
空心莲子草ApCAT 基因家族鉴定和表达模式
2023-12-11 10:09:26韩晓文胡义锋王梦如陈中义朱永兴尹军良
草业科学 2023年11期
韩晓文,韩 硕,胡义锋,王梦如,陈中义,朱永兴,尹军良
(1.长江大学湿地生态与农业利用教育部工程研究中心 / 长江大学农学院 / 长江大学园艺园林学院, 湖北 荆州 434025;2.中南民族大学生命科学学院, 湖北 武汉 430074)
空心莲子草(Alternanthera philoxeroides),也称水花生、空心苋,属多年生宿根草本植物,原产于南美洲,是一种“水陆两侵”的外来入侵物种[1]。目前,空心莲子草在20 余个省(市)均有分布,黄河以南的区域为害尤其严重,据2016 年统计,全国中度以上(盖度大于30%)危害面积超过500 万hm2,直接经济损失每年16 亿~18 亿元[2],如何高效防治空心莲子草已成为国内外研究的热点之一。
植物受到生物/非生物胁迫时,其细胞内的氧化还原动态平衡被打破,导致过氧化氢(H2O2)、羟自由基(OH-)、超氧阴离子(O2-)等氧自由基(ROS)的过量产生[3-4]。适量的ROS 在生物体内可作为第二信使参与细胞信号转导,激活免疫细胞、引起细胞增殖、衰老和凋亡[5],但ROS 的过度积累会导致植物细胞的氧化应激损伤[3]。其中过氧化氢酶(catalase, CAT)能够通过催化电子转移,将生物体内多余H2O2还原成H2O 和O2,有效清除植物光呼吸和线粒体电子传递过程中产生的过氧化氢[6]。CAT 的相对分子质量在200~400 kDa,通常由4 个亚基组成[7]。每个亚基在羧基端含有1 个血红素分子和1 个CAT 体靶向信号肽(PTS1)序列[8]。根据CAT催化中心结构的不同,CAT 可分为铁卟啉结构CAT(FeCAT)和锰卟啉结构CAT (MnCAT) 。
大量研究表明,CATs基因具有时空表达特异性,例如玉米(Zeamays)ZmCAT1和ZmCAT3在籽粒发育整个过程均有表达,ZmCAT2仅在籽粒发育后期表达[9-10]。水稻(Oryzasativa)OsCATA和OsCATC的表达受昼夜节律调控,OsCATA在晚上特异性表达,OsCATC在清晨特异性表达[11]。同时CATs基因在胁迫应答响应过程中发挥重要作用[12]。拟南芥(Arabidopsisthaliana) AtCAT1 通过降解H2O2来响应非生物胁迫,此外,AtCAT2 和AtCAT3 可以降解H2O2来维持细胞内ROS 的动态平衡[13];过表达OsCATA和OsCATC提高了水稻的抗旱性,同时OsCATC 也可以被STRK1 磷酸化激活,从而提高水稻的耐盐性和抗氧化性[11];ZmCAT2转基因烟草(Nocetoana tabacum)与野生型相比具有更强抗病性和活性氧胁迫的耐受性[14]。此外,CAT 在植物响应除草剂胁迫中发挥重要作用,例如不同浓度的乙草胺和阿特拉津均能使肉芝软珊瑚和短指软珊瑚CAT 酶活性快速上升,且在一定范围内,CAT 酶活性随着除草剂浓度升高和胁迫时间延长而持续上升[15]。不同浓度的噻隆·异噁酮施药7、14 d 时玉米CAT 酶活性显著提高,且活性随着施药时间的延长和药剂浓度的降低而逐渐降低[16]。
到目前为止,CAT基因家族已在小麦(Triticum aestivum)[5]、水稻[11]、油菜(Brassicanapus)[17]等多种植物中进行了鉴定分析,并对其胁迫响应模式进行了研究,但尚缺乏对空心莲子草CAT 家族成员的系统认识。因此,本研究对空心莲子草的CAT基因进行系统鉴定,并分析了该基因家族成员在除草剂胁迫下的表达模式,旨在了解CAT 在空心莲子草胁迫应答响应过程中的潜在作用,为空心莲子草的防治提供新思路。
1 材料与方法
1.1 空心莲子草ApCAT 的鉴定
获取空心莲子草基因组数据库[18]。从Pfam(http://pfam.xfam.org/)下载CAT (PF00199)的隐马尔可夫模型序列,BLASTp 搜索空心莲子草氨基酸序列数据库,鉴定候选ApCAT (e-value < 1e-10)[19]。获得的候选序列通过Pfam (v35.0,http://Pfam.xfam.org)和 InterProScan (v93.0, http://www.ebi.ac.uk/InterPro Scan)进行确认,排除不包含CAT 结构域的序列。
1.2 ApCAT 系统发育分析
使用ClustalW2 软件将ApCAT 与已知物种(水稻、拟南芥) CAT 氨基酸序列进行多序列比对。使用ITOL 工 具(Interactive tree of Life, http://ITOL.embl.de)绘制包含ApCATs、AtCATs 和OsCATs 的系统发育树,说明ApCATs 的系统发育关系,并辅助它们的命名[20]。
1.3 ApCAT 保守结构域和基序分析
根据GFF3 基因结构注释信息,利用TBtools 绘制ApCATs 的外显子/内含子结构图。使用MEME(版 本 5.5.1, http://meme-suite.org/tools/meme)识 别ApCATs 的保守基序[21]。参数设置如下:每个序列可以包含任意个的非重叠基序,不同基序的数量为10,基序宽度范围为6 到50 个氨基酸。利用TBtools绘制蛋白基序结构图。
1.4 ApCAT 蛋白质特征分析
使用ExPASyServer10 (https://prosite.expasy.org/PS 50011)分析ApCATs 的蛋白质基本特征,包括蛋白序列长度(Len)、分子量(MW)、等电点(pI)、总亲水性(GRAVY)[22]。使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/Plant multi)预测ApCATs 的亚细胞定位[23]。使用SWISS-MODEL (https://www.swissmodel.expasy.org/)对ApCATs 的三维结构进行同源建模。
1.5 ApCATs 表达模式分析
从NCBI 的SRA 数据库收集4 组空心莲子草的转录组测序数据[24]。详情信息:PRJNA256235,陆地和池塘中采集的混合组织样本;PRJNA256237,低钾胁迫下根部组织样本;PRJNA263573,水中和盆栽种植的茎节组织样本;PRJNA268359,山东济南收集的“北方”种群和上海收集的“中央”种群上部第1~3 对叶片组织样本。通过Cufflinks 计算ApCATs基因的表达水平(用标准化的FPKM 值表示)。使用log2(FPKM + 1)值,通过R 软件包pheatmap 绘制热图,展示不同条件下ApCATs基因的表达模式[25]。
1.6 除草剂防效测试
试验于10 月中旬在长江大学农学院实验田中进行,选取平均高度为10~15 cm、长势一致的空心莲子草植株,每次处理重复3 次,共18 个小区,小区面积1 m2,每个处理喷施药液量为250 mL,重复3 次,空白对照为清水。分别于0.5、1、3、5、7 d 观察防治效果并取样。供试药剂及浓度如表1 所列。
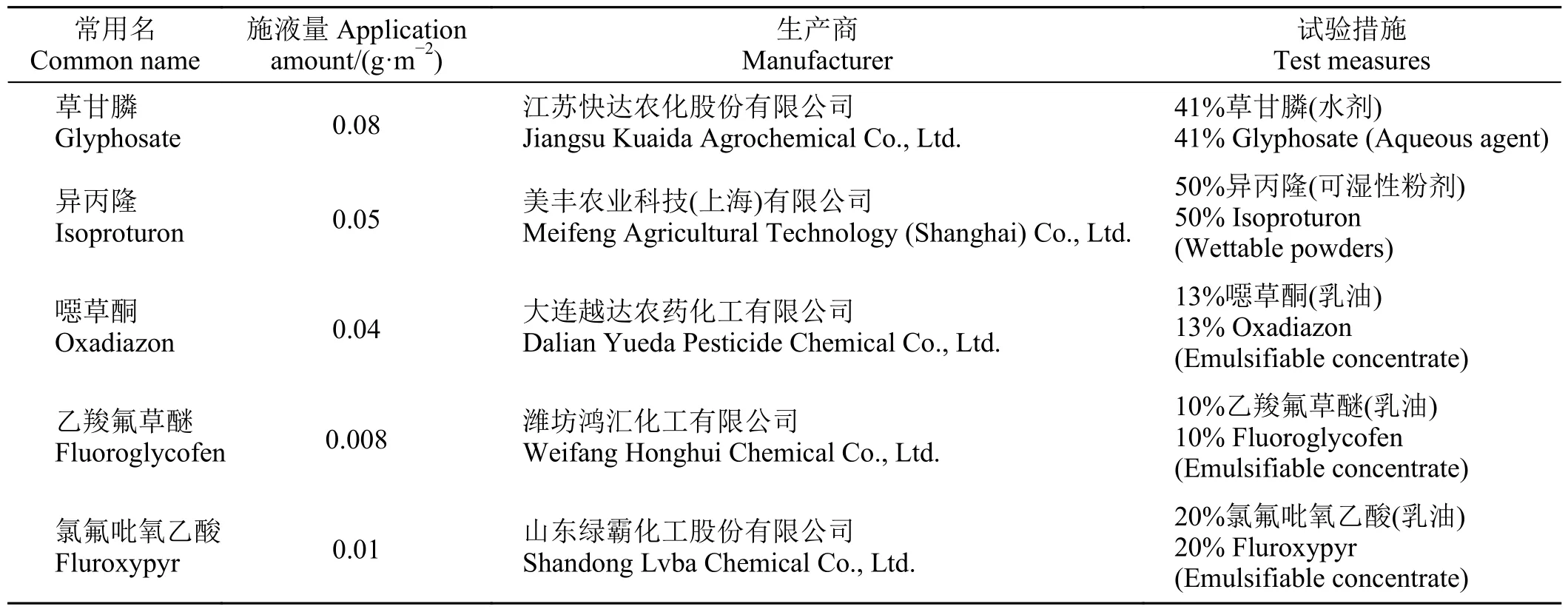
表1 试验药剂和浓度Table 1 Data for the agents and their concentrations
1.7 RT-qPCR 分析
除草剂处理0.5、1、3、5、7 d 时分别收集顶端嫩叶,-80 ℃保存。使用TRizol 试剂(GenStar,中国北京)提取总RNA,并用DNaseI (TaKaRa,中国大连)去除DNA。使用RevertAid 逆转录酶(Vazyme,中国南京)将RNA 逆转录为cDNA。然后将cDNA稀释至100 ng·μL-1。按照制造商的说明书,使用SYBR GreenMaster Mix (Vazyme,南京,中国)在CFX96 实时PCR 系统(BioRad,HercuLes,CA,USA)上进行RT-qPCR。使用PrimerPremier5 软件设计基因特异性引物。20 μL RT-qPCR 反应体系由10 μL 2 × SYBR Premix Extaq、正向和反向引物各0.4 μL、cDNA 2 μL 和7.2 μL ddH2O 组成。试验流程如下:95℃预变性30 s (步骤1);95 ℃变性5 s (步骤2),58℃退火15s,72 ℃延伸30 s,60 ℃荧光信号采集30 s(步骤3);步骤2 至步骤3 循环40 次。
1.8 统计分析
使用2-ΔΔCt方法计算ApCAT基因的相对表达水平[5],运用Excel 2016 对数据进行整理分析,使用单因素方差分析来计算平均值之间的显著差异,本研究差异显著水平均为P< 0.05。
2 结果与分析
2.1 ApCAT 的鉴定及系统发育分析
为鉴定空心莲子草基因组中的ApCAT基因,以CAT (PF00199)种子序列作为参考序列进行本地BLASTp,并利用Pfam 验证候选序列中是否含有CAT 结构域。最终从空心莲子草基因组中鉴定到20 个ApCAT基因(图1)。根据系统发育树所示的进化关系,将它们分别命名为ApCAT1到ApCAT20。根据系统发育树的聚类情况,将ApCAT基因家族分为3 个亚群(Group a、b、c)。
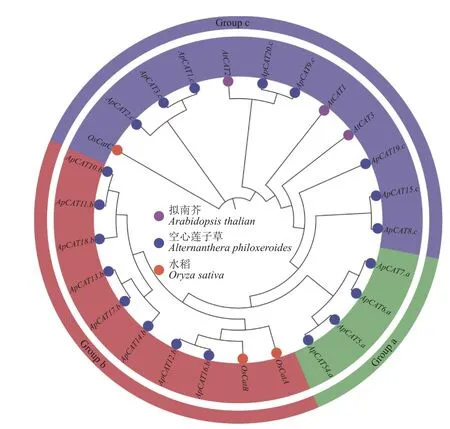
图1 空心莲子草、拟南芥和水稻CAT 成员的系统发育分析Figure 1 Phylogenetic analyses of catalase (CAT) from Alternanthera philoxeroides, Arabidopsis thaliana,and Oryza sativa
2.2 ApCATs 蛋白基序和基因结构
根据GFF3 基因结构注释信息,使用TBtools 绘制ApCATs的外显子/内含子结构图,并根据系统发育树的顺序排列(图2A)。所有ApCATs均不含内含子,其中ApCAT7.a、ApCAT17.b和ApCAT16.b仅在5’端包含UTR 区域;ApCAT2.c、ApCAT8.c、ApCAT15.c、ApCAT9.c和ApCAT1.c仅在3’端包含UTR 区域;而ApCAT13.b和ApCAT19.c在两端均含有UTR 区域(图2B)。保守基序分析发现,从ApCATs 中鉴定到10 个motif,其中ApCATs3.c 含有的motif 最少,为2 个;ApCATs13.b 含有的motif 最多,为11 个;结合系统发育分析发现,亲缘关系越近的ApCATs 含有越相似的motif (图2A、B)。结构域分析发现,motif 1 到motif 4 和motif 7到motif 9 均为氧化氢酶核心结构域(catalase domain),motif 5 为过氧化氢酶相关的免疫应答结构域(catalase-related immune-responsive domain)。其他motif 未匹配到已知的关键功能结构域(图3)。

图2 ApCATs 系统发育树(A)、ApCATs 基因结构(B)、ApCATs 蛋白motif 分布(C)Figure 2 Phylogenetic tree of ApCATs (A), gene structure of ApCATs (B) , and motif distribution of ApCATs (C)
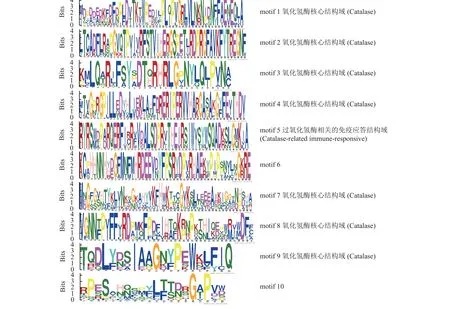
图3 ApCATs 保守基序Figure 3 Conservative motif in ApCATs
2.3 ApCATs 蛋白特征和三维结构
蛋白质特性分析发现,ApCATs 氨基酸数量平均值为194 aa,分布范围为74~492 aa,分子量平均值为22.32 kDa,分布范围为8.51~57.05 kDa。等电点平均值为7.39,分布范围为4.26~9.69,平均值小于8,故大部分ApCATs 为酸性蛋白质。不稳定指数平均值为32.52,分布范围为13.72~49.10,除ApCAT1.c、 ApCAT3.c、 ApCAT9.c、 ApCAT13.b、ApCAT15.c、ApCAT19.c、ApCAT20.c 外,其他ApCATs属于稳定蛋白质。亲水性平均值-0.55,分布范围为-1.17~-0.05,所有亲水性的数值小于0,说明ApCATs 都是亲水性蛋白质。亚细胞定位预测分析发现,所有ApCATs 均分布在过氧化物酶体中,同时部分ApCATs 也预测可能分布在线粒体、叶绿体和细胞核内(表2)。

表2 ApCATs 蛋白质特征分析Table 2 Characterization of Alternanthera philoxeroides ApCAT proteins
通过SWISS-MODEL 完成了ApCATs 的三维同源性建模(图4),所有的ApCATs 主要由α-螺旋、β-转角、延伸链和随机卷曲构成,而这4 种结构在ApCATs 蛋白中的比例不同,随机卷曲所占比例最大(23.71~51.14%),其次是α-螺旋(14.58%~37.11%)和延伸链(9.60%~33.98%),β-转角所占比例最小(5.75%~19.42%)。

图4 使用SWISS-MODEL 生成的ApCATs 的3D 模型预测Figure 4 Predicted three-dimensional models of ApCATs generated by using SWISS-MODEL
2.4 ApCATs 的表达模式分析
ApCAT1.c、ApCAT2.c、ApCAT3.c、ApCAT12.b、ApCAT14.b、ApCAT16.b、ApCAT17.b、ApCAT18.b和ApCAT19.c在不同处理下的表达水平普遍较高,而其他ApCATs表达水平偏低或不表达。在水中和盆栽种植的茎节组织样本中,ApCAT14.b、ApCAT18.b、ApCAT17.b、ApCAT3.c和ApCAT19.c的 表 达 量 先 上升,在12 h 时达到峰值后下降;而ApCAT16.b的表达量先下降,在12 h 降到最低后回升。此外,根系组织在低钾胁迫下,ApCAT6.a、ApCAT14.b、ApCAT17.b、ApCAT18.b和ApCAT19.c的 表 达 量 下 降,ApCAT4.a和ApCAT12.b的表达量上升。对于空心莲子草的不同种群,除了ApCAT14.b在山东济南收集的“北方”种群中的表达量高于上海收集的“中央”种群外,其余ApCATs在两个种群中的表达水平相差不大(图5)。

图5 不同组织和处理的ApCATs 表达热图Figure 5 Heat map of ApCATs in different tissues and under different treatments
2.5 除草剂对空心莲子草的防治效果
5 种除草剂对空心莲子草的防治效果不同。20%氯氟吡氧乙酸和13%噁草酮防治效果最好,20%氯氟吡氧乙酸在药后5 d 时引起植株退绿、萎蔫,停止生长,而13%噁草酮在药后3 d 时引起顶端幼嫩组织坏死;其次是10%乙羧氟草醚和41%草甘膦,在药后5~7 d 时引起顶端幼嫩组织失绿枯黄,并且41%草甘膦在药后7 d 时开始出现褐色枯斑;50%异丙隆防治效果不明显,药后7 d 后均无明显变化(图6)。

图6 5 种除草剂药对空心莲子草的生长的影响Figure 6 Influence of five herbicides on Alternanthera philoxeroides growth
2.6 ApCATs 的除草剂诱导表达模式分析
挑选6 个高水平表达的ApCATs 基因,利用RTqPCR 检测其在5 种农药处理下的相对表达量(图7)。ApCATs 对5 种农药处理均有响应。
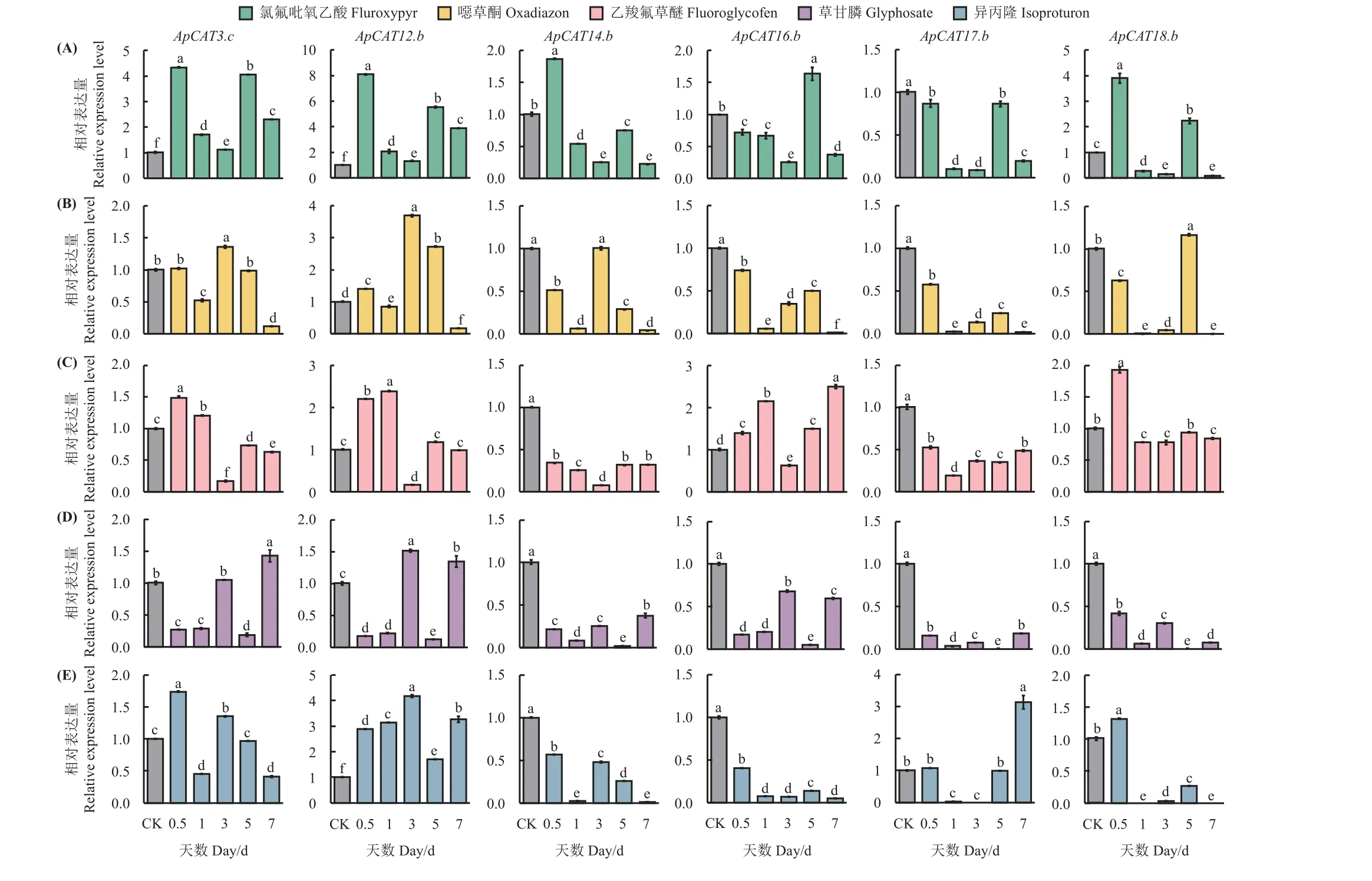
图7 5 种除草剂不同处理时间下ApCATs 的相对表达量Figure 7 Expression patterns of ApCATs in the presence of the five herbicides
在氯氟吡氧乙酸处理下,2 个ApCATs的表达量呈下降趋势,4 个ApCATs的表达量呈先升高后降低的趋势,但5 d 时6 个ApCATs表达量显著升高,其中ApCAT12.b的表达量升高最显著,是CK 的5.5倍。此外,6 个ApCATs在3 d 时表达量均降低,其中ApCAT17.b最显著,仅为CK 的9% (图7A)。
在 噁 草 酮 处 理 下,ApCAT3.c、ApCAT12.b和ApCAT14.b在3~7 d 时有明显的下降趋势,ApCAT16.b、ApCAT17.b和ApCAT18.b在1~5 d 时有明显的上升趋势。此外,在1 和7 d 时6 个ApCATs的表达量均下降,其中ApCAT18的表达量最低,分别只有CK 的0.3%和0.2% (图7B)。
在乙羧氟草醚处理下,5 个ApCATs 呈先升高后下降再升高的趋势,4 个ApCATs 在3 d 时表达量显著降低,其中ApCAT14.b的表达量最低,仅为CK 的0.8%,而ApCAT17.b在1 d 时表达量最低,为CK 的19%。此外ApCAT18.b在0.5 d 时表达量升高最显著,是CK 的1.9 倍,随后在1 d 时下降并稳定在CK 的78%~94%(图7C)。
草甘膦处理下,6 个ApCATs基因的相对表达量呈下降趋势(图7A),其中ApCAT14.b、ApCAT17.b和ApCAT18.b表达量降低最为明显,分别下降至CK 的19%、9%和17%。ApCAT3.c和ApCAT12.b,在3 和7 d 时表达量高于CK,最高达到1.5 倍。(图7D)。
异丙隆处理下,处理1 d 时,ApCAT12.b表达量与CK 相比上升3.1 倍,其余5 个ApCATs均低于CK,其中ApCAT18最低,仅为CK 的0.4%。此外ApCAT12在0~7 d 时表达量均较高,是CK 的1.7~4.2 倍,而ApCAT14.b、ApCAT16.b和ApCAT18.b表达量持续下降,尤其是ApCAT16.b在处理1 d 后均不足CK 的15% (图7E)。
3 讨论
空心莲子草在2003 年被列入“中国第一批外来入侵物种名单”,具有极强表型可塑性和入侵性,对我国生态系统和社会经济造成恶劣的影响。已有研究表明CAT 在空心莲子草响应锌、铜等重金属胁迫[26-28]和涝渍胁迫[29]中发挥调控作用。同时,CAT在植物响应除草剂中发挥重要作用[30-33],而CAT 在空心莲子草响应除草剂中的作用尚未研究。因此,开展基因组水平空心莲子草ApCATs基因的系统鉴定和分析,了解其表达模式,尤其揭示其响应除草剂的诱导表达特征,可为开展ApCATs逆境胁迫响应功能研究奠定基础,也为空心莲子草的有效防治提供指导。
本研究从空心莲子草全基因组中鉴定了20 个ApCAT基因家族成员(ApCAT1~ApCAT20),其成员数量高于拟南芥(3 个)[13]、水稻(3 个)[11]、小麦(10 个)[5]和甘蓝型油菜(14 个)[17]。其中ApCAT编码的蛋白质长度为74~492 aa (表2),其蛋白长度差异较大且相对于其他物种来说长度较短,例如在油菜 中[17],BnCATs 长 度 为477 (BnCAT13)~1 040 aa(BnCAT10); 在 小 麦 中[5], TaCATs 长 度 为 290(TaCAT3-A2)~494 aa (TaCAT2-A);而水稻[11]和大豆[34]中,所有的CAT 长度均为492 aa。通过保守基序分析发现,ApCAT基因含有10 个motif,其中motif1 到motif5、motif7 到motif9 均注释为过氧化氢酶功能结构域。此外结合系统发育分析发现,处于同一亚组的ApCAT 含有越相似的基因结构和motif,但个别家族成员的motif 数量存在一定的差别,推测该现象是因为ApCAT 在进化过程中丢失或新增Motif 所致[35]。
为了解ApCATs基因的表达水平,借助已经发表的RNA-seq 数据,对20 个空心莲子草CAT基因表达水平和表达模式进行了初步分析,并揭示了在不同环境条件下的ApCATs 表达水平(图6)。本研究表明,ApCATs的表达具有时空特异性。ApCAT14.b、ApCAT18.b、ApCAT17.b、ApCAT16.b、ApCAT12.b、ApCAT3.c、ApCAT2.c、ApCAT19.c和ApCAT1.c在 不同处理下(低钾胁迫、水分变化)的表达水平普遍较高,并且在不同的水分环境中总体表达量变化趋势相似。因此推测不同的水分环境的变化对ApCAT基因的表达影响较小。此外,在山东济南收集的“北方”种群和上海收集的“中央”种群中除ApCAT14.b的表达量在叶片差异较大外,其余ApCATs的表达水平相差不大。这表明,在正常生长的情况下,空心莲子草ApCAT的表达量受纬度、海拔、温度、光照等条件的影响较小,推测ApCATs的表达可能受到特定环境条件的诱导,例如干旱[36]、盐[37]、低温[38]等胁迫。
为了研究除草剂诱导ApCAT的表达特征,本研究对6 个高水平表达的ApCATs在5 种除草剂胁迫下的表达模式进行了系统分析。结果发现,20%氯氟吡氧乙酸药后5 d 时植株开始出现失绿、黄花的症状;药后7 d 时植株已经完全枯黄萎蔫,停止生长。与本研究结果类似,高宗军等[39]的研究表明,20%氯氟吡氧乙酸(100 g·hm-2, 有效成份)会使空心莲子草叶片紫化、萎蔫,并在10 d 时引起植株死亡。同时,表达模式分析发现ApCAT3.c、ApCAT12.b和ApCAT16.b在5~7 d 时的表达量均显著高于CK。而在水稻中,各浓度下氯氟吡氧乙酸在6 d 时能够引起水稻根系和叶片组织中活性氧的积累,同时积累程度随着浓度的增加而升高;且低浓度的氯氟吡氧乙酸(0.1 mg·L-1)显著增加了CAT 酶活性和CAT同工酶的表达量[32]。因此推测在空心莲子草中,氯氟吡氧乙酸可能同样导致植物体内活性氧的积累,从而诱导CAT基因的表达。
在噁草酮处理中,3 d 时出现了顶叶白化的现象。类似的,在高宗军等[39]的研究中,13%噁草酮(400 g·hm-2)药后空心莲子草叶片白化、干卷,并在3 d 时死亡。同时,水稻植株噁草酮(1 270 g·hm-2)药后6 d,CAT 酶活性增加了22%,CAT 同工酶的表达量也显著增强[31]。本研究中,ApCAT12.b和ApCAT18.b的表达量在5 d 时显著高于CK,而其余4 个ApCATs在6 d 左右时的表达量并未高于CK,并且所有ApCATs的表达量在7 d 时均显著降低,其原因未知,需要进一步的研究。
在乙羧氟草醚处理中,10%乙羧氟草醚药后7 d 时也引起心叶枯黄,而相关研究表明,20%乙羧氟草醚(80 g·hm-2)在药后5 d 时会引起空心莲子草心叶卷曲枯黄[39],这一差异可能是由于药剂浓度不同而导致的。表达模式分析结果表明,ApCAT3.c、ApCAT12.b、ApCAT16.b和ApCAT18.b在 施 药1 d 内表达水平显著升高,推测这4 个ApCATs在药后短时间内受到乙羧氟草醚的诱导。
在草甘膦的处理中,空心莲子草在41%草甘膦药后7 d 时出现轻微枯黄并开始出现枯斑。而高宗军等[39]研究表明,41%草甘膦(800 g·hm-2)在药后4 d 时引起空心莲子草叶片枯黄并有枯斑,与本研究结果存在一定的差异,推测原因可能是喷施方式和施药剂量不同。在玉米[40]、莎草(Cyperus rotundus)[41]和大豆(Glycine max)[42]等植物中,草甘膦药后3 d会引起植株体内CAT 酶活降低。而类似的,在本研究中ApCAT14.b、ApCAT16.b、ApCAT17.b和ApCAT18.b的表达量在药后7 d 内均较低,推测草甘膦可以通过抑制CAT基因的表达,从而降低CAT 酶活性。
在异丙隆处理中,空心莲子草在50%异丙隆处理7 d 内均无任何变化,而相关研究表明,异丙隆一种持效型除草剂,在药后10 d 后才会引起空心莲子草植株死亡[39]。结合表达分析发现,ApCAT3.c、ApCAT12.b和ApCAT18.b的表达量在0.5 d 时显著升高,并且ApCAT12.b在0.5 d 后继续升高,在3 d 时达到峰值,而ApCAT17.b在处理7 d 时显著升高并达到峰值。因此推测ApCAT12.b在处理前期受异丙隆的诱导,ApCAT17.b在处理后期受到诱导。
4 结论
本研究共鉴定出20 个ApCATs基因。所有的ApCATs 均只含有一个CAT 结构域,且均为亲水性蛋白;所有ApCATs 均预测定位于过氧化物酶体,个别ApCATs 也有定位线粒体、叶绿体和细胞核的可能。ApCAT成员受纬度、海拔、温度、光照、水分条件等影响较小,表达水平相对稳定,但响应除草剂胁迫而诱导差异表达。20%氯氟吡氧乙酸和13%噁草酮防治效果好,结合RT-qPCR 分析结果表明,ApCAT 家族成员在空心莲子草响应除草剂处理过程中显著差异表达,参与对除草剂胁迫的应答过程。
猜你喜欢
新医学(2023年10期)2023-12-09 15:04:51
中老年保健(2022年7期)2022-09-20 01:16:16
南方医科大学学报(2022年3期)2022-04-13 01:51:26
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10 07:07:36
今日农业(2019年15期)2019-01-03 12:11:33
红领巾·探索(2018年12期)2018-01-26 12:34:14
现代园艺(2017年19期)2018-01-19 02:50:21
小学生作文选刊(2017年6期)2017-07-03 12:16:36
丝路艺术(2017年5期)2017-04-17 03:11:51
营销界(2015年23期)2015-02-28 22:06:18
