
融合物联网多环境参数的茎干水分SSA-BP 预测模型
2023-11-26 10:12许泽海赵燕东
农业工程学报 2023年16期
许泽海,赵燕东
(北京林业大学工学院,北京 100083)
0 引言
对于多数植物来说,茎干组织向上连接着冠层枝叶,与大气进行水汽交换并传递冠层固定的光合产物,向下连接着根部,将土壤水分与无机盐传输至植物重要的组织[1-2],此过程需耗费一定时间,有时会导致用于冠层蒸腾的水分无法及时补充,随着植物蒸发需求的继续增加,可能会使木质部导管发生栓塞及空穴化,此时茎干组织会起到一定的缓冲作用,将自身储存的水分传输至冠层枝叶及其他组织,维持组织细胞代谢、光合固碳及蒸腾耗散等生理活动的正常运行[3-4]。也有研究表明当植物面临干旱、冻害、热及病虫害胁迫时,当其外在形态没有受到影响之前,茎干水分就已产生较明显变化[5]。同时,茎干水分在环境参数如空气温度、光合有效辐射、风速、饱和水汽压差等的非线性叠加作用下会呈现出不同变化特征[6-7]。可见,对不同时期茎干水分的准确预测对理解植物体内水分运移、平衡及其环境适应机制等具有重要意义[8]。
近年来,神经网络算法如BP 神经网络(back propagation neural network)、长短期记忆(long short term memory,LSTM)网络及支持向量机(support vector machine,SVM)等算法及其优化算法因可以很好地解决复杂且非线性建模问题得到了迅速发展[9],在农林业相关预测研究中也受到了很大的关注[10-12]。TU 等[13]利用BP 神经网络实现了对复杂、非线性且有不确定过程的茎流很好的预测;高鹏等[14]采用基于物联网和Deep-LSTM 模型实现对不同胁迫程度下的茶树净光合速率有效且快速预测;LI 等[15]设计了基于液流预测的CGRU(convolutional neural network-gated recurrent unit)模型,其效果较常见预测模型有了很大的提升。考虑到神经网络收敛速度慢、易陷入局部最优及过拟合等问题,多数研究者提出生物启发式算法如遗传算法(genetic algorithm,GA)[16]、粒子群优化算法(particle swarm optimization,PSO)[17]、灰狼优化算法(grey wolf optimizer,GWO)[18]、麻雀搜索算法(sparrow search algorithm,SSA)[19]等算法优化神经网络极大改善了模型预测精度及收敛速度。如ZHU 等[20]采用遗传算法优化后的BP 网络结合多传感器数据,较好地实现了对农用小车轨迹的预测;迟宇等[21]提出模拟退火的粒子群算法SA-PSO-LASSO-BP(simulated annealing-particle swarm optimization-least absolute shrinkage and selection operator-back propagation)神经网络模型,实现了对猪舍环境质量评价。祖林禄等[22]提出基于SSA 优化LSTM 的温室环境预测模型,实现了对6种环境参数的准确预测。
目前关于植物茎干水分预测研究还鲜有报道。为对生长季茎干水分变化特征进行解析,并构建茎干水分预测模型,本研究选取五角枫为研究对象,采用具有自主知识产权的茎干水分传感器获取茎干水分变化情况,同时监测各环境参数,搭建五角枫物联网生态信息监测系统;结合植物生理生态学知识分析生长季不同时期茎干水分变化特征;采用具有较好收敛速度和搜寻能力的麻雀搜索算法(sparrow search algorithm,SSA)优化BP神经网络即SSA-BP 模型对茎干水分进行预测,并与传统的BP 模型及GA-BP 模型的预测精度进行比较,以期为探究植物内部水分平衡及为林木水分管理提供技术指导与支撑。
1 材料与方法
1.1 研究材料
研究区域位于北京市海淀区北京林业大学三顷园苗圃,地处116°20′E,40°00′N,海拔约为50 m,园区内的土壤为黏壤土,pH 值在7~8 之间,气候属半湿润大陆性气候,全年日照时数在2 560 h 左右,年平均蒸散量在1 800 mm 左右。
在2022 年4 月初选择12 颗树龄为5 a 且平均胸径为5.2 cm 的五角枫,移栽于直径为85 cm 高为46 cm 的花盆中,人工定期对五角枫进行浇水及除草。
1.2 茎干水分传感器
采用自主设计的基于驻波率原理的茎干水分传感器(0~2 500 mV,±2%)[23],实现对茎干水分变化无损且实时采集。该传感器主要由100 MHz 信号源、50 Ω 同轴传输线、检波与放大电路、304 不锈钢探测环和保护外壳组成,测量原理如图1 所示。

图1 茎干水分传感器测量原理图Fig.1 Schematic of stem water content measurement principle
高频信号源产生的电磁波经传输线传送至探测环,如探测环阻抗改变,传输线上各点电压幅值会发生变化,通过检波电路获取a、b两点的差动信号,经放大电路得到传感器输出电压Uout[24]:
式中β为放大器系数;A为信号源的幅值,V;Z0为传输线阻抗,Ω;Zl为探测环阻抗,Ω;A和Z0均为常数,因而Uout由Zl决定。
研究表明[25],茎干组织中水分含量变化会导致Zl发生变化,因而Uout发生变化,测量Uout即可检测出茎干组织中的水分含量[26]。团队前期多次试验表明[3-4,25-28],Uout与茎干水分真实值的线性拟合决定系数均在0.96 以上,为极强相关。故本研究茎干水分采用Uout来表征[28]。此外,前期试验表明实际中的环境温湿度和电导率对传感器运行的扰动很小[27-29]。综上所述,该传感器可稳定运行,获取的数据可靠且可用来表征茎干水分。测量时,将传感器安装于五角枫胸径处即距地面约1.3 m 高处。
1.3 五角枫物联网生态信息监测系统搭建
为实现对监测期间五角枫茎干水分数据及各环境参数的实时采集与在线展示,本研究搭建了五角枫物联网生态信息监测系统,该系统主要由各环境参数传感器、茎干水分传感器、数据采集器、DTU 模块(data transmission unit,DTU)、数据服务器、监测平台等组成,如图2 所示。在栽种五角枫的附近区域安装各环境参数传感器,其中BYX 气象百叶箱,用于监测环境中的空气温度(T,℃)、相对湿度(RH,%)及二氧化碳(CO2,μmol/mol)信息;SL3-1 雨量传感器用于采集降雨量(RF,mm);GHFS 光合有效辐射传感器用于监测光合有效辐射强度(PAR,W/m2);EL15-1A 风速传感器获取监测期间的风速(WS,m/s);HYSWR-ARC 土壤水分传感器用于获取15 cm 深处土壤含水率信息(SM,%);DS18B20 温度传感器用于获取15 cm 深处土壤温度信息(ST,℃)。
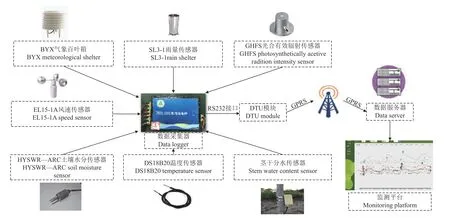
图2 五角枫物联网生态信息监测系统Fig.2 Acer mono Internet of Things ecological information monitoring system
将各气象传感器及茎干水分传感器均连接于实验室自主设计的基于ATMEGA2560 单片机的ZRDL 1 001 型多通道数据采集器上,对研究期间茎干水分及各环境参数进行获取,监测期间设置每10 min 采集一次各传感器数据,一天共采集144 组数据,各传感器数据存在采集器SD 卡内。
为实现对各传感器数据实时在线显示并及时了解各传感器运行情况,将装有SIM 卡的DTU 模块接于数据采集器232 通道,采用GPRS 网络将打包后的各传感器数据通过DTU 模块传输至远程阿里云数据服务器,阿里云物联网平台通过TCP 协议与DTU 模块建立连接,接收数据包存储于数据服务器端的MySQL 数据库,同时将数据包发送至监测平台,并通过TCP 协议连接访问服务器,接收数据服务器中储存的各传感器数据包,对数据包进行解析和处理,展示并绘制监测期间各传感器数据的动态变化情况,从而实现对茎干水分和各环境参数实时查看与数据下载。由此完成了五角枫物联网生态信息监测系统的搭建,从2022 年4 月17 日开始对生长季五角枫茎干水分及各环境参数进行持续监测。
此外,本研究引入饱和水汽压差(vapor pressure deficit,VPD),该参数是利用空气温度和相对湿度通过式(2)计算得到[30]:
式中VPD为VPD,kPa;a1为空气温度,℃;b1为相对湿度,%。
1.4 数据处理
采用Microsoft Office Excel 2019 对监测系统得到的茎干水分数据和环境参数进行整理和计算。
采用MATLAB 软件中的滤波算法剔除茎干水分及环境参数中因环境扰动、人为操作失误等因素产生的异常数据及数据中存在的噪声。
将处理后的茎干水分数据与同时刻获取的环境参数进行组合,作为后续茎干水分预测模型的数据集,其中将60%的数据集作为训练样本,40%的数据集作为测试样本,用于后续模型的训练与测试。本研究选取T、RH、PAR、WS、ST、SM、VPD 共7 个环境参数作为输入变量,以茎干水分作为输出变量构建预测模型进行训练和预测。
为消除各数据间存在量纲对模型拟合带来的负面影响,提高后续预测模型训练的精度和速度,将模型的输入输出数据归一化至[0,1],采用MATLAB 中的mapminmax 函数归一化处理,如式(3)所示:
式中Xmax和Xmin分别为待处理样本数据的最大值和最小值;X1为归一化后的值;Xx为待处理的样本数据。
1.5 预测模型构建
1.5.1 遗传算法优化BP 神经网络
BP 神经网络是将误差反向传播对网络的权值和阈值进行不断调整提升模型预测精度。作为局部搜索的优化方法,对权值和阈值的敏感度高,不同的初始权重和阈值会使网络收敛于不同的局部极小,陷入局部最优;此时权值迭代将停止,如果有多个局部最优,就难以确定最后得到的权值是否为最优[31],导致模型预测效率和稳定性较难满足实际的需求。
本研究采用GA 来优化BP 神经网络从而对生长季不同时期的茎干水分进行预测,GA 采用全局寻优得到最优解,根据遗传学原理,在优化过程中通常会设置一个固定规模的初始种群,其中种群中的每个个体都表示问题的一个可能解,利用适应度函数判别个体适应环境的程度,其中适应度较好的个体会留下来继续繁衍,在此过程中经过选择、交叉、变异等操作形成新的种群[32],把GA 优化后的初始权值和阈值赋值给BP 神经网络进行学习更新,如此反复得到最优参数并对BP 神经网络的初始权值和阈值进行赋值,得到较好的预测效果。
1.5.2 麻雀搜索算法优化BP 神经网络
本研究引入XUE 等[19]受麻雀种群觅食的行为启发提出的 SSA 优化算法,具有迭代次数少、有较快收敛速度和较强的搜索能力,大幅削减了模型训练的时间;有研究表明SSA 在局部及全局的寻优能力、搜索精度、稳定性及收敛性能等方面要更优于前述GA、PSO 及GWO等生物启发式算法,可更有效避免算法陷入局部最优,进而可有效提升后续预测模型的性能[33]。
此算法模拟了麻雀种群的觅食过程,把种群分成发现者、跟随者和侦查预警者,其中发现者为整个群体提供觅食方向和区域,跟随者主要是跟随发现者觅食,侦察预警者一旦意识到有危险发生,立即发出预警信息,种群将出现反捕食行为。
一般具有n只麻雀的种群可以表示为
式中X表示麻雀种群;d为待优化变量的维数;n为麻雀种群的数量;表示第n只麻雀的第d维位置。
所有麻雀的适应度函数(Fx)由式(5)得到:
式中f(x)为适应度值。其中拥有较好适应度值的发现者会在搜索过程时会率先得到食物,为麻雀种群寻觅食物并向加入者提供觅食方向,因而发现者拥有更广阔的觅食空间。
每次迭代时,发现者通过式(6)进行位置的更新:
式中t为当前迭代次数;j=1,2,3…d;Xij表示第i只麻雀在第j维中的位置信息;α(α∈[0,1])为随机数;C表示最大迭代次数;R2(R2∈[0,1])为预警值;ST为安全值;Q表示服从正态分布的随机数;L表示内部元素均为1大小1×d的矩阵,其。当R2<ST时,表示没有入侵者,可以执行搜索操作,相反,表示一些麻雀发现入侵者发出预警,迅速飞离。
跟随者执行式(6),监视发现者同时准备与之竞争,否则按式(7)更新位置:
式中Xp表示发现者所处的最佳位置;Xworst则为最差的位置;A为大小1×d的矩阵,各元素均在{-1,1}中选择,A+=AT(AAT)-1;i>n/2 表明适应度值较低的第i个加入者未获取到食物,并处于饥饿状态。
侦察预警者的位置按如下规则进行更新:
式中Xbest表示当前最优位置;β作为步长控制参数;fi、fg和fw分别为当前适应度、最优和最差适应度;K作为[-1,1]中的随机数,ε是避免分母为0 的常数。
为避免BP 神经网络易趋于局部最优,提升模型的预测效果,采用有着较强大的局部、全局搜寻能力及快速收敛能力的SSA 对BP 神经网络的初始权重和阈值进行更新优化,使种群朝全局最优移动,更好地解决了BP 神经网络存在的劣势;随后将优化后的权值和阈值赋于BP 神经网络,进而完成对BP 神经网络的优化,最后完成对生长季不同时期茎干水分变化趋势的较准确预测。算法流程如图3 所示。具体步骤如下:
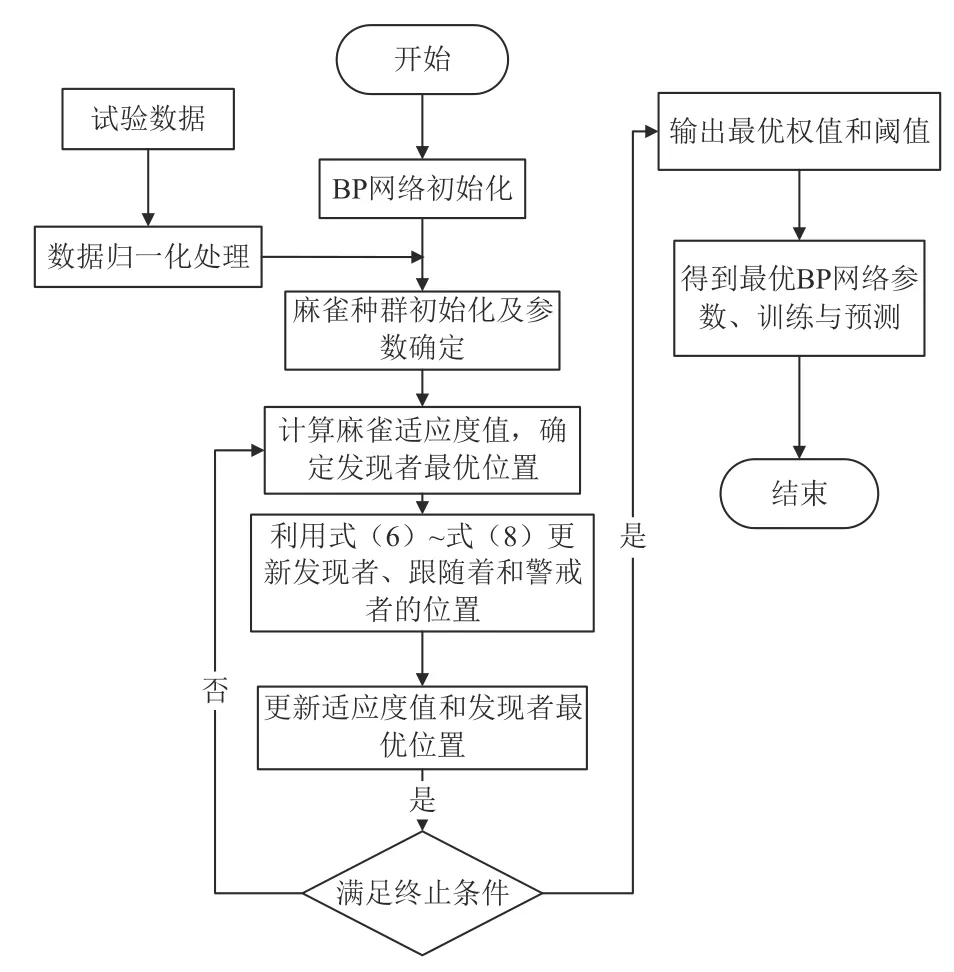
图3 SSA-BP 神经网络流程图Fig.3 Flow chart of SSA-BP neural network
1)初始化麻雀种群数量、初始位置、最佳位置、最佳适应度、最大迭代次数及BP 神经网络结构等参数。
2)对茎干水分以及各环境参数数据进行预处理。
3)利用式(6)~式(8)对发现者、跟随者和侦查预警者的位置进行更新。
4)更新最佳适应度,判断是否达到设定的误差和最大迭代次数,如满足,执行下一步,否则,返回步骤3)。
5)BP 神经网络训练,将 SSA 优化后的最优权值和阈值赋予 BP 神经网络,完成茎干水分的训练和预测。
1.5.3 各模型参数设置
试验中,设置BP 神经网络模型为3 层,分别使用tansig 和purelin 函数作为输入层至隐藏层及隐藏层至输出层的激活函数,最大训练次数为1 000 次,学习率设置为0.01,训练精度为10-6;本研究将7 个环境参数作为输入,茎干水分作为输出,因而设置模型输入节点数为7,输出节点数为1,隐藏层的节点数通过式(9)确定[34]:
式中h为隐藏层节点数,n2为输入节点数,m为输出节点数,a2是1~10 的常数,因此隐藏层节点数在4~13范围内,经试凑法设置隐藏层节点数为7。
设置GA-BP 算法的迭代次数为50,种群规模为30,采用轮盘赌法进行选择,交叉概率为0.4,变异概率为0.05。
设置初始麻雀数量为30,最大迭代次数为50,阈值和权值的上下边界值分别为5,-5;发现者占麻雀种群的20%,安全值设为0.8,通过式(10)得到麻雀种群维度数(搜索空间维度)d1为64:
1.5.4 模型评价指标
为直观地评价SSA-BP 模型预测能力,本研究采用决定系数(coefficient of determination,R2)、均方根误差(root mean square error,RMSE)和平均绝对误差(mean absolute error,MAE)作为BP 模型、GA-BP 模型及SSA-BP 模型预测性能的评价指标,计算式见文献[35]。
2 结果与分析
2.1 生长季各环境参数的变化特征
图4 为本研究搭建的物联网生态信息监测系统获取的生长季各环境参数包括土壤含水率、风速、降雨、光合有效辐射、空气温度、相对湿度、饱和水汽压差及土壤温度的变化特征。监测期间土壤含水率在补水后呈显著上升趋势,随后逐渐降低,其波动范围在31.4%~68.8%;风速在春秋两季波动较为明显,夏季波动范围较小,在0~35.2 m/s 范围内波动;降雨集中在6—9 月,最大降水量达88 mm,春季和秋季中后期降雨较少;光合有效辐射从春季开始到夏季呈逐渐增强的趋势,随后到了秋季逐渐减弱;空气温度在2.4~44 ℃范围内变化,全年的日波动范围较大;土壤温度在8.9~31.3 ℃范围内变化,除萌芽期日波动范围较大外,其他时期的日波动范围相对较小;相对湿度与空气温度变化大致相反,在8.9%~89.9%范围内变化;饱和水汽压差与空气温度的变化大致相同,同样呈先升高后降低的趋势。
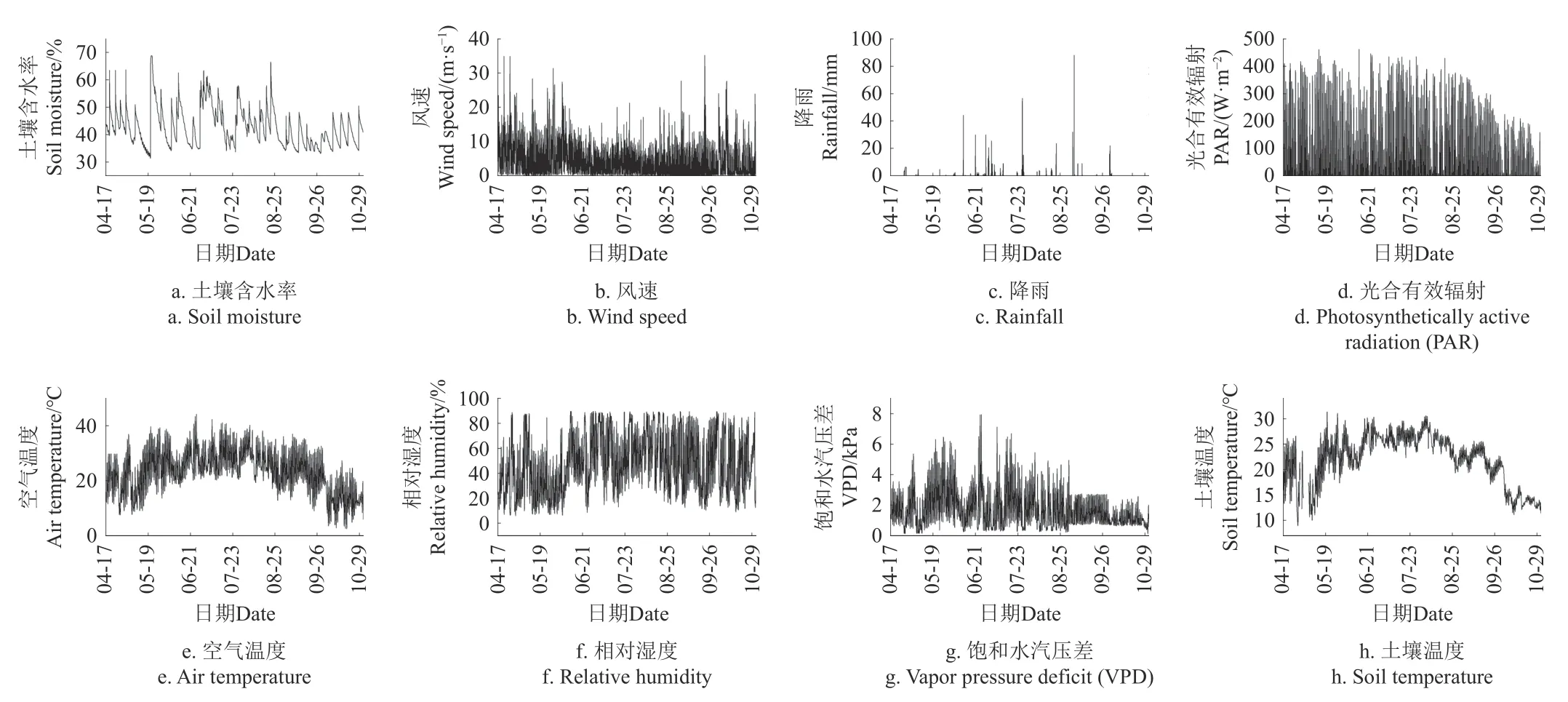
图4 生长季各环境参数变化Fig.4 Variation of environmental parameters in growing season
2.2 生长季茎干水分变化规律
因生长季各环境参数如空气温度、相对湿度、饱和水汽压差、土壤含水率等均处于持续变化之中,植物会不断调节自身生理活动及供水需求以适应环境参数带来的非线性叠加效应,导致茎干水分在生长季各时期呈不同的变化规律。如下对萌芽期、生长期及落叶期及整个生长季茎干水分变化规律进行阐述。
2.2.1 萌芽期茎干水分变化规律
图5a 为4 月18—24 日获取的一段萌芽期茎干水分变化规律,随着空气温度逐渐回升且在较充足的土壤水分条件下[36],五角枫长出少量叶片,各项生理活动如蒸腾耗散和光合固碳等逐渐恢复[37-39]。植物体耗水量即茎干组织充放水量还较少,因而茎干水分的峰值及日波动范围还较小,从凌晨开始逐渐上升,基本于13:00—14:00 达到一天中的极大值,随后开始至第二天凌晨呈逐渐下降趋势。茎干水分呈“昼升夜降”的日变化规律,这与BEEDLOW 等[8]发现的春季构树(Broussonetia papyrifera(L.) L'Hér.ex Vent.)茎干水分变化趋势较为一致。

图5 萌芽期、生长期、落叶期及整个生长季茎干水分(SW)变化Fig.5 Stem water content (SW) variation throughout the germination stage,growth stage,defoliation stage and the entire season
2.2.2 生长期茎干水分变化规律
图5b 为6 月14—20 日获取的一段生长期茎干水分变化规律,随着温度升高及在较充沛降雨条件下,五角枫生长旺盛,蒸腾耗散及光合固碳等逐渐增强[40]。茎干水分的峰值及日波动范围逐渐增大,基本从凌晨开始上升,于06:00—07:00 达到一天中的极大值;当植物体耗水量超过根部吸水量时,茎干组织会将自身储水运输至各需水组织[41-42],使得茎干水分达到极值后呈下降趋势,基本于15:00—17:00 达到极小值;随后植物体各生理活动逐渐减弱,根部吸水量超过其耗水量,茎干水分开始上升。茎干水分呈“昼降夜升”的日变化规律,与CHENG等[3-4]发现的茎干水分变化规律较为一致。
2.2.3 落叶期茎干水分变化规律
图5c 为10 月17—23 日获取的一段落叶期茎干水分数据变化规律。此时五角枫生长几近停止,叶片开始脱落,蒸腾耗散及光合固碳等生理活动的强度逐渐减弱,植物体耗水量逐渐减少,其超过根部吸水量的时刻出现滞后,即茎干水分达到极大值的时刻较前期有所后移[43],基本从凌晨开始缓慢上升,一般于12:00—13:00 达到一天中的极大值,随后至第二天凌晨呈下降趋势。茎干水分又呈现出如萌芽期时“昼升夜降”的日变化规律。
2.2.4 整个生长季茎干水分变化规律
图5d 为整个生长季茎干水分变化规律,可观察到从4 月中旬监测开始至5 月中旬,茎干水分处于较低水平,其变化缓慢且呈逐渐增加的趋势;从5 月中旬开始至8 月下旬,五角枫生长旺盛,茎干水分逐渐上升至较高水平,波动范围较前期有所增大;从8 月下旬至9 月下旬,五角枫生长变缓,茎干水分呈下降趋势且波动范围也逐渐减小;从9 月下旬开始至11 月初,五角枫出现落叶现象,茎干水分又逐渐上升至较高水平,波动范围较前期有所增加,这与HAO 等[44]发现秋季时根压重新恢复使茎干水分重新上升有较好的一致性。生长季中传感器监测到的茎干水分急剧变化是由降水导致的。
2.3 模型预测效果对比分析
分别构建BP 预测模型、GA-BP 预测模型及SSABP 预测模型利用各生长周期茎干水分数据和各环境参数组合成的数据集对茎干水分进行预测,各模型预测结果如表1 和图6 所示。

表1 各茎干水分预测模型的性能分析Table 1 Performance analysis of stem water content prediction model
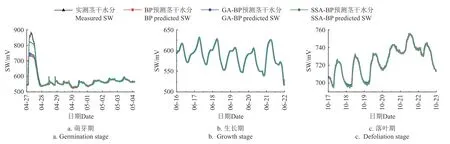
图6 生长季各时期茎干水分(SW)模型的预测值与实测值Fig.6 Predicted and measured values of stem water content (SW) at each stage of growth season
从表1 可以看出SSA-BP 模型相较于BP 模型及GABP 模型对茎干水分的预测精度高即模型评价指标R2、RMSE 和MAE 相对较低。
具体的:萌芽期,SSA-BP 模型的R2、RMSE 和MAE 分别为0.896、12.971 mV 和5.031 mV,GA-BP 模型的预测精度有所降低,其R2、RMSE 和MAE 分别为0.855、25.334 mV 和6.829 mV,而BP 模型的预测效果则较差,其R2、RMSE 和MAE 分别为0.817、27.299 mV和8.905 mV;生长期,SSA-BP 模型的R2、RMSE 和MAE 分别为0.934、2.170 mV 和0.762 mV,GA-BP 模型的R2、RMSE 和MAE 分别为0.916、3.924 mV 和1.246 mV,BP 模型的R2、RMSE 和MAE 分别为0.901、5.276 mV 和2.275 mV;到了落叶期各模型预测精度均有所提高,SSA-BP 模型的R2、RMSE 和MAE 分别为0.987、0.314 mV 和 0.232 mV,GA-BP 模型的R2、RMSE 和MAE 分别为0.973、0.636 mV 和0.439 mV,而BP 模型的R2、RMSE 和MAE 分别为0.966、0.861 mV和0.639 mV。
图6a~6c 分别为各模型对生长季萌芽期、生长期及落叶期中茎干水分变化的预测值与实测值变化趋势,可以看出各模型的预测值与实测值基本呈相同变化趋势,基本符合各时期茎干水分的日变化规律。
从图6a 可看出,相较于BP 模型和GA-BP 模型,SSA-BP 模型预测值与实测值间的拟合程度较高。由于4 月27—28 日出现降雨事件,茎干水分出现了较大的提升,而各预测模型的输入变量中不包含降雨量,因而在此期间,各模型预测效果均较差,这是导致萌芽期各预测模型的评价指标相对较大的原因(表1),其中SSABP 模型的预测值与实测值的接近程度较高。从图6b 和图6c 可看出,生长期与落叶期各模型预测效果较萌芽期均有明显提升,其中SSA-BP 模型预测值和实测值间的吻合程度较其他模型高即模型的预测曲线与实测曲线间的差异较小。
综上可知,SSA-BP 茎干水分预测模型可以实现对茎干水分较准确的预测。
3 结论
本研究结合无线传感网络技术与传感器技术搭建了可长期稳定运行的五角枫物联网生态信息监测系统实现对茎干水分数据及各环境参数远程实时获取与在线显示,其中采用自主设计基于驻波率原理的茎干水分传感器实现对五角枫茎干水分获取;结合植物生理生态学的相关知识分析了生长季各时期茎干水分变化特征,并构建了麻雀搜索算法(sparrow search algorithm,SSA)优化BP 神经网络即SSA-BP 茎干水分预测模型,获得结论主要有:
1)茎干水分在萌芽期、生长期及落叶期会呈现出不同的变化特征,具体的:在萌芽期和落叶期,茎干水分从凌晨开始缓慢上升,至中午时刻达到峰值,随后缓慢下降,呈现“昼升夜降”的日变化趋势;生长期,茎干水分从凌晨开始至早晨逐渐上升至极大值,随后迅速下降,至下午时刻达到极小值后开始上升,呈现“昼降夜升”的日变化趋势。
2)采用生物启发式算法即SSA-BP 构建茎干水分预测模型,相较于BP 模型和遗传算法优化后的BP 网格模型有着较好的预测性能表现即模型各性能指标要更优。在萌芽期,SSA-BP 模型的R2、均方根误差(root mean square error,RMSE)和平均绝对误差(mean absolute error,MAE)分别为0.896、12.971 mV 和5.031 mV;生长期,SSA-BP 模型的R2、RMSE 和MAE 分别为0.934、2.170 mV 和0.762 mV;落叶期,SSA-BP 模型的R2、RMSE 和MAE 分别为0.987、0.314 mV 和0.232 mV。
本研究通过多学科融合呈现出一种可行的茎干水分采集系统、分析及预测方法;然而不同地域相同树种的茎干水分在不同生长时期受不同环境参数的影响下又会呈现出不同的变化特征,因此,未来还需要在不同区域开展对相同树种和其他树种茎干水分的预测研究,从而为更系统地揭示茎干组织内部水分变化规律及为林地水分管理提供一定的指导与借鉴作用。
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29
现代园艺(2020年12期)2020-12-20
艺术品鉴(2020年6期)2020-08-11
农业机械学报(2020年3期)2020-04-01
种子(2019年5期)2019-07-02
现代园艺(2018年13期)2018-07-11
大众摄影(2018年6期)2018-06-19
现代农业科技(2017年13期)2017-08-09
山东农业科学(2017年2期)2017-03-15
江苏农业科学(2016年8期)2017-02-15
