
金属抗菌肽SIF4基于胞内生物大分子靶点的大肠杆菌非膜损伤抑菌机理
2023-12-11 06:57:56李玉珍肖怀秋曾梦琪赵谋明
食品科学 2023年21期
李玉珍,肖怀秋,*,刘 淼,王 琳,曾梦琪,赵谋明
(1.湖南化工职业技术学院制药与生物工程学院,湖南 株洲 412000;2.华南理工大学食品科学与工程学院,广东 广州 510000)
大肠杆菌(Escherichia coli)是引起食品公共安全事故与食物中毒的主要病原菌,由被污染食品或人际传播,通过产生耐药酶或改变药物靶点、改变膜通透性、改变外膜孔蛋白结构、构建主动外排机制或形成生物膜获得耐药性,多重交叉耐药菌株的出现使食源性致病菌防控变得复杂[1]。吴萱等[2]从北京市6 个辖区大型超市和农贸市场随机采集生鲜肉样本259 份,大肠杆菌检出率65.25%,分离菌株对甲氧苄氨嘧啶-磺胺甲恶唑高度耐药;李明昊等[3]从山东不同地市41 个连锁超市采集猪肉和鸡肉样品201 份,大肠杆菌检出率100%,鸡肉源菌株对头孢菌素类耐药率极显著高于猪肉源菌株(P<0.01),有48 株菌携带blaCTX-M耐药基因;唐雪林等[4]从乌鲁木齐市7 区19 家超市采集畜禽肉样品353 份,大肠杆菌检出率71.39%,其对四环素和氨苄西林耐药性最高,耐药基因A检出率高达90.4%;马振报等[5]从广州市不同区域零售市场和超市采集畜禽肉样本323 份,大肠杆菌检出率74.61%,对氨苄西林(63.07%)、多西环素(47.72%)和复方新诺明(43.15%)耐药率位列前三。
抗菌肽具有抗菌谱广、稳定性好、特异性强、对哺乳动物细胞毒副作用少且不易产生耐药性等优势,可多靶点作用于食源性致病菌,是抗生素最具潜力的替代品,特别适用多重耐药性食源性致病菌生物防控[6]。根据抗菌肽作用靶点不同,可分为靶向作用于细胞质膜的抗菌肽和非细胞质膜抗菌肽两类[6]。天然抗菌肽(如G1OLO-L2OL2[7]、Arg-Ser-Ser[8]和zp37[9]等)可通过破坏细胞质膜结构和扰乱其生物功能造成胞内容物外泄和菌体凋亡,细胞质膜是主要作用靶点。研究表明,部分抗菌肽并不破坏细胞质膜结构和(或)扰乱细胞质膜功能,而是透过细胞质膜并靶向作用于胞内细胞器或生物大分子,如胞内核酸、拓扑异构酶、电子传递酶、热休克蛋白、微管蛋白或G蛋白、信号通路受体蛋白等,或影响基因复制、转录与表达调控以及摧毁细胞骨架等[10],如Buforin II可在不破坏细胞质膜前提下特征性与胞内DNA结合[11];Indolicidin可抑制DNA合成并影响RNA和蛋白质合成,还能引发SOS修复途径,引起DNA降解[12];Linezolid可特征性结合核糖体50S亚基以抑制蛋白质起始复合物形成[13];Histatin可与细胞质膜特定受体结合并诱导胞内ATP损失,破坏细胞周期并导致活性氧簇产生[14]。部分抗菌肽具有细胞质膜/非细胞质膜双重效应靶点,如Indolicidin[15]和β-defensin[16]等。前期研究发现,金属抗菌肽(以下简称金抗肽)SIF4对大肠杆菌有较好的抑制活性,并可破坏细胞质膜结构、增强膜通透性、诱导膜去极化和膜氧化损伤,导致胞内物质泄漏[17],或抑制糖酵解途径已糖激酶活性从而调控糖氧化效率[18],或影响呼吸代谢、削弱细胞质膜离子通道ATP酶活性和抑制胞内ATP合成从而抑制菌体呼吸与能量代谢[19],或破坏菌体细胞壁结构和改变菌体形貌[20];综上,金抗肽SIF4可基于细胞质膜/非细胞质膜对大肠杆菌实施抑菌,但基于大肠杆菌胞内生物大分子靶点的非细胞质膜抑菌机理尚不明晰,为揭示SIF4对大肠杆菌基于胞内生物大分子的非细胞质膜损伤机理,本实验对SIF4作用下胞内核酸与蛋白质合成抑制情况和SIF4与胞内核酸靶点作用情况进行分析,以期为将SIF4应用于食源性大肠杆菌的生物防控提供理论与实践参考。
1 材料与方法
1.1 材料与试剂
金抗肽SIF4由本课题组前期研究获得,对大肠杆菌ATCC25922的最小抑菌浓度(minimal inhibitory concentration,MIC)为0.4 mg/L[17];大肠杆菌ATCC25922 上海保藏生物技术中心;4’,6-二脒基-2-苯基吲哚(4’,6-diamidino-2-phenylindole,DAPI) 美国Sigma-Aldrich公司;ATP、pBR322DNA、蛋白酶K、溶菌酶、RNase和溴化乙锭(ethidium bromide,EB)上海生工生物工程(上海)股份有限公司;牛肉膏、蛋白胨、NaCl 国药集团化学试剂有限公司;其他试剂均为国产分析纯。
1.2 仪器与设备
UV-2550紫外分光光度计、RF-5301PC荧光分光光度计日本岛津公司;JASCO J-810圆二色光谱仪 日本分光株式会社;H1850R高速冷冻离心机 湖南湘仪实验室仪器开发有限公司;JY98-IIIDN超声波细胞粉碎机宁波新芝生物有限公司;YXQ-LS-70A立式高压灭菌锅上海博迅医疗生物仪器股份有限公司;琼脂糖凝胶电泳装置 北京六一生物科技有限公司。
1.3 方法
1.3.1 SIF4处理下胞内核酸生物合成情况分析
胞内核酸生物合成分析参考Li Lirong等[21]的方法并略作修改。离心收集对数生长期菌体并用磷酸盐缓冲液(phosphate buffered saline,PBS)(0.2 mol/L、pH 7.2)洗涤3 次,用LB培养基重悬至1×108CFU/mL,加入SIF4至终质量浓度分别为1/2 MIC、MIC和2 MIC,以未添加SIF4为对照组(0 MIC,下同),37 ℃、120 r/min培养24 h,每4 h取2 mL培养液,4 000 r/min离心10 min,菌体沉淀用PBS洗涤3 次并重悬,取重悬液9.85 mL加入0.15 mL DAPI染色液(终质量浓度为15 μg/mL),避光振荡培养10 min,测定DAPI-DNA荧光强度(λex=364 nm,λem=460 nm)和DAPI-RNA荧光强度(λex=400 nm,λem=460 nm)。
1.3.2 菌体基因组DNA的制备
菌体基因组DNA制备参考李婷等[22]的方法并略作修改。将大肠杆菌接种至LB培养基,于37 ℃、120 r/min条件下培养12 h。取10 mL菌悬液5 000 r/min离心10 min,用TE溶液洗涤菌体2 次,5 000 r/min离心10 min,菌体沉淀重悬于4 mL TE缓冲液。加入8 μL 50 mg/mL溶菌酶并于37 ℃温育30 min;加入10 μL 10 mg/mL RNase和0.5 mL质量分数10% SDS溶液,37 ℃温育30 min;加入10 μL 20 mg/mL蛋白酶K,37 ℃温育90 min;加入等体积苯酚-氯仿-异戊醇混合液(25∶24∶1,V/V)混匀,8 000 r/min离心5 min;上清液转入另一支离心管,加入1.8 mL 3 mol/L醋酸钠、18 mL冰无水乙醇,12 000 r/min离心2 min,重复抽提2 次,所获沉淀即基因组DNA。沉淀用体积分数70%乙醇溶液洗涤一次,室温干燥。加入0.5 mL TE缓冲液溶解并测定OD260nm和OD280nm以评价DNA含量和纯度,-20 ℃保藏备用。
1.3.3 SIF4与EB竞争性结合DNA荧光光谱分析
传统上,对于无线点数较少,覆盖范围不广的区域,一般会采用胖无线接入点AP(Access Point),如图1所示。这种方式需要对逐台设备进行配置和管理,无法形成一个整体,无法实现无线网络中的漫游服务功能。这种方式在校园内无线点数比较密集,用户移动性比较频繁的情况下,是不适合的。
DNA荧光光谱分析参考陈旋等[23]的方法并略作修改。将1 mL 50 μg/mL基因组DNA与15 μL 100 μg/mL EB溶液混匀,37 ℃避光温育10 min。加入SIF4至终质量浓度分别为1/2 MIC、MIC和2 MIC,以等体积无菌蒸馏水代替SIF4为对照组。振荡混匀,37 ℃避光温育30 min。用荧光分光光度计检测反应液在550~750 nm波长范围内的荧光强度(λex=535 nm,λem=550 nm)。
1.3.4 紫外吸收光谱分析
紫外吸收光谱分析参考Tang Yali等[24]的方法并略作修改。将50 μg/mL基因组DNA与SIF4混合,使SIF4终质量浓度分别为1/2 MIC、MIC和2 MIC,以等体积蒸馏水代替SIF4为对照组,振荡混匀,37 ℃孵育30 min,于220~320 nm区间进行紫外扫描。
1.3.5 圆二色光谱分析
圆二色光谱分析参考郝刚等[25]的方法并略作修改。移取100 μL 50 μg/ml基因组DNA与SIF4混合,使SIF4终质量浓度分别为1/2 MIC、MIC和2MIC,以未添加抗菌肽的为对照组,37 ℃温育10 min,室温条件下进行圆二色光谱扫描(220~320 nm)。
1.3.6 SIF4处理下胞内蛋白质生物合成情况分析
SIF4处理下胞内蛋白质生物合成情况分析参考郭娟等[26]的方法并略作修改。离心收集对数生长期菌体并用PBS洗涤3 次,用LB培养基重悬至1×108CFU/mL,加入SIF4至终质量浓度分别为1/2 MIC、MIC和2 MIC,以未添加SIF4为对照组,37 ℃、120 r/min培养24 h,每4 h移取2 mL培养液于4 000 r/min离心10 min,菌体沉淀用PBS洗涤3 次,用50 μL TE缓冲液和50 μL溶菌酶重悬,37 ℃温育10 min;取出置冰浴中并加入500 μL冰纯水和500 μL预冷20%(质量分数,下同)三氯乙酸溶液,混匀后冰浴10 min,离心弃上清液,加入1 mL预冷10%三氯乙酸溶液悬浮,离心弃上清液。将沉淀转移至干净试管并加入500 μL 0.15 mol/L的NaCl和5 mL考马斯亮蓝染色液,混匀后于595 nm波长处测定吸光度。
1.4 数据处理与分析
实验结果表示为平均值±标准偏差(n=3),采用SPSS Statistic 25软件对平均值进行方差分析检验,仅进行组内平均值差异多重比较。方差齐性时采用最小显著性差异法进行事后多重比较,否则采用塔姆黑尼(T2)进行多重比较。
2 结果与分析
2.1 SIF4对胞内核酸生物合成的影响
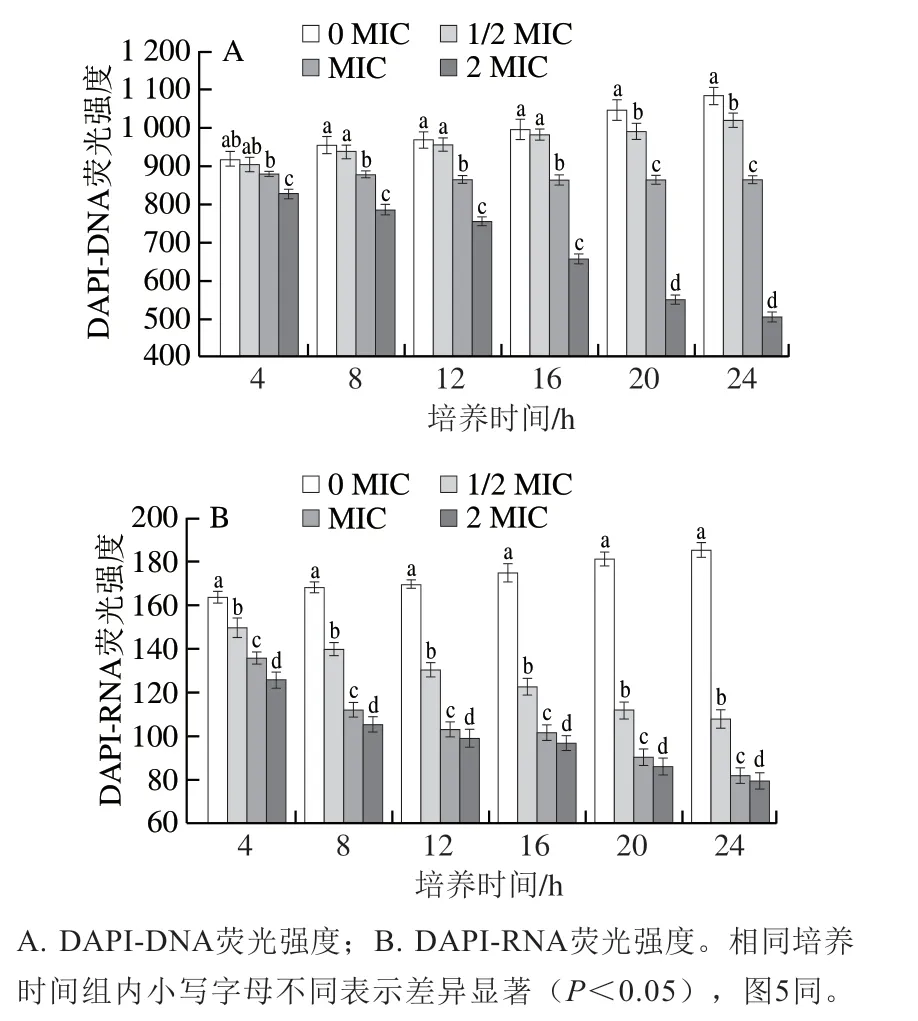
图1 SIF4对胞内核酸生物合成的影响Fig. 1 Effect of SIF4 on intracellular nucleic acid biosynthesis
由图1可看出,各实验组荧光强度均低于对照组,表明经SIF4处理后,大肠杆菌胞内核酸的生物合成均受到不同程度抑制,荧光强度下降与处理剂量和处理时间呈正相关;由图1A可看出,培养4~16 h时,1/2 MIC与对照组荧光强度无显著性差异(P>0.05),继续培养则呈现显著性差异(P<0.05);MIC和2MIC组与对照组均有显著性差异(P<0.05),培养20 h时,相比对照组,1/2 MIC、MIC和2 MIC组DAPI-DNA复合物荧光强度分别下降5.37%、17.43%和47.52%,而培养24 h时,1/2 MIC、MIC和2 MIC组所形成的DAPI-DNA复合物荧光强度相比对照组则分别下降5.96%、20.24%和53.39%,DAPI-DNA复合物荧光强度明显下降说明SIF4能够竞争DAPI和基因组DNA的结合位点,即推断金抗肽SIF4与基因组DNA的结合方式也为沟槽结合,且沟槽结合程度存在剂量效应关系;由图1B可知,实验组DAPI-RNA荧光强度与对照组均有显著性差异(P<0.05)。培养20 h时,相比对照组,1/2 MIC、MIC和2 MIC组DAPI-RNA复合物荧光强度分别下降38.24%、50.04%和52.44%,而培养24 h时,各实验组所形成的DAPI-RNA复合物荧光强度相比对照组则分别下降41.72%、55.79%和57.07%,DAPI-RNA荧光强度显著降低说明金抗肽SIF4可与RNA进行结合。由图1A和图1B可看出,金抗肽SIF4对DAPI-DNA和DAPI-RNA复合物结合的抑制趋势基本相似,推测SIF4可能通过抑制DNA生物合成来影响RNA转录生物量[22],金抗肽SIF4与基因组DNA的结合方式仍需进一步确认。
2.2 DNA荧光光谱分析结果
EB是一种灵敏度高、选择性强的DNA荧光探针,结构中含有一个可嵌入DNA碱基平面的三环平面基团,可通过静电吸附与DNA磷酸骨架结合,或以嵌入方式插入双链DNA碱基平面[21]。当抗菌肽与EB竞争性共存并结合DNA时,EB-DNA体系荧光强度会减弱,说明抗菌肽以类似EB嵌插方式与DNA结合。SIF4与EB竞争性结合DNA荧光光谱如图2所示。
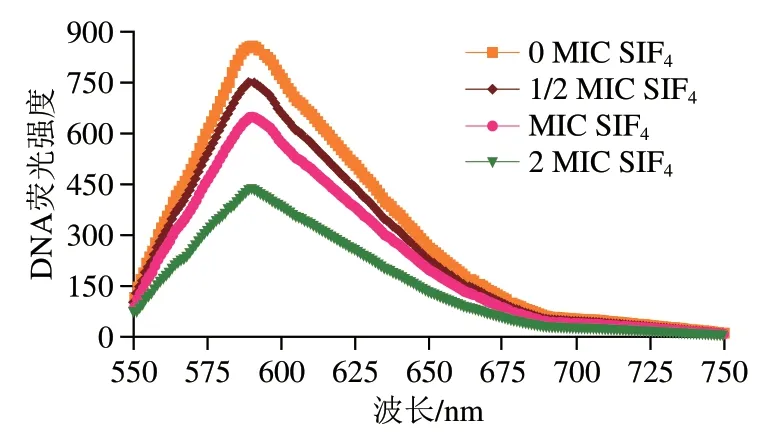
图2 SIF4与EB竞争性结合DNA荧光光谱Fig. 2 Fluorescence spectra of competitive binding of SIF4 and EB to genomic DNA
由图2可看出,EB-DNA复合物的特征性荧光吸收峰在590 nm波长处,随着SIF4质量浓度增加,基因组DNAEB荧光强度存在不同程度衰减,且组间存在显著性差异(P<0.05)。本研究还发现,随着抗菌肽浓度增加,EB-DNA复合物荧光强度虽有不同程度衰减,但复合物发射峰特征性光谱位置并没有发生明显红移或蓝移,说明SIF4添加后并未对EB-DNA体系微环境造成显著影响,只是竞争性地将EB从基因组DNA碱基平面替换下来,从而表现为EB-DNA荧光强度衰减。由此可说明,SIF4可与基因组DNA以嵌插的方式结合。此外,SIF4为阳离子金属抗菌肽[27],结构中正电荷的碱性氨基酸残基可与DNA酸性磷酸基团静电吸附,C-端的两亲α-螺旋疏水基团嵌插至DNA内部疏水沟槽中,从而减弱了EB-DNA体系荧光强度[21]。
2.3 SIF4与基因组DNA互作紫外光谱吸收分析结果
DNA可通过嵌插结合、静电结合及沟槽结合的方式与小分子物质互作并引起吸收光谱红(蓝)移或增(减)色效应,是小分子物质与DNA碱基间π-π体系、偶极-偶极互作及疏水相互作用综合结果[28]。DNA由于含有嘌呤和嘧啶等共轭双键芳香碱基及磷酸生色基团,在近紫外区有强的π→π*电子跃迁,在260 nm波长处呈现强吸收峰,若抗菌肽以静电吸附的方式结合到DNA磷酸骨架,或嵌入到DNA碱基平面,可改变DNA构象并产生减色效应或红移现象,而DNA双螺旋结构若破坏则会产生增色效应,嵌插作用强弱与减色效应呈正相关[15]。SIF4与基因组DNA结合紫外光谱如图3所示。
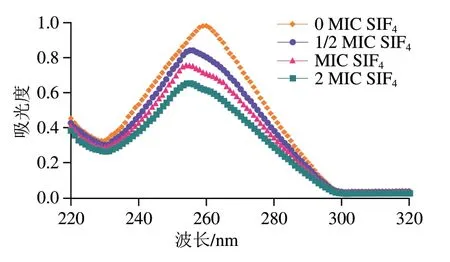
图3 SIF4与基因组DNA结合紫外吸收光谱Fig. 3 UV spectra of SIF4 binding to genomic DNA
由图3可看出,基因组DNA在约260 nm波长处有特征性吸收光谱,各实验组最大吸光度随SIF4处理剂量的增加而呈递减趋势,说明抗菌肽SIF4可与基因组DNA以静电吸附或嵌插的方式结合,使DNA分子构象发生改变并引起减色效应[15]。本研究还发现,随着抗菌肽的加入,SIF4与基因组DNA结合后最大吸收紫外光谱峰由260 nm蓝移至255 nm左右,蓝移了5 nm,说明金抗肽SIF4可能以嵌插方式与基因组DNA结合[29]。由于SIF4与基因组DNA结合并未引起增色效应,说明SIF4与基因组DNA结合并不会导致基因组双螺旋DNA结构断裂,而是通过静电结合或嵌插至DNA碱基平面引起DNA结构的改变,从而影响基因组DNA的紫外吸收光谱[30]。
2.4 SIF4与基因组DNA结合方式的圆二色光谱分析
由于脱氧核糖具有不对称碳原子,其偶极能和碱基不对称地发生互作并显示圆二色性,嘧啶核苷表征正康普顿效应(正圆二色光谱),嘌呤核苷表征负康普顿效应(负圆二色光谱)[31]。小分子物质与DNA互作可干扰螺旋度和碱基堆积的DNA结构,DNA圆二色光谱特征峰常位于230~300 nm。当DNA发生变性时只在约270 nm波长处出现正峰,240 nm波长处峰消失;DNA由B构型向C构型转变时,双螺旋结构变得松散,碱基间堆积力变弱,负峰发生红移[31]。SIF4与基因组DNA互作的圆二色光谱如图4所示。
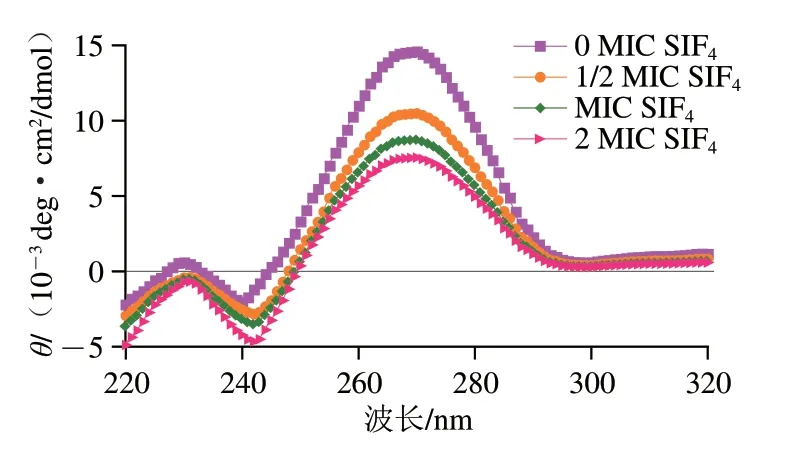
图4 SIF4与基因组DNA互作的圆二色光谱Fig. 4 CS spectra of interaction between SIF4 and genomic DNA
由图4可看出,大肠杆菌基因组DNA呈典型B-DNA结构,270 nm和240 nm波长处出现特征正峰和负峰,270 nm波长处正峰是由碱基堆积力作用形成,240 nm波长处负峰是由双螺旋B型构象形成,与右手螺旋有关[25]。当基因组DNA与SIF4结合后,270 nm波长处正峰强度明显降低,说明基因组DNA与SIF4结合后构象发生改变,削弱了碱基间π-π堆积效应,抗菌肽还与DNA碱基发生沟槽结合,且随着质量浓度增加,沟槽结合互作效果更明显,双螺旋结构变得松散[25]。同时还发现,SIF4与基因组DNA结合后没有引起增色效应,说明SIF4加入只影响DNA二级结构,并未引起双螺旋解链[25];此外,各实验组240 nm波长处负峰强度相比对照组均有一定衰减并发生轻微红移(243 nm),可能是DNA结构由B构型向C型转变以及DNA双螺旋结构疏松导致[25],从而对DNA复制、mRNA转录和蛋白质翻译等产生不同程度的影响。
2.5 SIF4对胞内蛋白质生物合成的影响
胞内物质与能量代谢关键酶、组织蛋白、细胞质膜蛋白及受体蛋白等的主要化学组成为蛋白质,胞内蛋白质合成水平关系着菌体细胞健康状态,监测胞内蛋白质合成水平可表征菌体生存状态。SIF4对胞内蛋白质合成的影响如图5所示。
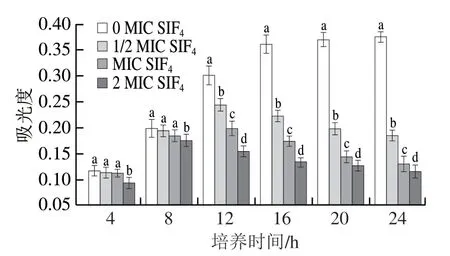
图5 SIF4对胞内蛋白质生物合成的影响Fig. 5 Effect of SIF4 on intracellular protein biosynthesis
由图5可看出,对照组胞内蛋白质水平随培养时间延长而增加,特别是0~16 h内,胞内蛋白质增幅明显,表明菌体蛋白质转录和翻译进程正常;由图5还可看出,在培养4~8 h时,1/2 MIC和MIC组与对照组相比差异不显著(P>0.05),可能是由于抗菌肽作用于菌体时间较短,短时间内细胞膜结构受损较轻,故对胞内蛋白质生物合成影响较少,这与郭娟等[26]的研究结论基本一致;2 MIC组与对照组均存在显著性差异(P<0.05),培养4、8、12、16、20 h和24 h时,相比对照组,2 MIC组胞内蛋白质含量分别下降19.43%、11.62%、48.84%、63.89%、65.73%和69.23%,由此可说明,SIF4对胞内蛋白质合成的抑制效应与处理时间存在明显关系,提示SIF4可能结合DNA和(或)RNA,在转录水平或翻译水平抑制蛋白质生物合成,本课题组前期研究还发现,SIF4可破坏细胞质膜结构并使胞内蛋白质泄漏,从而降低胞内蛋白质含量[17]。
3 结 论
DAPI荧光光谱分析结果表明,SIF4处理后可不同程度地抑制胞内核酸生物合成,抑制效果与SIF4处理剂量和处理时间密切相关。培养4~16 h,低质量浓度(1/2 MIC)SIF4处理对菌体胞内DNA合成影响不显著(P>0.05),MIC和2 MIC组与对照组DNA合成均有显著差异(P<0.05),培养24 h时,1/2 MIC、MIC和2 MIC组DAPI-DNA复合物荧光强度相比对照组分别下降5.96%、20.24%和53.39%,表明SIF4存在竞争DAPI和基因组DNA的结合位点,结合方式为沟槽结合;各实验组DAPIRNA荧光强度与对照组均有显著差异(P<0.05),表明SIF4可竞争性结合DAPI-RNA结合位点,推测SIF4可能通过抑制DNA生物合成影响RNA转录生物量;EB竞争性结合DNA荧光光谱分析结果表明,随着SIF4质量浓度的增加,基因组DNA-EB荧光强度存在不同程度衰减,且组间存在显著差异(P<0.05),基因组DNA-EB复合物特征光谱位置没有发生明显位移,说明SIF4可以通过嵌插方式竞争性替换EB与基因组DNA结合[21];SIF4与基因组DNA互作紫外光谱分析结果表明,SIF4可与基因组DNA发生静电吸附或嵌插结合,使DNA分子构象发生改变并引起减色效应[15,29]。SIF4与基因组DNA结合未观察到增色效应,说明基因组DNA没有发生结构断裂,SIF4只通过静电结合或嵌插至DNA碱基平面引起DNA结构的改变并影响基因组DNA紫外吸收光谱;圆二色光谱分析结果表明,基因组DNA与SIF4沟槽结合后,双螺旋结构未断裂,但构象发生改变,碱基间π-π堆积力减弱[25]。240 nm波长处负峰不同程度衰减并发生轻微红移,提示DNA结构由B构型向C型转变,双螺旋结构变得疏松[25];短时间培养(4~8 h),1/2 MIC和MIC组与对照组胞内蛋白质含量差异不显著(P>0.05),但2 MIC组与对照组均存在显著性差异(P<0.05)。总体来说,SIF4可通过与基因组DNA进行静电吸附或嵌插结合进入到DNA沟槽,影响DNA复制、RNA转录生物量以及蛋白质翻译,实现大肠杆菌非膜损伤抑制。
本课题组前期研究发现,SIF4可破坏细胞质膜结构、细胞质膜通透性与膜生物功能,影响菌体糖代谢、能量代谢及细胞结构与形貌[17-20],其可能是通过破坏菌体细胞壁和细胞膜结构后进入胞内,并影响胞内生物大分子结构与功能从而实现协同抑菌,抑菌作用是否存在先后尚不清楚,需进一步研究确认。为更好地全面解析金抗肽SIF4对大肠杆菌的抑菌机理,本课题组下一步拟在分子生物学层面、转录组学与蛋白质组学层面开展抑菌机理的深层次探究,具体包括:1)研究金抗肽SIF4不同作用剂量条件下对DNA复制相关的rnhA、ssb、dnaG、ligB等基因及DNA损伤修复相关的recA、recN等基因表达水平的影响并进行基因表达分析;同时,采用流式细胞术分析金抗肽SIF4对大肠杆菌细胞周期的影响;2)采用转录组学RNA-seq技术研究金抗肽SIF4作用下菌体胞内mRNA转录调控状态和差异基因,筛选重要差异表达基因,注释这些差异表达基因参与的代谢途径,同时,结合菌体代谢途径验证其基因表达相关功能,理清SIF4处理对菌体代谢相关基因正/负调控机理;3)采用代谢组学技术对金抗肽SIF4处理后菌体内部代谢物种类和含量变化规律进行解析,筛选差异代谢物并进行通路注释分析,结合转录组学结果构建菌体基因-代谢物调控网络图,在代谢组学层面解析菌体内部物质变化、流向及调控规律。
猜你喜欢
科学导报(2024年20期)2024-04-22 09:54:13
现代畜牧科技(2021年9期)2021-10-13 06:38:40
传染病信息(2021年6期)2021-02-12 01:52:08
中成药(2017年12期)2018-01-19 02:06:27
福建轻纺(2017年12期)2017-04-10 12:56:39
广东饲料(2016年5期)2016-12-01 03:43:21
国外医药(抗生素分册)(2016年5期)2016-07-12 14:25:37
上海农业学报(2016年5期)2016-02-10 06:53:40
中国蔬菜(2015年9期)2015-12-21 13:04:38
应用海洋学学报(2015年3期)2015-11-22 07:39:26
