
外源GA3对MdCoL基因及GA相关基因表达的影响
2023-12-04 14:24:16王子昱肖凌云李贵学王鸣枭张玉刚
青岛农业大学学报(自然科学版) 2023年4期
王子昱,肖凌云,李贵学,王鸣枭,张玉刚,4
(1.青岛农业大学园艺学院,山东青岛 266109;2.山东省郓城县林业局,山东郓城 274700;3. 青岛西海岸新区自然资源局,山东青岛 266000;4.东营青岛农业大学盐碱地高效农业技术产业研究院,山东东营 257091)
苹果的树形结构对果树的整形修剪、果园管理和苹果产量都具有重要的影响。柱型苹果树的茎粗壮直立,顶部和底部与短节间的直径几乎没有差异,像一个圆柱体。自1960年在加拿大发现柱型突变体‘威赛克旭’(McIntosh Wijcik)以来[1],其独特的树形一直备受关注。在苹果中,已经发现了许多短枝型突变体。Lapins[2]对‘金冠’(Golden Delicious)× ‘威赛克旭’ 后代进行测交,发现 ‘威赛克旭’ 的柱型生长特性受单一的显性基因Co控制。
根据Koornneef等[3 ]的研究可知,柱型苹果存在的一些表型特征如植株矮化和叶片颜色加深与赤霉素(gibberellins,GA)缺陷突变体的特征相似,因此认为在柱型苹果中内源GA的含量降低是造成柱型苹果节间生长紧凑的关键性因素[3-5]。Looney等[6]对柱型苹果树中GA的含量做了研究,发现在柱型苹果幼苗的茎尖中GA的活性较低。使用硅胶分隔柱色谱法和矮秆水稻生物测定法对‘威赛克旭’茎尖进行了检测,结果表明GA的含量极低且可能不具备生物活性[7];在外源施加GA3后,柱型苹果的幼苗生长量增加,但仍未达到对照的高度,因此可以认为,‘威赛克旭’表型矮化可能与低水平的GA相关。
在高等植物中,GA的合成途径由三个阶段组成。前两个阶段主要包括GA合成基因柯巴基焦磷酸合酶(CPS)和内根-贝壳杉烯合酶(KS),环化后经内根-贝壳杉烯醇(KO)和内根-贝壳杉烯酸(KAO)催化形成GA12醛或GA53,第三阶段中GA12醛被细胞质基质中的赤霉素2-β-双加氧酶GA20ox、GA3ox和GA2ox氧化生成GA12,然后再转化为其他赤霉素。Fukazawa等[8]发现GA信号转导途径中的负调控因子DELLA蛋白GAI(GA-insensitive)和TPR4(TOPLESS-related 4)在转录因子GAF1(GAI-ASSOCIATED FACTOR 1)的介导下结合GA20ox基因启动子,抑制GA的生物合成并参与GA的负反馈调节。GA20ox高表达能够促进GA的生物合成,然而高含量的GA能够通过以上途径反过来抑制GA20ox的表达,从而维持植物体内GA含量的平衡。
为了进一步研究GA对苹果柱型外观的影响,本试验利用外源GA3对柱型苹果‘威赛克旭’和普通型苹果‘旭’新梢进行处理,分析经过处理后柱型苹果和普通型苹果新梢MdCoL基因表达量及GA生物合成相关基因(MdCPS、MdKS、MdKO、MdKAO、MdGA2oxI、MdGA20oxI、MdGA3oxI)以及GA信号传导途径相关基因(MdGID1a、MdRGL2和MdGAI)表达量变化,同时对转MdCoL基因烟草的表型、烟草中GA生物合成及信号转导相关基因表达量的变化进行分析,研究GA与苹果柱型性状之间的关系。
1 材料与方法
1.1 材料
试验材料为青岛农业大学胶州农业科技示范园柱型苹果‘威赛克旭’(McIntosh Wijcik,文中代号为Wi)和普通型苹果 ‘旭’(McIntosh,文中代号为Mc)的新梢。野生型本生烟(Nicotianabenthamiana)及转MdCoL基因的烟草株系CoL-5和CoL-15(本实验室保存)。
1.2 试验方法
1.2.1 GA3对普通型苹果和柱型苹果的处理
选择生长状况一致的普通型苹果‘旭’和柱型苹果‘威赛克旭’,随机挑选新萌发的新梢(2022年5月11日),以100 mg·L-1外源GA3对新梢进行喷施处理,处理程度为表面湿润无液滴滴落即可。分别于GA3处理后的1 h、3 h、6 h、12 h、24 h对新梢取样,以处理前取材作为0 h对照组,重复3次。取样后,将样品在液氮中迅速冷冻,然后放入-80 ℃冰箱保存。
1.2.2 GA3对转MdCoL基因和野生型烟草的处理
以转MdCoL基因烟草和野生型烟草为材料,设置5组重复。用100 mg·L-1GA3对烟草叶片及其生长点进行喷施处理,处理程度为表面湿润无液滴滴落即可,以5%酒精为对照。喷施时间为上午10:00,每隔3 d喷施1次,连续喷施5次。在每次喷施前,测量烟草植株高度和节间长度,测定叶绿素相对含量。节间长度的测量选择自上而下第3~6节,测量后取其平均值(cm);采用SPAD-502便携式叶绿素仪测量烟草顶端3片叶的SPAD值作为叶绿素相对含量。
1.2.3 RNA的提取及RNA的反转录
RNA提取参照北京原平皓生物技术有限公司植物RNA提取试剂盒说明书,对试验材料进行RNA提取。RNA反转录按照Takara生物技术有限公司的反转录试剂盒说明书进行,用反转录后得到的cDNA为模板进行定量试验。
1.2.4 实时荧光定量PCR
实时荧光定量PCR (qRT-PCR)试验的反应体系配制按照SYBR Green PCR Master Mix说明书操作。反应体系配比如下:SYBR Premix Ex Taq Ⅱ(2x)10.0 μL,上、下游引物各0.5 μL,cDNA 2.0 μL,ddH2O 7.0 μL,反应体系共20 μL。所用引物(见表1),生物学重复3次。

表1 qRT-PCR试验引物名称和序列
qRT-PCR测定程序如下:95 ℃预变性30 s、95 ℃变性5 s、60 ℃退火30 s,共44个循环。采用2-ΔΔCT方法计算基因相对表达量。
1.3 数据处理与分析
试验数据均通过Microsoft Excel 2016进行处理;图表均通过GraphPad Prism 8绘制。
2 结果与分析
2.1 外源GA3对苹果MdCoL基因及GA相关基因表达的影响
2.1.1 外源GA3对苹果MdCoL基因表达的影响
以100 mg·L-1的GA3处理柱型苹果春梢,苹果柱形基因MdCoL在苹果新梢中的表达结果如图1所示。GA3处理后,普通型苹果‘旭’和柱型苹果‘威赛克旭’新梢中MdCoL基因表达量变化趋势基本一致,都是在1 h时出现下调,12 h下调到最小,24 h表达又上升,但总体下调表达。除12 h时两种苹果新梢中MdCoL基因表达量差异不大外,其他处理时间MdCoL基因在柱型苹果新梢中的表达量远高于普通型苹果,说明外源GA3处理抑制了MdCoL基因在普通型和柱型苹果中的表达,并且也说明柱型苹果新梢中MdCoL基因表达量高于普通型苹果。
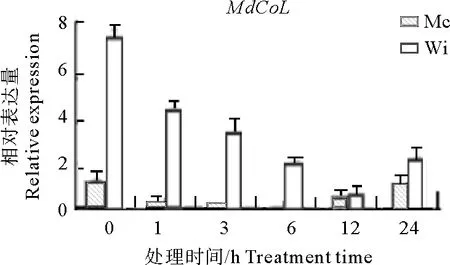
注:Mc(旭)为普通型苹果;Wi(威赛克旭)为柱型苹果,下同。
2.1.2 外源GA3对GA生物合成和信号转导相关基因表达的影响
外源GA3处理后对苹果GA生物合成和信号转导相关基因表达量影响见图2。在0 h时,GA合成途径中的关键基因MdCPS、MdKO、MdGA20oxI和MdGA3oxI以及受体相关基因(MdGID1a)和DELLA 蛋白相关基因(MdRGL2和MdGAI)在柱型苹果中的表达量高于普通型,而其他GA合成基因MdGA2oxI、MdKS和MdKAO的表达量低于普通型。

图2 GA3处理后GA有关基因在‘旭’和‘威赛克’苹果春梢中的表达量
外源GA3处理后MdCPS基因(图2A)在柱型苹果中表达量总体呈下调趋势,而在普通型苹果中,MdCPS的基因表达量在处理6 h时出现显著上调,随后又有所降低;MdKS和MdGA20oxI基因(图2B、2F)在外源GA3处理后在柱型苹果中的表达量总体上呈现出先上升后下降的趋势,两基因分别在3 h和6 h达到最大值,而在普通型苹果中均在12 h达到最高值,随后降低。MdKO基因(图2C)表达量在柱型苹果中处理3 h时达到最大值,随后出现下降,而在普通型苹果中3 h达到最小值,随后逐渐升高。MdKAO基因(图2D)在两种类型苹果中的表达量在处理1 h上调表达,在其他时间均下调表达。MdGA2oxI基因表达量在普通型苹果中的下调趋势非常明显,1 h时只有0 h时的12%,到12 h表达量又升高;而在柱型苹果中则一直缓慢下降,在24 h时表达量仅为0 h的30%,可见外源GA3对普通型苹果新梢MdGA2oxI的影响更为明显(图2E)。MdGA3oxI基因(图2G)在普通型苹果中处理6 h表达量达到最高。
GA受体相关基因(MdGID1a)和 DELLA 蛋白相关基因(MdRGL2和MdGAI)在外源GA3处理1 h后表达量上调,在普通型苹果中,MdGID1a和MdGAI表达量均在处理12 h达到最大值(图2H、2J),MdRGL2(图2I)表达量在处理3 h后达到最高,随后开始下降;在柱型苹果中,MdGID1a表达量在处理1 h达到最大,MdRGL2和MdGAI表达量均在处理3 h达到最大值,随后下降。DELLA蛋白相关基因的上调可能与DELLA蛋白存在反馈调节有关。
2.2 转MdCoL基因烟草中GA相关基因表达量变化
从图3中可以看出,在转基因烟草中GA合成相关基因NbKO和NbGA2oxI以及GA信号传导相关基因NbGAI的表达量低于野生型(图3A、3B、3E);而GA合成相关基因NbGA20oxI和GA信号传导相关基因NbRGL2表达量高于野生型(图3C、3G),说明柱型性状中低含量的GA3可能与高表达的NbGA20oxI和NbRGL2有关。在转MdCoL基因烟草中,可以观察到NbKAO基因表达量变化结果不一致(图3D),在转基因株系CoL-5中的表达量低于野生型,而在CoL-15中高于野生型,NbKAO基因对苹果柱型性状形成的影响还有待研究。NbGAI在转基因烟草中低于野生型(图3E),NbRGL1基因在不同转基因株系中表达趋势不同,在CoL-5中的表达量高于野生型,而在CoL-15中表达量低于野生型(图3F)。

WT为野生型烟草;Col-5和CoL-15分别为转MdCoL基因烟草不同株系。
2.3 外源GA3对野生型和转MdCoL基因烟草表型的影响
野生型和转MdCoL基因烟草以及外源GA3处理对两者表型的影响结果如图4所示。转MdCoL基因烟草与野生型烟草相比,具有植株矮化、节间短、叶片颜色深、叶绿素含量高等特点。外源GA3处理后,野生型和转MdCoL基因烟草(CoL-5和CoL-15)的株高和节间明显增长,转基因烟草的变化更加明显(图4A、4B、4C、4G、4H)。转MdCoL基因烟草在GA3处理3 d后叶片颜色变浅、叶绿素相对含量下降,在6 d后趋于稳定,CoL-5叶绿素含量下降幅度更明显,而野生型烟草叶绿素含量仅有轻微下降(图4D、4I)。从图4F可知转基因烟草在外源GA3处理后开花提前。试验表明,在GA3处理后转基因和野生型烟草均出现了植株高度增加、节间增长、叶绿素含量下降以及提早开花的现象,这些表型变化在转基因烟草中更为明显(图4E、4G、4H),说明转基因烟草对外源GA3的处理更敏感,进一步证明了低水平GA3是影响苹果柱型性状的因素之一。
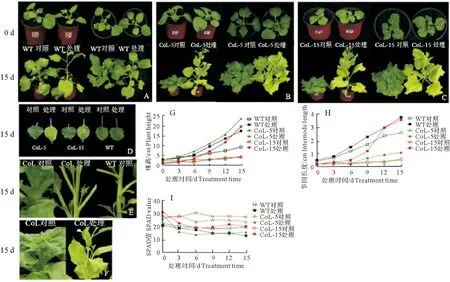
注:WT为野生型烟草;Col-5和CoL-15分别为转MdCoL基因烟草不同株系; A、B、C为野生型和转基因烟草GA3处理0 d和15 d后株型变化;D为处理15 d后野生型和转基因烟草叶片变化情况;E为处理15 d后野生型和转基因烟草节间及分枝数变化情况;F为处理15 d后转基因烟草提早开花;G、H、I为处理15 d内野生型和转基因烟草株高、节间长度和叶绿素相对含量变化情况。
3 讨论与结论
为了进一步明确MdCoL基因是否能通过GA调节来影响苹果柱型性状表型,试验研究了野生型烟草和转基因烟草(CoL-5和CoL-15)的表型区别以及喷施外源GA3后烟草表型的变化,并利用qRT-PCR技术分析了烟草中与GA合成及信号转导相关基因的表达量变化。相比于野生型烟草,转MdCoL基因烟草植株更矮、节间变短、分枝变短、叶片颜色变深、叶绿素含量增多,这些特点和柱型苹果的表型特性相似。喷施外源GA3后转基因烟草出现了植株高度增加、节间增长、叶绿素含量下降等现象,说明GA3对MdCoL基因的调节是影响苹果柱型性状产生的重要因素之一。
对转基因烟草中GA相关基因的表达量进行分析,发现GA合成基因NbGA2oxI和NbKO明显低于野生型烟草,这与田义轲[9]以及白牡丹等[10]的研究结果相同,而NbGA20oxI基因及NbRGL2基因的表达量在转MdCoL基因烟草中含量较高,与柱型苹果中结果相似。研究结果说明在转基因烟草中低含量的GA3能促进NbGA20oxI和NbRGL2基因的表达,抑制了NbGA2oxI基因的表达,从而促进柱型性状的形成,当利用外源GA3处理后,转基因烟草的表型趋于野生型性状,说明柱型性状确实受低浓度的GA3影响。
经外源GA3处理后,苹果柱型基因MdCoL的含量出现明显下调,由此可进一步证明了柱型性状与GA相关。前人研究表明,GA2oxI是GA合成代谢的关键基因之一,可以使植物体内GA的活性降低,从而使植物矮化及节间缩短[11-13]。Thomas等[14]发现GA2oxI在柱型苹果中呈现下调趋势。另一合成基因GA20oxI以及反馈调节GA合成的DELLA蛋白相关基因RGL2在GA3处理前柱型的表达量高于普通型,在处理后两者均有明显上调。GA20oxI高表达促进了GA的生物合成,过多的GA又抑制GA20oxI的表达,二者相互制约,维持植物体内GA含量的相对稳定。因此,柱型性状可能与GA20oxI基因表达量上调促进GA的合成有关;也可能与上调表达的RGL2基因结合到GA20oxI基因启动子上抑制GA20oxI的表达从而减少GA的合成有关。
综上所诉,GA3可以抑制苹果柱型基因MdCoL的表达,二者之间可能存在着一种反馈调节机制来共同调节柱型性状的形成。
猜你喜欢
骨科临床与研究杂志(2022年5期)2022-09-15 01:57:08
舰船科学技术(2022年11期)2022-07-15 07:51:56
煤气与热力(2021年5期)2021-07-22 09:02:14
中国民间疗法(2021年1期)2021-04-20 02:30:40
广东医科大学学报(2020年6期)2020-02-06 06:01:14
西藏农业科技(2019年3期)2019-11-04 00:35:10
汽车与驾驶维修(汽车版)(2019年4期)2019-05-13 03:21:48
现代园艺(2018年3期)2018-02-10 05:18:12
文艺生活·中旬刊(2017年8期)2017-09-15 15:10:53
上海农业学报(2017年3期)2017-04-10 12:39:12
