
孙氏新铺龙(爬行纲:海龙目)一新幼年个体研究
2023-12-01 06:23:54何竞虎艺薇张娣沈越峰
地质论评 2023年6期
何竞,虎艺薇,张娣,沈越峰
1)合肥工业大学资源与环境工程学院,合肥,230009;2)中国地质调查局成都地质调查中心,成都,611230
内容提要:新铺龙是产于我国西南地区的海龙目的一个属,最早发现于贵州省关岭晚三叠世卡尼期小凹组地层。虽然新铺龙材料众多,但其未成年个体骨骼学特征及个体发育模式并未被详细地记述。笔者等报道了一个保存完好的孙氏新铺龙未成年标本,是目前发现最小的完整的孙氏新铺龙个体,标本产自贵州省关岭县新铺乡。依据其体长小于正型标本的一半,头骨长度和躯干比值大,头骨骨缝清晰等特征,判断该标本为孙氏新铺龙的幼年个体。新标本补充了孙氏新铺龙幼年个体的形态学信息,如上颞骨内侧突短于外侧突,翼骨无牙齿发育,神经弓与椎体未融合,尺骨两端微凸,五边形坐骨等。此外,结合已发表的新铺龙标本数据进行统计分析发现,该物种吻部相对头部呈负异速生长,证明了吻部占头骨比例的大小属于新铺龙种内个体发育差异,而并非之前所认为的新铺龙种间差异。
二叠纪—三叠纪生物大绝灭事件后,一些爬行动物在三叠纪早期入侵海洋生态系统,包括鱼龙、鳍龙和海龙等主要类群(Jiang Dayong et al.,2005;Chen Zhongqiang et al.,2012; Kelley et al.,2014)。这其中海龙类是一类起源未知、亲缘关系不明的海生爬行动物(Müller,2005;Liu Jun et al.,2013;Druckenmiller et al.,2020;Chai Jun et al.,2021),广泛分布于北美(东太平洋地区)、欧洲中部(西特提斯地区)以及我国华南(东特提斯地区)等地(Bardet et al.,2014;Motani et al.,2015;孙媛媛等,2020)。
海龙类由海龙超科Thalattosauroidea和谜龙超科Askeptosauroidea组成,包括18个公认的有效种。1904年在东太平洋的美国西海岸加利福尼亚的卡尼阶—诺利阶地层中首次发现了海龙类化石(Merriam,1904)。Merriam命名了4个种属(Merriam,1905),其中两个种属至今依然有效,分别是Thalattosaurusalexandrare和Nectosaurushalius。随后Paralonectesmerriami,Agkistrognathuscampbelli,Thalattosaurusborealis以及Nectosaurushailus陆续被报道(Nicholls et al.,1993;Nicholls,1999)。近年来,Druckenmiller等(2020)在北美地区上三叠统诺利阶地层中又发现了海龙类一新属种Gunakadeitjoseeae。目前东太平洋地区共确认了4属5种的有效性。西特提斯生物区系海龙类的研究始于Askeptosaurusitalicus的首次报道(Nopsca,1925)。此后Hecheleriarubeli,Claraziaschinzi以及Endennasaurusacutriostris相继在圣乔治山动物群等地被发现报道(Rieppel,1987;Müller,2002,2005;Müller et al.,2005)。经过Müller的系统性研究(Müller,2005,2007),目前共确认了3属4种的有效性。20世纪末东特提斯生物区系中晚三叠统地层中发现了大量完整的海龙类化石标本,迄今共确定了8个有效种及1个未定名种属(孙媛媛等,2020):黄果树安顺龙Anshunsaurushuangguoshuensis(Liu Jun,1999;Rieppel et al.,2000;Liu Jun et al.,2005;Maisch,2015),孙氏新铺龙Xinpusaurussuni(尹恭正等,2000;Liu Jun,2001;Liu Jun et al.,2001;罗永明等,2002;Rieppel et al.,2006;瞿清明等,2008),巴毛林新铺龙Xinpusaurusbamaolinensis(程龙,2003),戈氏新铺龙Xinpusauruskohi(Jiang Dayong et al.,2004;Maisch,2014),乌沙安顺龙Anshunsauruswushaensis(Rieppel et al.,2006;Liu Jun,2007),短吻贫齿龙Miodentosaurusbrevis(程龙等,2007;Wu Xiaochun et al.,2009;Zhao Lijun et al.,2010),双列齿凹棘龙Concavispinabiserdens(Liu Jun et al.,2013;赵丽君等,2013),兴义新铺龙Xinpusaurusxingyiensis(Li Zhiguang et al.,2016)和1未正式命名新种(Chai Jun et al.,2021)。
新铺龙属(Xinpusaurus)于2000年由尹恭正等人在我国首次发现报道(表1),模式种为孙氏新铺龙Xinpusaurussuni,最开始被归入鱼龙目杯椎鱼龙科Cymbospondylidae(尹恭正等,2000)。随后一件新的新铺龙头骨化石被详细描述,系统发育分析后重新确定为海龙目的分子(Liu Jun,2001)。程龙(2003)基于一件未完全修复的标本,建立了巴毛林新铺龙。Jiang Dayong等(2004)报道并建立了新铺龙属第三个种戈氏新铺龙。Liu Jun(2013)提出巴毛林种和戈氏种均为孙氏新铺龙的同物异名体,但Maisch(2014)认为戈氏新铺龙为有效种。Li Zhiguang等(2016)在中三叠统拉丁期地层中发现新铺龙一新种,描述并建立了兴义新铺龙。其中吻部长度与头骨长度之比的不同被认为是新铺龙的种间差异(程龙,2003),但随后Liu Jun(2013)对孙氏新铺龙、巴毛林新铺龙和戈氏新铺龙吻部长度与头骨长度之比进行分析后发现吻部占头骨比例随个体发育而变化,据此提出不能以吻部长短作为种间鉴定特征的观点。

表1 已发表的所有新铺龙属标本
新铺龙模式种孙氏新铺龙标本丰富,迄今共报道发表了8件化石标本(表1)。其中尹恭正等(2000)首次发现新铺龙时报道了孙氏新铺龙正型标本Gmr 010及三件参考标本Gmr 011,Gmr 012,Gmr 013,基于以上4具侧视且保存完好的标本建立新铺龙属,确立新铺龙属的鉴定特征。但是该研究仅局限于部分骨骼形态描述和标本数据测量,缺少对骨块间相互接触关系以及肩带和腰带完整的形态解剖学研究。随后Liu Jun等(2001)描述了一个基本完整的侧面保存的头骨及下颌(IVPP V 11860),补充了孙氏新铺龙的头部骨骼的相互接触关系,揭示了颅内骨骼部分形态学特征。此外还依据18个性状展开了支序系统分析,但由于材料不完整,提供的有效性状较少,导致系统发育分析支撑较弱,还需更多的标本补足完整的头骨信息。Liu Jun(2001)描述了一件不完整的孙氏新铺龙标本(IVPP V 12673),包括部分头骨和头后骨骼。文章详细描述了前18节椎体和肩带骨骼的形态学特征,并将已发表标本的肢骨数据整理比对,推测了孙氏新铺龙在水中的运动方式:在运动过程中通过摆动尾巴为身体前进提供动力,四肢主要起舵的作用。遗憾的是该标本仍未提供腰带骨骼信息,且肢骨保存不完整,后肢骨数据不足。GGSr 001由罗永明等(2002)报道,是一件完整、近立体的头骨材料,文章结合了进一步修理的正型标本Gmr 010,对孙氏新铺龙头骨进行了再研究,侧面保存的Gmr 010和腹面保存的GGSr 001作为互补材料,补足了孙氏新铺龙头骨骨骼间接触关系和头骨腹面的形态学特征。但是由于标本保存制约,枕区的骨骼形态学信息还亟待明确。Rieppel等(2006)对头骨侧视保存的一件新标本IVPP V 14372进行了细致的解剖学研究,提供了骨块间更为精确的描述,补足了孙氏新铺龙头骨枕区骨骼形态学信息。
由上可见,孙氏新铺龙已发表的标本众多,虽然头骨骨骼解剖学特征已经研究清楚,但仍缺乏对头后骨骼的深入研究。除此之外,目前尚无明确的孙氏新铺龙幼年个体报道,个体生长发育模式也亟待讨论。笔者等通过描述一件来自贵州关岭地区小凹组保存完整的幼年孙氏新铺龙标本(HFUT GL-17-006),提供了详细的孙氏新铺龙幼年个体形态学信息。除此之外,通过吻部和头骨的异速生长研究,揭示了吻部长度和头骨长度之比的差异是个体发育导致,并不能作为新铺龙种间的差异。
1 系统古生物学
海龙目 THALATTOSAURIA Merriam,1905
海龙超科 THALATTOSAUROIDEA Nopcsa,1928
孙氏新铺龙XinpusaurussuniYin et al.,2000
正型标本:Gmr 010,标本保存完整,长238 cm。
副型标本:Gmr 011。
参考标本:Gmr 012,Gmr 013,IVPP V 12673,IVPP V 11860,IVPP V 14372,GGSr 001,PKU-P-1230。
研究标本:HFUT GL-17-006,保存于合肥工业大学地质博物馆,标本保存完整。
产地和层位:贵州省关岭县新铺乡尧善田西约500 m处上三叠统小凹组,GPS坐标E 105°23′2.82″,N 25°54′16.30″(WGS84坐标体系)。
机构缩写:GMPKU—北京大学地质博物馆(Geological Museum of Peking University,Beijing,China);Gmr和GGSr—贵州省地质矿产局区域地质调查研究院(Geological Survey of Guizhou,Guiyang,Guizhou,China);HFUT—合肥工业大学(Hefei University of Technology,Hefei,Anhui,China);IVPP V—中国科学院古脊椎动物与古人类研究所(Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of Sciences,Beijing,China);XNGM WS—兴义国家地质公园博物馆(Xingyi National Geopark Museum at Wusha,Xingyi,Guizhou,China)。
2 孙氏新铺龙新材料(HFUT GL-17-006)解剖学特征描述
研究标本HFUT GL-17-006头骨为背腹向保存(图1),近立体保存完好,骨缝清晰,长117 mm(吻端—上颞骨后缘),约是正型标本头骨长度的1/3。标本全长约96 cm,在目前所报道的新铺龙标本中个体最小,小于孙氏正型标本的1/2。

图1 新铺龙未成年新标本(HFUT GL-17-006)照片(a)及左侧肱骨(b)和尾椎骨(c)组织薄片取样后制模填充照片,取样位置用黄色方框标记
2.1 头骨
前颌骨:成对的前颌骨细而长,构成新铺龙吻端。不同于参考标本(IVPP V 14372),前颌骨后外侧突构成外鼻孔的前外侧缘,但这与另一参考标本(IVPP V 11860)相似。
上颌骨:上颌骨背缘构成外鼻孔的外侧缘。具上颌齿,右侧上颌齿可见1颗尖端被磨圆的较大的牙齿和2颗细小的尖牙。由于受肋骨挤压,最前端较大的牙齿保存位置向外侧偏移,可能是前人提到的孙氏新铺龙向前倒伏的、较大的上颌骨牙齿(Rieppel et al.,2006)。左上颌骨可见1颗较大牙齿和1颗被肋骨覆盖只有部分露出的细小尖牙。
额骨:额骨成对保存完整,构成大部分颅顶。前内侧突包围了前颌骨的后内侧突。与IVPP V 14372和IVPP V 11860不同的是,额骨前外侧突向前延伸至外鼻孔后缘处。额骨后缘与顶骨间骨缝呈V形,类似于参考标本(IVPP V 14372)的U形骨缝。此外,HFUT GL-17-006额骨与上颞骨无接触。
松果孔位于顶骨中部靠后,区别于参考标本(IVPP V 11860),长明显大于宽,菱形。
上颞骨:上颞骨前缘呈V型分叉,同参考标本(IVPP V 14732,Rieppel et al.,2006)。新标本中内侧突短于外侧突,不同于参考标本(GGSr 001,罗永明等,2002)表现出的外侧突前缘位于顶骨与鳞骨之间。
基枕骨:腹视椭圆形,呈三块凸起状,中心凹,形成枕区的基部,似西特提斯洋生物区系的Hescheleriaruebelia(Rieppel,1987)。
犁骨:犁骨保存完整。前缘与前颌骨接触,外侧缘与上颌骨缝合接触,后缘与腭骨前支缝合接触。由于犁骨前中部被一根粗壮的肋骨覆盖,仅观察到8颗犁骨齿,表面均保存发亮的釉质。自前向后第3对齿是犁骨最大的一对牙齿,可能用于牵制住被捕食的猎物,增加食物在腔内的摩擦(Matsumoto et al.,2017)。
腭骨:腭骨不完全对称,与犁骨、上颌骨组成内鼻孔外缘。末端收缩与翼骨缝合;左侧腭骨外缘呈弧状向内收缩。
翼骨:翼骨前缘宽,中部收缩变厚出现脊状,向后右侧脊消失,末端扩展约与前端等宽。翼骨中部形成脊,翼骨后缘与基蝶骨前缘接触。作为孙氏新铺龙重要特征的翼骨齿,并没有在此标本上发现,这一性状不同于已发表的新铺龙属成年个体标本(罗永明等,2002;Jiang Dayong et al.,2004;Rieppel et al.,2006;Li Zhiguang et al.,2016)。
舌骨:在新标本中仅观察到左侧舌骨,舌骨原位保存。腹视舌骨前部粗壮,后部细长。
外翼骨:外翼骨位于翼骨后。左侧外翼骨暴露较多,内缘与翼骨接触。
齿骨:齿骨保存完好,侧面暴露(图2)。向后延伸在上隅骨上下两侧分成两支,与上隅骨V型缝合接触。左侧与右侧齿骨各具7颗牙齿,左侧第1、4颗及右侧第1、2颗为锥状齿,其余为球状。齿骨齿数量少于参考标本(GGSr 001,左侧齿骨具15颗齿)(罗永明等,2002)。

图2 贵州关岭地区孙氏新铺龙标本(化石编号:HFUT GL-17-006)的头骨背视照片(a)、头骨背视骨骼解剖素描图(b)、头骨腹视图(c)及头骨腹视骨骼解剖素描图(d)
冠状突明显,左侧冠状突隆起较高似三角形,右侧冠状突似矩形。
上隅骨:上隅骨前缘与齿骨V型缝合,与正型标本一致,但不同于参考标本(IVPP V 14372和IVPP V 11860)倾斜状的骨缝。
2.2 头后骨骼
HFUT GL-17-006头后骨骼保存较好(图1)。通过神经棘和椎体判断脊椎数不少于110节,其中约34节荐前椎,寰椎被头骨和颈肋覆盖,未完全暴露,仅见其与枢椎紧密相连。寰椎神经棘较直立。枢椎侧视圆筒状,神经棘较高(图3a)。椎体前后端无扩展,区别于正型标本(Gmr 010)。颈肋与正型标本一致,单头型,自近端向远端收缩变细。
荐前椎椎体圆筒状,中部明显下凹。椎体高大于长,椎体高度约是椎体长度的1.5倍,与Gmr 010一致。相邻的神经棘排列紧密,间距较参考标本(IVPP V 12673)小,荐前椎神经棘微微向后倒伏,倒伏程度逐渐变缓至直立,其中第19节背椎神经棘最高。肋骨单头型,长度约86 mm,自近端向远端逐渐收缩变细。腹肋极细,毛发状。
荐椎椎体高大于长,椎体横突较背椎椎体横突低。第1至第3节荐椎神经棘均高于背椎神经棘和尾椎神经棘,并逐渐增高,第4节荐椎神经棘最矮且矮于部分背椎(表2)。荐肋较背肋粗且短,均未与椎体融合。肋骨远端扩张。

表2 HFUT GL-17-006 标本部分骨骼数据
尾椎长,末端微微上扬,保存状态与正型标本(Gmr 010)不同(末端微微下弯)。约72节尾椎,椎体高略大于长,自近端向远端椎体高度逐渐近于长度。第1节尾椎神经棘约是第4节尾椎神经棘2倍宽,自第5节尾椎起神经棘呈背腹向倒伏,与正型标本(Gmr 010)存在差异的是末端尾椎神经棘明显增高且更宽。笔者等研究标本保存典型Y型脉弓。
肩带:背视暴露,包括锁骨,间锁骨,肩胛骨和乌喙骨。其中右侧保存完整,左侧未见肩胛骨和乌喙骨(图3)。锁骨保存良好,细长弯曲状骨骼,远端收缩变细,与参考标本(IVPP V 12673)相区别。左侧锁骨近端和远端骨杆弯曲约130°,与参考标本(IVPP V 12673)相似(Liu Jun,2001)。间锁骨十字形,有明显的短的前突,后突长且自近端向远端收缩变细。肩胛骨细长,长度与肱骨相近,未见关节窝似结构,左侧肩胛骨近缘几乎平直,与参考标本(IVPP V 12673,Liu Jun,2001)相同。乌喙骨具乌喙骨孔,近圆形,与正型相同(Liu Jun,2013,Fig.1),但是不同于参考标本(IVPP V 12673)拱形的乌喙骨(Liu Jun,2001)。
腰带位于整件标本的中前部,整体比肩带骨骼大,左侧腰带暴露,右侧被肋骨覆盖难以辨别。髂骨微微弯曲,近端较远端窄,在外侧缘1/4处形成凸缘,向后从1/4至1/2处形成凹缘。自中部至远端扩展形如矩形。耻骨被腹肋覆盖,远端连接坐骨。坐骨呈不规则五边形,区别于正型标本(Gmr 010)圆三角形的坐骨(尹恭正等,2000)
肱骨棒状,近端宽度大于远端宽度的2倍(表2),与已发现的新铺龙标本形态一致(Liu Jun,2001)。桡骨肾形,两端平直无凸缘,近端厚于远端,内侧缘微微内凹,外侧凸缘明显,最宽处位于桡骨前1/3处。尺骨棒状,不同于参考标本(IVPP V 12673,Liu Jun,2001),两端均形成轻微凸缘。新标本尺骨近端较远端扩展明显,中部收缩,同时外缘收缩较内缘收缩强烈。左侧股骨保存良好,完全暴露,远端宽近端窄。约2/3处向后扩展形成宽大的远端。右侧股骨前中部被左髂骨和荐肋覆盖,只可见扩展的远端,远端宽与左股骨远端宽相近。胫骨同正型标本(Gmr 010)相似,远端较近端窄,中部向内收缩,内侧缘收缩较外侧缘明显(尹恭正等,2000)。近端宽约为远端宽的2倍。胫骨长度约是股骨长度的1/2。腓骨近端被胫骨覆盖,似扇形,与正型标本(Gmr 010)和参考标本(IVPP V 12673)扇形腓骨相似。附肢骨异位保存,约有3块棒状细小骨骼,推测可能是趾骨。
3 讨论
笔者等的研究标本(HFUT GL-17-006)具有孙氏新铺龙的一些典型形态学特征,包括吻突尖且细;三射状轭骨;上颌骨前端牙齿扩大;犁骨具齿;齿骨前部牙齿椎状,后部牙齿球状;下颌细长,齿骨联和部窄;颈椎数量小于7节;肱骨近端宽,远端窄;桡骨肾形;背椎神经棘向后逐渐增高且顶部无内凹;尾椎长且分节多,神经棘背腹向倒伏,因而可鉴定为孙氏新铺龙。
但和正型标本相比,研究标本体长不及正型标本的一半,头骨长度约是正型标本的1/3。头骨长度和躯干(肩臼—髋臼)的长度之比也比正型标本更大,反而与乌沙安顺龙的一个未成年个体标本KM512相近(前者为0.43,后者为0.4,Liu Jun,2007)。同时,研究标本的头骨骨缝清晰,且神经弓与椎体未融合。上述特征都表明研究标本是未成年个体(Currie et al.,1984;Carroll et al.,1985;Rieppel,1993;Maisch,2015)。
笔者等的研究标本和孙氏新铺龙成年个体相比,除上述较为明显的差异外,还有如下可能因个体发育阶段不同导致的差异,如:头骨中,翼骨齿作为新铺龙属重要的鉴定特征之一(Liu Jun et al.,2001;Jiang Dayong et al.,2004;Rieppel et al.,2006;Li Zhiguang et al.,2016),在研究标本中却并未发育翼骨。同时翼骨上也未发育脊状构造。这与黄果树安顺龙的一个未成年个体相似(Maisch,2015)。上颞骨与笔者等的参考标本(GGSr 001)不同,研究标本表现出前缘V型分叉,内侧突短于外侧突。左右冠状突形态不一,左侧顶端尖而右侧平直。
头后骨骼中,研究标本肩带处锁骨远端明显收缩变细,而笔者等的参考标本(IVPP V 12673)锁骨远端无明显收缩。乌喙骨不似参考标本(IVPP V 12673)的拱形,而是近圆形,同时未成年的研究标本具乌喙骨孔且关节窝位于背腹侧。腰带部分正型标本的坐骨是圆三角形,研究标本却表现出不规则的五边形。肢骨中,研究标本尺骨近端与远端均形成凸缘,尤其近端凸缘明显,不同于两端形态不同的参考标本(IVPP V 12673)。参考标本(IVPP V 12673)桡骨由近端向远端扩张再收缩,最宽处出现在桡骨中部,而研究标本桡骨最宽处位于距近端1/3处,较参考标本(IVPP V 12673)更靠近端。股骨由近端向远端先轻微收缩变细再扩张,远端形成凸缘,但收缩程度较参考标本(IVPP V 12673)弱。正型标本(Gmr 010)具100节尾椎,而研究标本约72节尾椎,椎体长度约是正型标本的1/3。这些差异是否为个体发育所致需要更多幼年标本的研究来确认。
前人对于孙氏新铺龙的研究中,关于不同个体吻部占头骨比例的差异究竟是属于种间差异还是个体发育差异这个问题仍有较大争议(程龙,2003;Jiang Dayong et al.,2004;Liu Jun,2013)。为了解决这一问题,笔者等收集了头部骨骼完整且已发表的七件新铺龙标本数据(尹恭正等,2000;罗永明等,2002;程龙,2003;Jiang Dayong et al.,2004;Liu,2013),并依据标本大小对这些数据进行了排序(表3)。笔者等利用线性回归的方法探究吻部和头骨在个体发育中生长速度的关系,采用最小二乘法,因此将回归线定义为
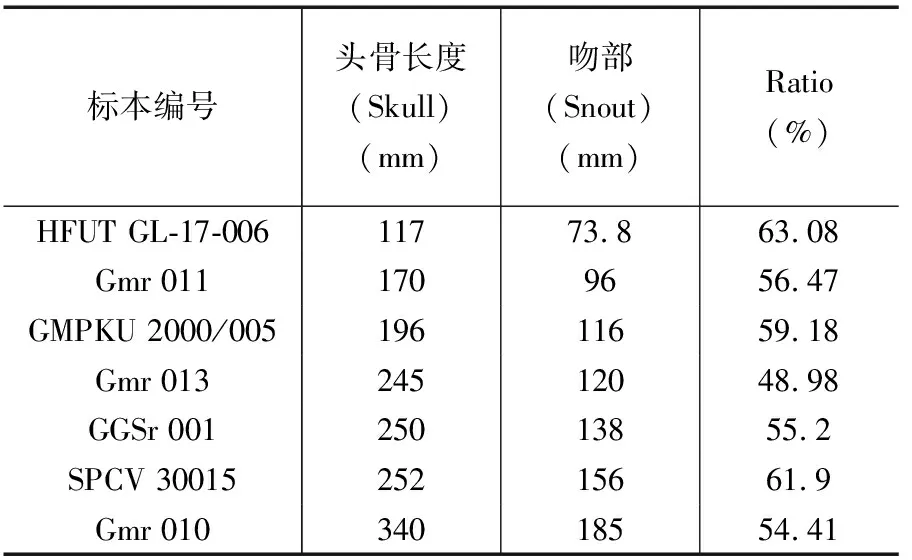
表3 新铺龙属头骨测量数据
lgy=b+klgx
其中x表示头骨长度,y表示吻部长度。置信区间设置为95%,计算中不包括误差项的总和(Russell,1970;Currie,2003;Farlow et al.,2005)。为了避免极端值的影响,在分析之前对测量数据进行了取对数处理。
分析结果显示k= 0.86,判定系数R2为0.94,其中P= 0.0002,呈显著差异。k小于1表明新铺龙吻部相对头骨呈负异速生长(图4),即随着个体的生长,吻部生长速度比头骨整体的生长速度慢,因此吻部相对头骨的比例逐渐变小。分析结果支持新铺龙吻部占头骨的比例主要受个体发育影响,不能作为新铺龙种间差异的证据。
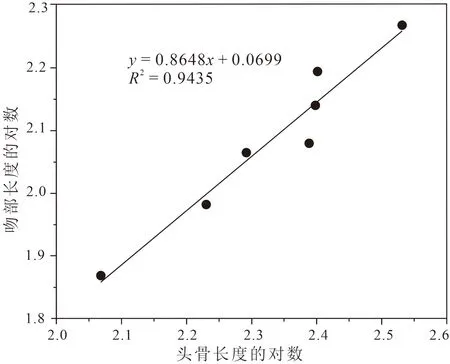
图4 头骨长度对数与吻部长度对数异速生长示意图
4 结论
笔者等通过对孙氏新铺龙幼年个体与成年个体形态学特征进行对比,提供了孙氏新铺龙幼年个体的性状特征,如:头骨上颞骨内侧突短于外侧突,翼骨无齿,头后骨骼中神经弓和椎体没有愈合,约72节尾椎,锁骨远端明显收缩变细,乌喙骨近圆形且具乌喙骨孔,坐骨呈五边形,神经弓和椎体没有愈合,附肢骨部分肢骨上尺骨两端均呈凸缘且近端更明显等特征,为孙氏新铺龙的后续研究提供依据。对新铺龙属吻部发育进行线性分析,发现呈现明显的负异速生长模式,因此认为吻部占头骨比例的大小不能作为新铺龙属种内区分的鉴定特征,研究结果可以为新铺龙属种厘定提供新的依据。
致谢:感谢李良友和佐藤哲哉(Tetsuya Sato)对标本的修理,合肥工业大学古生物实验室其他成员在野外及实验室内的帮助,刘俊教授和Andrzej S.WOLNIEWICZ博士对标本的指导讨论,以及刘俊教授对论文全文的详细修改。感谢中国科学院古脊椎动物与古人类研究所刘俊研究员所提出的宝贵评审意见。
猜你喜欢
环球时报(2022-11-10)2022-11-10 17:18:50
中老年保健(2021年5期)2021-12-02 15:48:21
杭州师范大学学报(社会科学版)(2021年5期)2021-11-01 05:53:08
中老年保健(2021年5期)2021-08-24 07:06:28
奥秘(2020年7期)2020-08-02 11:00:27
山海经(2017年15期)2017-09-03 10:21:31
山海经·故事版(2017年8期)2017-08-22 23:47:04
小布老虎(2017年1期)2017-07-18 10:57:27
百家讲坛(2016年10期)2016-07-04 18:28:46
意林(2010年19期)2010-10-22 11:57:12
