
黔南高等植物多样性与分布特征
2023-12-15 12:15:14吴天柔安明态
中国野生植物资源 2023年11期
吴天柔,安明态*,刘 锋
(1.贵州大学 林学院,贵州 贵阳 550025;2.贵州大学 生物多样性与自然保护研究中心,贵州 贵阳 550025;3.贵州大学 生命科学学院,贵州 贵阳 550025)
高等植物又称茎叶体植物,结构上具有胚胎构造,形态上一般具有根、茎、叶的分化,主要由非维管植物和维管植物两大类组成[1]。其中,维管植物包含石松与蕨类、种子植物,非维管植物包含苔藓植物。全球拥有30 余万高等植物,相比低等植物,其对环境有更高的适应性,广泛分布于高山、海洋、沼泽、沙漠等地,种类繁多,数量庞大,在植物资源中占据主体地位。植物资源是一个国家的重要战略资源,物种多样性及遗传基因的演化事关国家生态、环境安全发展,植物物种多样性的分布模式已被广泛应用于全球生物多样性保护优先级的设置。中国物种多样性仅次于巴西,拥有37 699 种高等植物,是北半球高等植物资源最丰富的国家,约占全球物种8%~12%[2]。高等植物主要分布于我国95°E~110°E 和20°N~34°N 之间的西南地区[3],位于该区域范围内的中国西南部地区的贵州省高等植物资源丰富多样。据统计,贵州省拥有苔藓植物94 科366 属1 643 种[4],蕨类植物53 科151 属770种[5],种子植物215科1 661属7 762种[6]。
黔南位于贵州省中南部地区,地处云贵高原东南边缘的倾斜地带,喀斯特地貌与常态地貌镶嵌交错,为中国西南喀斯特地貌高度聚集的典型分布区之一。境内山脉水系交错分布,苗岭山脉横贯其中形成长江和珠江两大流域的分水岭,喀斯特生态系统特殊的二元水文结构以及特殊复杂的地形地势形成独具特色的小区域气候类型[7]。植物群落生境异质性高,森林结构完整且典型,保存了丰富的珍稀濒危、特有以及古老孑遗植物,是珍稀濒危物种高度聚集地[8],也是全省高等植物丰富性、多样性、特有性最突出的地区之一。物种分布受其地理位置、形成历史、海拔、生境类型等因素影响较大[9],发展趋势不稳定且难以预测,构建适宜的生物多样性评价指标体系、监测评估生物多样性状况及变化趋势,已被纳入我国生物多样性战略计划[10]。黔南地区高等植物多样性、物种组成与数量、分布等资源特征的系统性研究未有报到,限制了该地区对高等植物多样性保护和利用的科学评价与管理决策。在全球生物多样性成为生态学研究主旋律且生物多样性丧失日益严重的大背景下,开展黔南州高等植物物种多样性与分布特征研究是科学评价该区植物资源特色及其生态功能地位的重要基础和必要前提。在大量查阅黔南地区高等植物资料并结合多年积累的调查数据基础上,通过地区数据查阅整理摸清黔南州高等植物资源情况,分析植物多样性与地理分布格局特点,探究该区域物种多样性、分布特征与环境因子的相互关系,以期为该地区植物资源的合理利用和生物多样性保护提供科学依据。
1 材料与方法
1.1 研究区概况
黔南布依族苗族自治州位于贵州省中部至南部地区,与广西河池市相邻,整体呈“凸”字形,地理坐标为106°12′E~108°18′E,25°04′N~27°29′N,东西宽约207.9 km,南北长约269.4 km,总面积2 620 000 hm2,占贵州省国土面积的15%[11]。全州共2 个县级市、1 个自治县、9 个县共计12 个县级行政单元。全州处于云贵高原东南部边缘的斜坡地带,整体上呈西高东低的倾斜面,平均海拔800~1 000 m。该州地形以山地、高原为主,多为喀斯特地貌和侵蚀地貌区,生境异质性强,生态环境脆弱[12]。区域内气候为云贵高原、亚热带季风气候地区,属中亚热带季风湿润气候类型,水热充沛,雨热同季。由于其独特的地理环境及气候条件,使得该地区具有极为丰富的生物多样性,其中荔波县和三都县部分保护区及乡镇属于我国南岭生物多样性优先保护区范围,罗甸县部分地区为桂西黔南石灰岩生物多样性保护优先区域(https://www.mee.gov.cn/gkml/hbb/bgg/201601/t20160105_321061.htm)。
1.2 植物数据来源
主要参考中国数字植物标本馆收录植物标本(https://www.cvh.ac.cn/)、贵州大学林学院植物标本库的标本数据以及《黔南布依族苗族自治州林木种质资源》、《贵州省维管植物名录》、《贵州省非维管植物名录》、《贵州省生物多样性现状》等相关资料[13-18],收集近几年在黔南州发表的新种及新记录种(科考集及专著资料未记录种),并对物种分布较少、调查相对薄弱的县域进行补充调查。依据《中国植物志》等[4,5,19,20]对物种名称与分类进行校对,剔除同物异名或同名异物的物种、人工栽培种,将变种、变型等种下等级设计为独立的分类单元,分布信息统一到县级单元,统计物种分布、生活型及海拔等数据。在完整统计植物名录(剔除栽培种)的基础上,对高等植物物种多样性和分布特征进行研究分析。
1.3 环境因子数据来源
使用气候、地形、人为干扰等3 个变量的19 个环境因子与黔南高等植物进行相关性分析,其中气候变量来源于世界气候数据库(https://www.worldclim.org/)的生物气候变量图层(bio1-bio19);地形变量来源于地理监测云平台(http://www.dsac.cn/);人为干扰变量中人口密度来自NASA 的社会经济数据和应用中心(https://sedac.ciesin.columbia.edu/)。
1.4 数据分析
1.4.1 相似性分析
通过R 语言的Vegan 包完成Jaccard 函数计算,获得各研究单元物种相似性系数矩阵及聚类图,其表达式[21]:S 为相似性系数;A,B 分别为两地物种数;C 为两地共有物种数。
1.4.2 分化度分析
植物科所含属越多,属所含物种数越多,即(属数/科数)与(种数/属数)两者比值之和越大,视为植物种系分化度越高;反之越小分化度越低,其表达式[22]:
SD 为物种分化度系数;n1为科数;n2为属数;n3为种数。
1.4.3 热点分布区识别
物种热点分布区采用Dobson 筛除算法进行识别[23],先筛选出植物最丰富的地区,将该地区内分布所有物种在剩余地区中筛除,在剩余区域中再次筛选出物种最丰富的区域,同上步骤将该次级区域内所有分布的植物在其余区域中筛除。以此类推,获得降序物种丰富区排序,按序统计各区物种数与总物种数占比,超过总数80%所含区域设为热点分布区。
1.4.4 物种空间自相关
通过GeoDa软件利用空间自相关模型计算丰富度的空间自相关系数(Moran’s I 和Moran’s Ii)作为基本的度量指标,检验各县(市)空间上是否具有高高相邻(H-H)、低低相邻(L-L)或高低相邻(H-L)的聚集类型。为减小物种数受区域面积大小的影响,进行数据处理:C = S/lnA(C为丰富度标准值,S为区域物种数,A 为区域面积),并通过ArcGIS 10.2 完成图的绘制。空间自相关指数计算式[24]:
wij为权重,i和j在本研究中为统计单元区域,当空间点i和j为邻接关系时,wij= 1;否则wij= 0。xi、xj分别为空间点i和j的物种数值,n为研究区分类单元数。Moran’s I 指数的取值在[-1,1]之间,越接近1,空间相关性越明显,I= 0 表示各区域之间没有空间相关性,I< 0表示区域之间有负空间自相关。
1.4.5 植物垂直分布
通过地理监测云平台下载地形变量(EDM 高程),运用ArcGIS 10.2 软件提取黔南地区高程数据。以200 m 为一个海拔段,进行重分类并计算各海拔区间像元与总像元数做比值计算,按比例乘以黔南州面积,获得200~1 970 m 区间200 m 为一个梯度的9 个海拔区间面积Ⅰ、Ⅱ、Ⅲ、…、Ⅸ。在Origin 2021 软件中绘制出各段海拔面积与物种分布变化图。
1.4.6 环境因子的相关性分析
通过ArcGIS 10.2软件提取19个环境因子信息,运用SPSS 26.0 软件进行环境因子之间的相关性分析,筛除相关系数绝对值大于0.9 的环境因子消除因子间的自相关性。以物种丰富度、分化度、密度3 个指标作为物种多样性的衡量指标与筛选环境因子在Canoco 5.0 软件中进行冗余分析(Redundancy analysis, RDA)。
2 结果与分析
2.1 物种多样性及相似性
据统计,黔南州分布高等植物326 科1 664 属6 487 种。其中,苔藓类植物97 科239 属908 种,分别占总科、属、种的30%、14%、14%;石松与蕨类植物34 科123 属689 种,分别占总科、属、种的10%、7%、11%;种子植物195 科1 302 属4 890 种(裸子植物8 科31 属82 种;被子植物187 科1 271 属4 808种),分别占总科、属、种的60%、78%、75%,科、属、种占主要优势。整体上以科和属为统计单元,禾本科(Poaceae)86 属201 种、蔷薇科(Rosaceae)35 属292 种、豆科(Fabaceae)81 属259 种为主要优势科,物种数占总数11.59%;以杜鹃花属(RhododendronL.)93 种、气藓属(AerobryumDoz. et Molk.)87 种、悬钩子属(RubusL.)83 种为主要优势属,物种数占总数4.1%。
以县为统计单元计算物种相似性系数矩阵并构建相似性聚类图,结果表明(图1),黔南州物种可分为6大聚类组,长顺县与平塘县类聚;瓮安县与福泉市类聚;龙里县与惠水县类聚;都匀市与贵定县类聚;独山县与三都县类聚;荔波县与罗甸县类聚,且物种相似性最高。不同地理区域物种间的相似性系数存在一定差异,相距较远地区物种相似性较低。

图1 黔南州高等植物行政单元物种相似性聚类Fig.1 Species similarity clustering of administrative units of higher plants in Qiannan Prefecture
2.2 植物分化度与热点分布区识别
分化度计算结果表明(图2),黔南高等植物科、属、种的组成与SD 值分别为:都匀县222 科973属2 670 种,SD 值为7.13;罗甸县225 科979 属2 683种,SD 值为7.09;荔波县229 科939 属2 693 种,SD值为6.97;独山县221 科939 属2 520 种,SD 值为6.93;三都县229 科933 属2 445 种,SD 值为6.69;贵定县220 科819 属2 114 种,SD 值为6.3;惠水县210科737 属1 968 种,SD 值为6.18;龙里县198 科661属1 735 种,SD 值为5.96;福泉市196 科661 属1 633种,SD 值为5.84;平塘县198 科682 属1 621 种,SD值为5.82;长顺县196 科675 属1 575 种,SD 值为5.78;瓮安县263 科790 属1 953 种,SD 值为5.48。研究区域中都匀市、罗甸县、荔波县等地物种SD 值较高,物种存在较分化的特性,瓮安县SD 值最低,物种分化性质较弱。

图2 黔南州高等植物物种丰富度与分化度Fig.2 Species richness and differentiation of higher plants in Qiannan Prefecture
通过Dobson 筛除算法逐级筛除分布物种,当包含物种数累计超过80%时共筛选出4 个分布区,即荔波县、罗甸县、都匀市、独山县拟定为黔南州高等植物分布热点地区,同时也是黔南州种系高分化区。物种的分化度与丰富性之间存在密切联系,即物种分化度越高的县域具有丰富的物种多样性。
2.3 物种空间自相关分析
高等植物不同分布区域与相邻区域物种间的依赖水平与相关程度的空间分布结果表明:Global Moran’s I = 0.115,物种丰富度空间关系呈正相关,物种丰富度的空间分布表现出显著的空间相关性和聚集性(图3)。三都县与相邻县物种丰富度呈H-H 型聚集,相邻区域物种丰富度受该地区物种影响,形成黔南州东南地区以三都县为中心的物种丰富度高值聚集区域。罗甸县、贵定县均与其相邻地区形成H-L 型聚集,两者相邻县区为低值区,罗甸县与贵定县为丰富度高值区。惠水县与相邻县区为L-L 型聚集,为黔南州物种丰富度低值区。显著性检验结果表明有8 区域不显著,这些区域的丰富度值在空间上为随机分布。
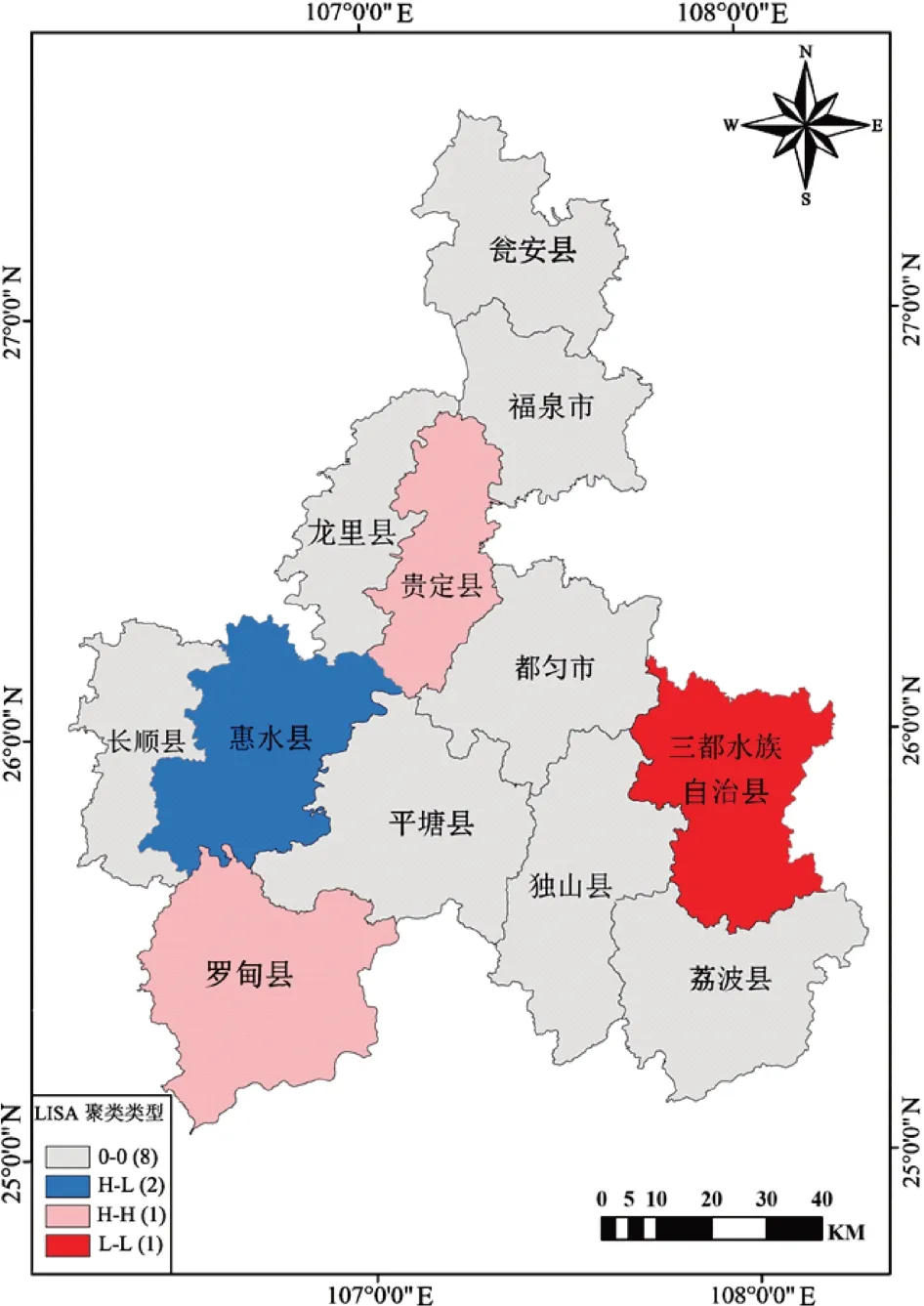
图3 黔南州高等植物局部莫兰聚类图Fig.3 Local Moran clustering diagram
2.4 物种海拔梯度变化
黔南高等植物物种随海拔梯度变化呈现出“单峰型”的分布特征(图4),物种丰富度高值出现于800 ~ 1 200 m 的海拔段,共分布物种4 593 种,占总数的70.8%。其中乔木1 009 种、灌木2 015 种、草本3 463种,将木本植物划分常绿、落叶植物,草本植物分为一、二年生草本植物与多年生草本植物,随海拔梯度变化均呈先上升后下降的变化。常绿植物与一、二年生草本植物物种丰富度减少幅度较大,落叶植物与多年生草本下降幅度相对较小。密度曲线与物种海拔梯度变化表现出较大差别(图4-a),曲线在1 600 m 海拔区间前较为平缓无明显变化,海拔1 600 m 以上出现急剧上升趋势,不考虑面积影响下高海拔地区(> 1 600 m),物种多样性为聚集分布。
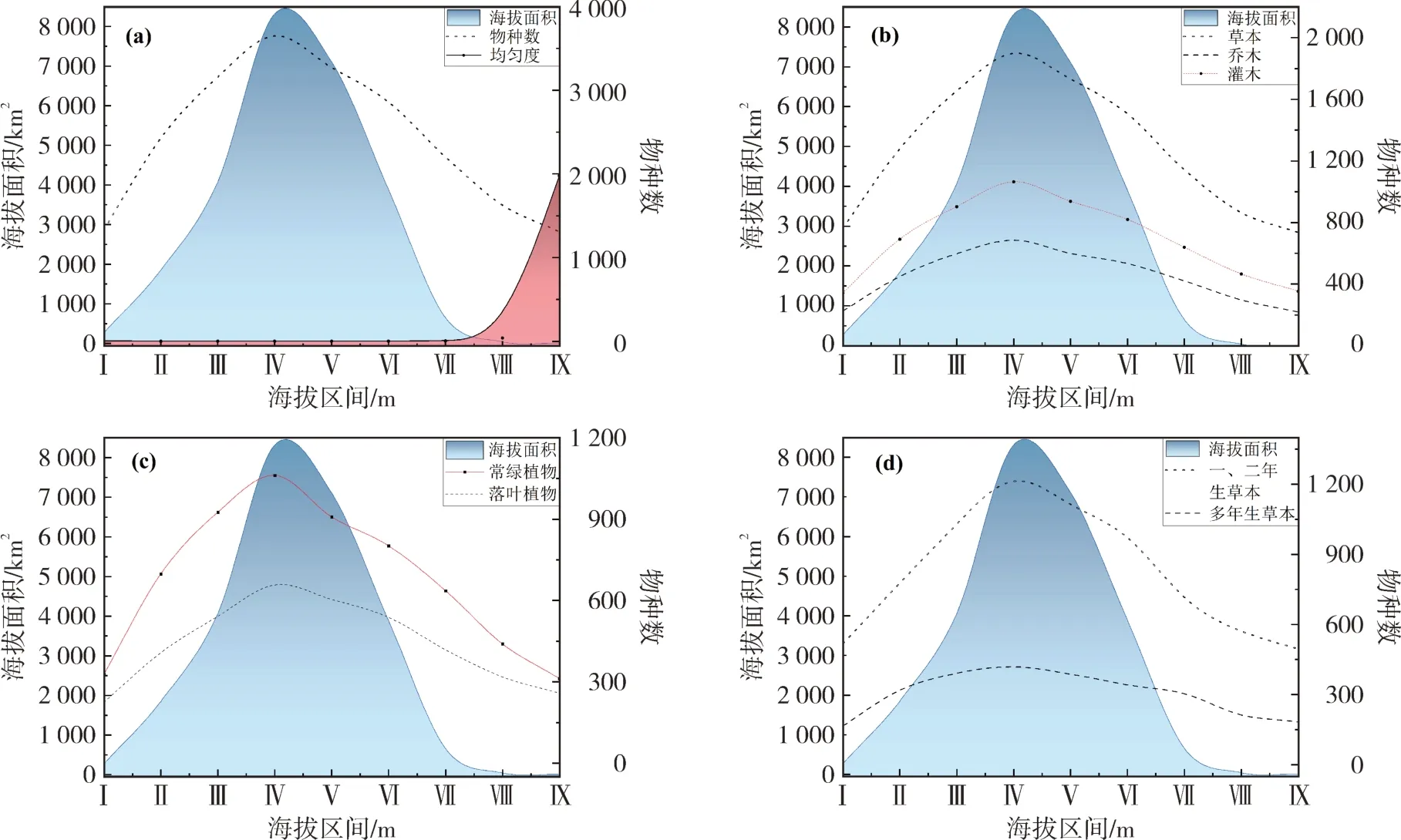
图4 黔南州高等植物垂直分布特征Fig.4 Vertical distribution characteristics of higher plants in Qiannan Prefecture
2.5 物种与环境因子相关关系
由RDA 排序结果可知(图5),前两个坐标轴的排序(RDA1 和RDA2)解释了总方差的75.3%,物种丰富度、分化度及密度信息能够为以下7 个环境变量所解释。其中,海拔(DEM)的解释率最高,解释了总方差的50.7%。物种丰富度(SR)、分化度(SD)及密度(SDD)与年降水量(BIO12)、最湿季降水量(BIO17)、植被净初级生产力(NPP)及县域面积(CA)呈正相关。年降雨量越多、植物固定和转化光能化合物的效率越高,物种越丰富、分化,且密度越高。同时,物种分化度与年降雨量相关性高于最干季降雨量、净初级生产力,物种丰富度、密度与植被净初级生产力相关性较高。丰富度、分化度、密度3 者均与人口密度(PD)、经度、海拔呈负相关,与海拔的相关性达到显著水平(P< 0.05)。反映出往西部地区人口越密集的高海拔地区,物种丰富、分化、密度均呈下降趋势,主要受海拔因素影响较大。
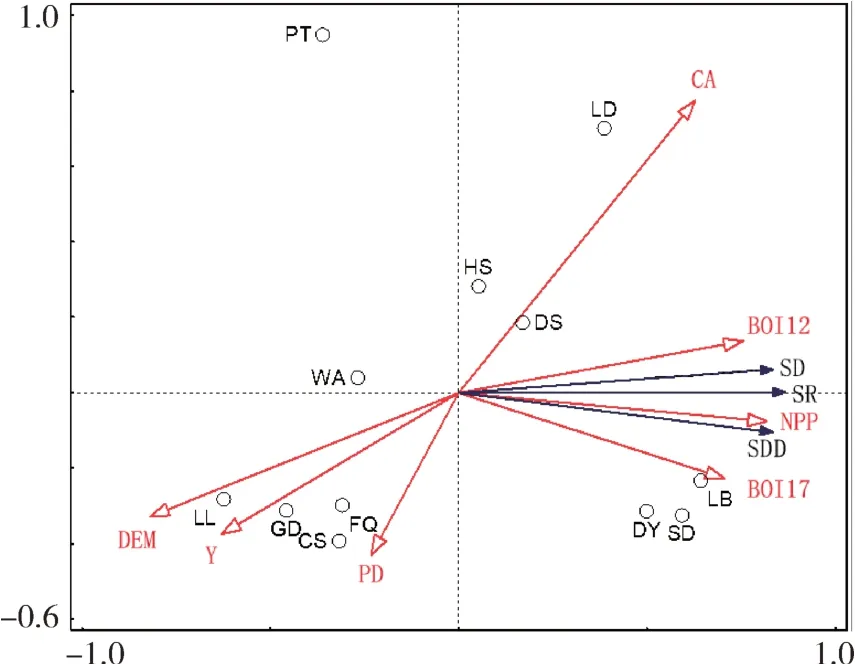
图5 物种信息与环境因子的冗余分析Fig.5 Redundancy analysis of species information and environmental factors
3 讨论与结论
黔南州高等植物物种资源丰富,组成复杂多样,种子植物占优势地位,相较于其他类别物种更具有适应复杂多样生境条件的生存策略[25]。不同研究区物种相似性与相邻或相近区域物种规律性聚为一类的现象,说明生存物种的趋同或趋异现象与地理位置和环境条件之间存在密切联系,地理环境的相似性是驱动物种多样性产生差异的重要因素[26]。空间分布特征从水平分布角度上看,黔南州东南部地区地理位置的生存环境、水热条件等环境因素对物种丰富度高值聚类影响较大,相邻区域物种间的联系更为密切。东南地区向西北地区过渡,物种丰富度逐渐降低的分布特点,与分化度的分布格局相似。物种丰富度高值区与分化度高值地区相重合,两者之间存在相对应的关系,与刘经伦等[27]研究结果相似,造成此现象的主要原因可能是越分化的植物类群表现出越强的环境适应能力,更容易获得形成新种的环境机遇,物种多样性越高。同时也可能同域同源类群分化受到抑制导致丰富度降低的影响,丰富度与分化度之间存在密切联系但不是唯一相对应的关系[28]。具有热带性质起源背景物种在重新面对寒化生存环境或其他新环境时,更具有产生新物种的环境机遇,则物种分化度越高越丰富,若物种处于分化初期,可能会出现物种丰富度低值区与分化度高值区相一致的现象[29-30]。黔南州位于云贵高原东南部边缘斜坡地带,山脉水系对环境土壤、养分等有切割作用,生境异质性较高,是影响物种丰富度空间分布格局产生差异的主要原因[31]。
从垂直分布角度分析,“单峰型”的海拔丰富度格局与众多学者的研究结果相一致[32],但多项研究表明,物种沿海拔梯度的分布格局具有多样性[33-34],不遵循固定的分布模式,其中“单峰型”分布模式最为普遍,与解释物种丰富度分布格局的五大模型之一的“中域效应”假说理论相符。海拔面积与种丰富度之间的线性关系也是最主要的影响因素之一,曾被认为是生态学中几个真正的定理之一[35],黔南州海拔面积很可能是影响物种海拔分布格局的主要因素。消除面积影响物种多样性在高海拔地区出现聚集分布特征,可以用“负密度制约假说”中物种受资源竞争、有害生物侵害等导致群落降低同种个体生存,为不同物种提高生存空间,促进不同物种共存来解释[36]。划分不同层次物种海拔丰富度格局存在小差异大共性(单峰型分布格局),有研究发现,不同的类群、进化史、功能型的物种丰富度格局不同,能反映物种多样性格局形成机制上的重要差异[37],该分布特征与Werff 和Consiglio[38]的研究结果相一致,与Gebrehiwot 等[39]研究结果不同。有些学者认为,“单峰型”物种丰富度分布模式是环境梯度造成的,相较于“中域效应”的结果,“水分梯度”效应对物种丰富度分布格局产生影响更大,干旱地区“单峰型”分布模式完全可以用“水分-能量平衡”假说解释[40]。综合冗余分析结果讨论,黔南州物种多样性与海拔、年降雨量、最干季降雨量、植被净初级生产力和区域面积等生境因素间有着密不可分的关系,海拔因素对物种多样性解释率高达50.6%,生境脆弱且缺水的浅土层成为物种生存和繁殖的关键限制因素[41]。与Albuquerque 等[42]对英国木本及草本植物多样性与环境因子决策研究结果相一致,可能由于海拔梯度下潜在的水分、土壤、养分、温度等环境梯度变化对物种的过滤筛选作用有关[43]。深入研究分析物种多样性与空间分布特征,利于揭示该地区植物物种自身特性、生境异质性、物种与生存环境相适应的生态过程,助于监测和预测当地区自然生态系统发生发展和变化趋势,为该区域下一步物种多样性保护与管理提供理论支撑。
猜你喜欢
清华金融评论(2022年4期)2022-04-13 21:33:11
散文诗(2021年22期)2022-01-12 06:14:04
国际放射医学核医学杂志(2021年10期)2021-02-28 08:43:46
环球时报(2019-05-23)2019-05-23 06:17:36
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
乡村地理(2018年3期)2018-11-06 06:51:10
贵茶(2018年4期)2018-09-28 09:04:32
贵茶(2018年6期)2018-05-30 09:53:44
——黔南州老年大学校歌
老年教育(老年大学)(2017年3期)2017-03-23 08:15:35
河北遥感(2014年4期)2014-07-10 13:54:58
