红光处理带叶插穗,探索其不定根发生机制,可为长梗柳的推广栽培提供有效指导。
1 材料与方法
1.1 材料
2021年9月在福建农林大学林学院种苗繁育实验室开展相关研究工作。上午8点采集长梗柳长势良好、无病虫害、向阳的当年生半木质化嫩枝,用浅水浸泡带回实验室。
1.2 方法
1.2.1 扦插方法
选择带顶芽枝条作为插穗,制穗长度约10 cm,留2片半叶,插穗下端45°斜切且尽量靠近叶节处。将穗条放入80%多菌灵800倍溶液中浸泡消毒10 min后,蒸馏水冲洗3次,将稍微晾干的穗条垂直插入泥炭土、蛭石、细沙(2∶2∶1)混合的穴盘中,扦插深度约为插穗长度1/3。将穴盘放入密封箱中,其规格为长51.5 cm、宽37 cm、高30 cm,侧面开一个直径5 cm圆孔,贴上2层纱布,密封箱盖为透光率92%的亚克力板。密封箱再放入自制智能扦插箱中进行红光处理实验。参数设定:温度(30±2) ℃,湿度为90%,光强为50 μmol/(m2·s)16 h光照/8 h黑暗。设置红光(波长660 nm),3个重复。
1.2.2 组织结构观察
长梗柳插穗根原基发育分为3个时期:Ⅰ期为扦插起始时期、Ⅱ期为根原基分化期、Ⅲ期为根原基成熟期。从处理开始至根原基成熟约7 d,其中,Ⅰ期为实验开始当天,Ⅱ期、Ⅲ期分别对应第3天与第7天。对插穗基部0.5~1.0 cm茎段取样,制作解剖学切片进行观察[8]。
1.2.3 内源激素提取与测定
分别于Ⅰ、Ⅱ、Ⅲ期取样。取样部位为插穗顶芽和基部,每个取样3个重复。参考吴颂如等[9]的酶联免疫吸附法测定内源激素含量,ELISA检测试剂盒购自福州麦力生物科技有限公司。
2 结果与分析
2.1 不定根的生长与发育
长梗柳扦插,不定根从皮部部位伸出,形态上属于皮部生根类型。长梗柳扦插生根插穗基部形态变化如图1所示。扦插3 d,50%插穗顶芽开始生长,多数插穗基部有明显膨大和长条状白斑,白斑下部皮孔张开;扦插7 d,80%插穗顶芽逐渐长大,部分插穗侧芽也开始生长,大部分插穗基部有明显不定根;插穗未出现愈伤组织,因此,推断长梗柳扦插生根方式为皮部生根型。

(a)插穗原始基部形态 (Ⅰ期) ;(b)扦插3 d插穗基部出现白斑,皮孔开始张开 (Ⅱ期) ;(c)扦插7 d,插穗基本生根 (Ⅲ期)。
如图2(a)所示,通过连续横切切片发现,长梗柳插穗组织结构由髓、木质部、维管形成层、韧皮部、皮层及表皮等构成,且未发现潜伏型根原基存在。如图2(b),在Ⅱ期即表型中发现白色絮状物质,可观察到皮部处有大量薄壁细胞,这些薄壁细胞是由皮层薄壁细胞(Pc)分化而来,此时根原基开始诱导,且发现不定根原基由皮层和韧皮部薄壁细胞诱导发育而来。如图2(c)和(d),Ⅲ期即表型中皮部白色絮状物质分化形成不定根原基(Arp),不定根原基穿过皮层伸出体外,形成不定根(Ar)。
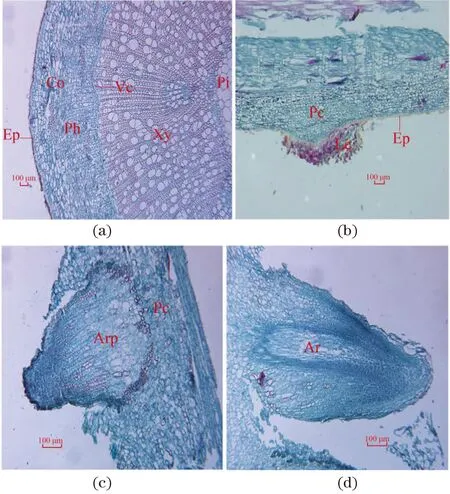
(a)穗条Ⅰ期;(b)Ⅱ期;(c)和(d)Ⅲ期;(a)横切结构,(b)~(d)纵切结构。Ep:表皮;Co:皮层;Ph:韧皮部;Vc:维管形成层;Xy:木质部;P:髓;Le:皮孔;Pc:薄壁细胞;Arp:不定根原基;Ar:不定根。
2.2 不定根发生过程中内源激素含量
2.2.1 IAA含量
如图3所示,长梗柳带叶插穗内源IAA含量在不定根形成过程中芽内含量呈现显著性上升(P<0.05)后继续缓慢上升趋势,且Ⅲ期含量最高,为739.52 ng/g FW,比Ⅰ期提高19.27%;而皮内IAA含量则表现为先显著性上升再显著性下降(P<0.05),且Ⅲ期IAA含量下降后低于Ⅰ期,下降16.01%(P<0.05)。在不定根形成过程中,不定根原基诱导和不定根形成过程是消耗大量IAA的过程,从而导致皮内IAA急剧下降,比Ⅱ期下降41.63%(P<0.05);随着根原基形成,插穗到达发育阶段,消耗IAA的同时自我合成新的IAA,因此,芽内IAA含量呈现上升趋势。
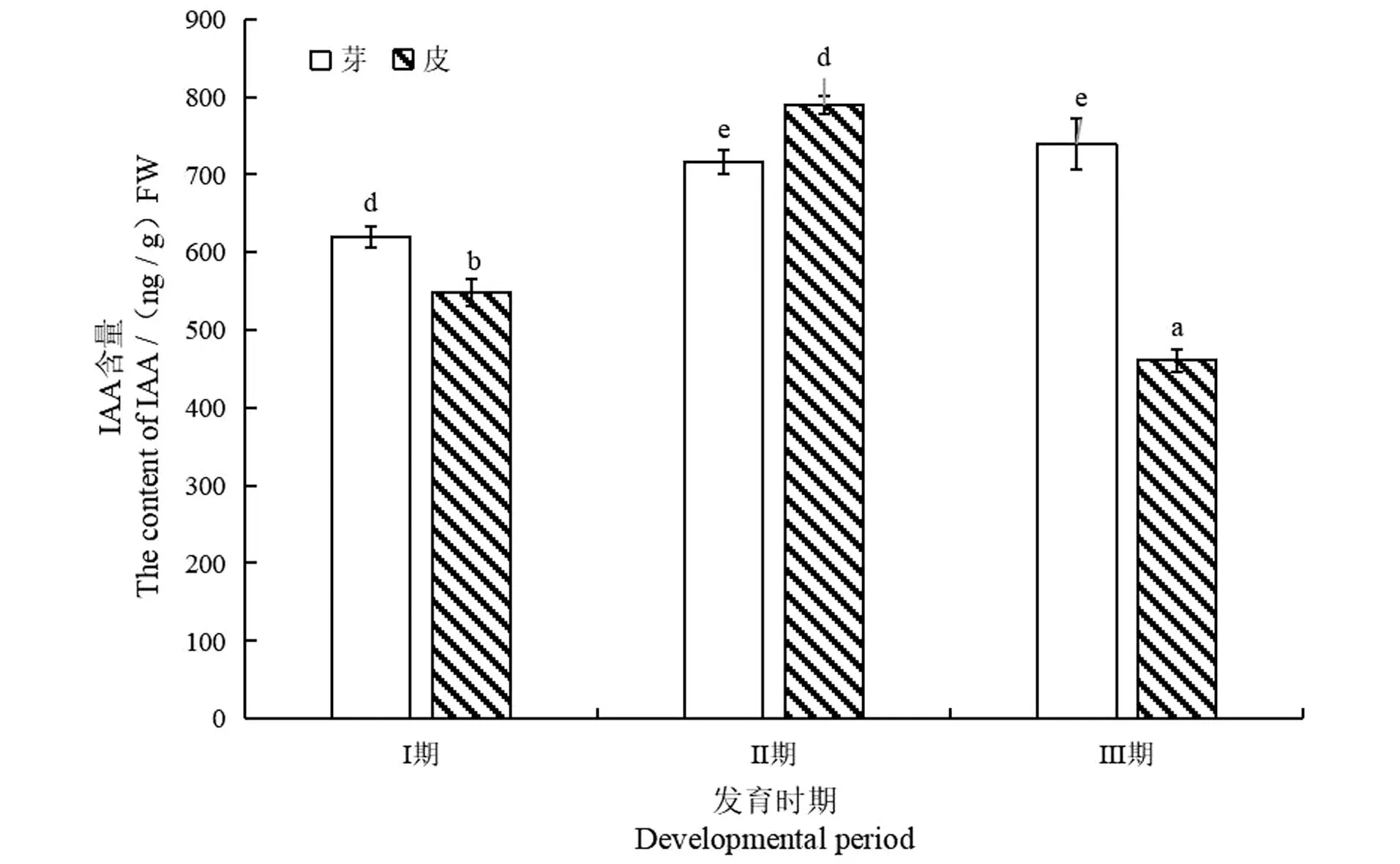
相同字母间表示差异不显著,不同字母间表示差异显著。
2.2.2 ABA含量
如图4所示,长梗柳扦插生根芽内ABA含量先显著性降低再缓慢下降 (P<0.05),比Ⅰ期降低56.95%,其中Ⅲ、Ⅱ期ABA含量均低于Ⅰ期。Ⅱ期和Ⅲ期皮内ABA含量比较高,均大于600 ng/g FW,Ⅱ期比Ⅰ期提高57.01%,Ⅲ期比Ⅱ期增加9.77%(P<0.05)。初期插穗损伤促进ABA积累,随着不定根发育,ABA积累下降和消耗上升并存,Ⅲ期ABA总含量低于Ⅰ期和Ⅱ期,说明低水平ABA有利于不定根形成与表达。
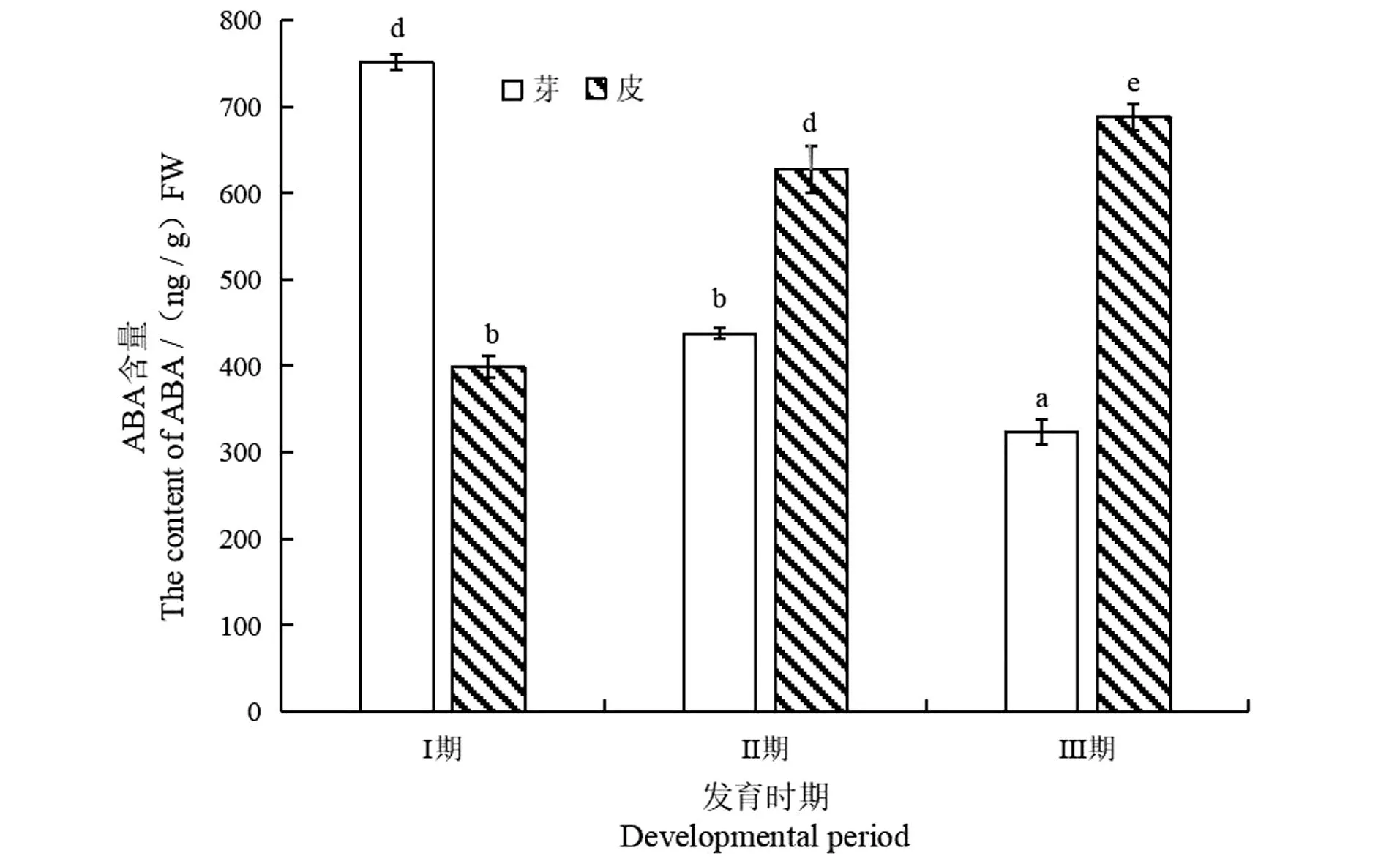
相同字母间表示差异不显著,不同字母间表示差异显著。
2.2.3 BR含量
如图5所示,长梗柳扦插生根芽内BR含量先显著上升再缓慢下降,但仍高于Ⅰ期18.92%(P<0.05);长梗柳扦插生根皮内BR含量先缓慢上升再显著下降,最后反而比Ⅰ期下降29.11%(P<0.05)。随着大量薄壁细胞形成快速积累,达到峰值(882.39 ng/g FW),为不定根形成奠定了基础。随着不定根形成,BR含量呈现急剧下降趋势。

相同字母间表示差异不显著,不同字母间表示差异显著。
2.2.4 GA含量
如图6所示,长梗柳扦插生根芽内GA含量表现为持续缓慢上升(P<0.05),比Ⅰ期增加3.41%;皮内GA含量均呈现下降再缓慢回升趋势,比Ⅰ期下降27.76%(P<0.05)。表明大量薄壁细胞生成是消耗GA含量的过程,当不定根形成,插穗恢复合成GA的能力,因此Ⅲ期GA含量呈现缓慢回升趋势,可见GA对长梗柳带叶插穗不定根形成起着重要作用。
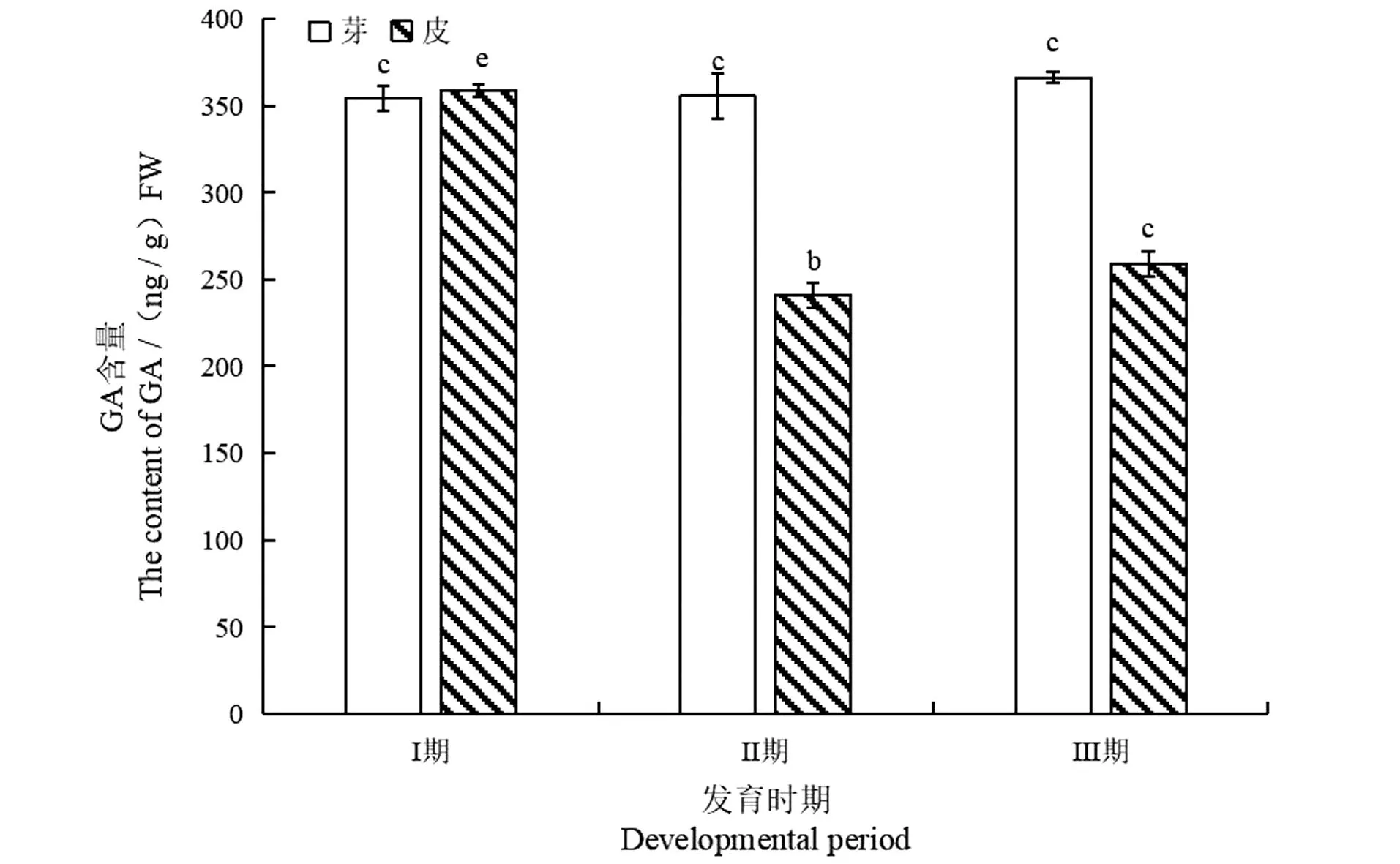
相同字母间表示差异不显著,不同字母间表示差异显著。
2.3 BR bHLH基因的表达量
bHLH类基因中,Sadunf02G0184300在P-T1时期表达量逐渐上调,在B-T2时期和L-T1时期表达量显著性下调;Sadunf14G0114600在P-T0时期、P-T1时期、P-T2时期表达量较高,B-T1时期、B-T2时期和L-T1时期、L-T2时期表达量显著上调;Sadunf04G0094100在P-T0时期、P-T1时期、P-T2时期表达量逐渐下降,在B-T0时期、B-T1时期、B-T2时期表达量逐渐升高,其他时期几乎不表达;Sadunf12G0052400在P-T0时期、P-T1时期、P-T2时期表达量逐渐上调,其他时期几乎不表达;Sadunf15G0056200在P-T2时期表达量显著上调,其他时期几乎不表达,见图7。
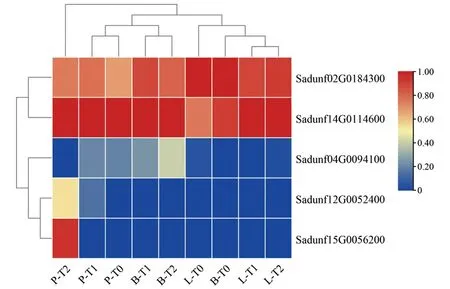
P-T0、B-T0、L-T0分别为穗条皮部、芽和叶的Ⅰ期;P-T1、B-T1、L-T1分别为穗条皮部、芽和叶的Ⅱ期;P-T2、B-T2、L-T2分别为穗条皮部、芽和叶的Ⅲ期。
3 讨论与结论
植物根系由一个初生根建成,它由胚胎发生过程中形成的胚根发育而来,胚后初生根可以分枝并形成侧根[10]。胚后根可以由茎、叶或其他营养性非根器官发育而来,也可以是对胁迫的应激反应而生成,这样的根被称为不定根(Adventitious root,Ar)[11]。植物扦插能否成功的关键因素就是插穗能否产生不定根,不定根形成是一个复杂的、多层次调控的发育过程。在插穗中,Ar可以从茎的维管束附近的中柱鞘组织细胞分化而形成,或从损伤后分化的愈伤组织中发育分化出根原基而形成,因此,将前者称为皮部生根类型,后者则称为愈伤生根类型[12-13]。本研究通过长梗柳扦插生根插穗外部形态学观察,发现插穗均是皮部开裂长出白色絮状物质,因此,认为长梗柳扦插生根方式为皮部生根类型。
通过石蜡切片观察发现,长梗柳带叶插穗茎的显微结构与形态观察结果一致。前人研究认为不定根形成分为3个时期,诱导期插穗会产生一系列生化反应[14];起始期细胞开始分裂并产生根原基[15];表达期是插穗根原基继续分化生长,形成不定根并突出皮孔[16]。因此,本研究将长梗柳扦插不定根形成分为3个时期:Ⅰ期即插穗原始时期;Ⅱ期扦插3 d后,皮孔上开始出现白色絮状物,即根原基开始诱导分化;Ⅲ期扦插7 d后,根原基分化及不定根形成。根原基可分为潜伏根原基和诱生根原基[17-19]。诱生根原基指离体前穗条中不存在根原基,插穗离体扦插后因应激损伤或非生物胁迫,而由髓射线、韧皮射线、维管形成层以及韧皮薄壁组织等部位细胞诱导分化产生,或由插穗基部切口的愈伤组织产生根原基[20]。本研究通过显微结构观察,在扦插前穗条茎中并未发现根原始体,而是后期由韧皮部及皮层薄壁细胞诱导分化形成,因此长梗柳带叶插穗根原基属于诱生根原基。
内源激素在植株扦插生根中起重要作用。本试验中,IAA含量高峰与根原基出现的高峰相同,是促进不定根发生的主要激素[21]。ABA会抑制植物生长发育,主要是作为拮抗剂通过和其他内源激素(IAA、CTK、GA、BR、乙烯)间相互作用来影响不定根形成发育[22]。研究发现,ABA总含量在扦插初期较高,在生根过程中持续下降,有利于插穗内淀粉水解为糖,促进根原基形成和插穗生根[23]。BR是植物中的甾醇类激素,在植物光形态建成、叶片衰老、生殖发育、茎和根中的细胞分裂等发育过程中起重要作用。本研究结果表明,植株可通过影响体内IAA极性运输来促进诱导不定根形成,而低浓度BR能诱导侧根形成,且部分是通过影响生长素极性运输来实现,因此认为BR与生长素具有协同作用[24]。在基因的表达量分析中,Sadunf14G0114600和Sadunf02G0184300在Ⅱ期和Ⅲ期中表达量较高。因此,可能对生根过程有着重要作用。夏季长梗柳萌发生根约15 d,秋冬季约1个月,而在红光培养箱中3~7 d就可萌发生根。因此,分析认为BR升高可能是由于红光作用。不同树种GA研究结果不同。本研究长梗柳扦插与陈凌艳等[25]在杂种杜鹃 (Rhododendronhybrida) 组培中发现高浓度GA会抑制不定根形成的研究结果一致。而赵云龙等[26]在糙叶杜鹃 (Rhododendronscabrifolium) 扦插生根过程中分析发现GA含量积累,对插穗愈伤组织的诱导和不定根发生具有正相关影响。
综上所述,通过红光处理下长梗柳扦插生根类型、生根数量、内源激素水平的影响及基因表达量分析研究表明,长梗柳带叶插穗不定根发育是由不同内源激素调控的复杂发育过程。
猜你喜欢
作物研究(2022年1期)2022-11-27
作物学报(2022年10期)2022-07-21
现代农业研究(2021年10期)2021-11-02
计算机应用(2018年10期)2018-11-22
天然产物研究与开发(2018年4期)2018-05-07
华南师范大学学报(自然科学版)(2017年4期)2017-09-11
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19
上海农业学报(2016年2期)2016-10-27
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
动物营养学报(2015年10期)2015-12-01
