
干旱胁迫对西藏半干旱河谷区典型灌木生理特性和解剖结构的影响1)
2023-11-24 06:02:24张喜年张长耀李紫帅辛福梅
东北林业大学学报 2023年12期
张喜年 张长耀 李紫帅 辛福梅
(林芝市林业和草原局,林芝,860000) (西藏农牧学院)
干旱是影响植物生存、生长及分布的重要环境因子之一,全球有三分之一以上土地位于干旱半干旱地区,其他地区在生长季节也会发生不同程度干旱。植物对干旱胁迫的响应包含形态结构、生理生化反应等极其复杂的变化,并形成了遭受遗传性制约的适应机制[1-4]。干旱胁迫会导致植物体内活性氧产生和清除的平衡遭到破坏而出现活性氧的积累,并由此引发氧化胁迫[5-6]。在此过程中,植物进化出有效的抗氧化系统以清除活性氧,如抗氧化保护酶、渗透调节物质等[7]。干旱胁迫下,植物通过主动积累渗透调节物质来降低体内的渗透势,使其在逆境下仍可获得维持正常生长所需的水分,从而提高抗逆适应性[8]。不同植物的不同保护酶及丙二醛(MDA)应对干旱胁迫及随之而来的氧化胁迫的响应可能存在一定差异,不同程度胁迫时植物体内起主导作用的保护酶也可能不同。叶片作为植物光合作用的主要器官,其形态结构的变化是植物对环境适应的基础[9]。在干旱胁迫环境中生长的植物,叶片往往会形成一系列抗旱耐旱的形态解剖结构特点,比如叶片小而厚、栅栏组织发达、主脉维管束直径大,角质层增厚、关闭气孔等,以形态变化来应对水分亏缺,提高水分利用效率以适应逆境[10-11]。
有关干旱胁迫对植物生理生态影响及抗逆适应机理的研究已广泛开展,研究大多是基于盆栽模拟水分胁迫,通过特定水分梯度或多水分梯度实现胁迫[12-13],研究特定水分梯度的干旱胁迫对植物生理的影响能更准确的反应植物对干旱的响应特征,特别是能较为准确地定位不同水分梯度时叶片的解剖结构特征。青藏高原大部分地区处于干旱半干旱区,长期在此生存的植物为适应干旱,已形成一系列的形态和生理适应机制[14]。以往对西藏代表性灌木耐旱的研究多是基于少数树种或不在典型的干旱半干旱河谷完成[13,15]。鉴于此,本文以西藏干旱半干旱河谷区的5种典型灌木为研究对象,研究苗木的抗氧化保护酶系统、细胞膜伤害程度、渗透调节物质、叶片气孔以及解剖特征在不同梯度干旱胁迫下的响应特征及适应机理,同时对各苗木的抗旱能力进行综合评价,以期深入了解不同梯度干旱胁迫下该区典型造林灌木的生理生态抗旱机理,为该区退化生态环境的恢复重建提供树种参考。
1 研究区概况
试验在西藏自治区林业厅林木科学研究院内完成,该研究院地处拉萨市堆龙德庆县柳梧乡桑达村,为典型的拉萨半干旱河谷地带。全年多晴朗天气,冬无严寒,夏无酷暑,气候温暖、干燥,年均气温7.4 ℃,干湿季明显,冬季干燥少雨,降水主要集中在湿季,仅6、7、8月降水量就占到全年降水总量的88.3%,多夜雨,夜雨率达80%左右,是西藏雨季夜雨最多的地区之一,降水量200~500 mm,干燥度1.5~10.0,干湿指数3~7,10 ℃以上积温2 177 ℃,无霜期133 d,全年日照时间3 000 h以上;该地区植被类型主要为亚高山灌丛和草甸植被以及河谷人工林群落[16]。
2 材料与方法
选取3年生、长势良好、形态特征相近的5种该地区典型乡土灌木树种为研究对象。所用土壤为拉萨河谷灌丛草原土,土壤田间持水量为19.41%,密度为1.23 g/cm3。栽植用花盆盆口直径为30 cm,高25 cm,每盆装入灌丛草原土14 kg,每个花盆栽植苗木1株。试验于2014年6月在西藏自治区林业厅林木科学研究院内进行,苗木栽植后经2个月缓苗期后,于8月10日开始干旱胁迫,试验共设4组处理,第1组充分供水(CK),土壤含水量为田间持水量的90%~95%;第2组轻度干旱(LS),田间持水量的70%~75%;第3组中度干旱(MS),田间持水量的50%~55%;第4组重度干旱(HS),田间持水量的30%~35%。每组设5株重复。试验时用保鲜膜覆盖花盆上表面以防止土壤水分蒸发,同时密封花盆底部以防止水分渗漏和花盆内土壤漏失。每天通过注射器在保鲜膜上扎孔注水以保证花盆内土壤含水量在设定范围内。
2.1 测定内容及方法
生理生化指标:干旱胁迫10 d后,选取各灌木幼苗枝条中上部健康完整叶片,装入自封袋后保存于装有冰块的保温箱内带回实验室备用。叶片分别剪碎混匀后,取出适量样品进行相关生理生化指标的测定,每种树种各指标重复3次。
叶绿素质量分数以丙酮提取后用分光光度计进行测定;可溶性糖质量分数采用蒽酮法测定;可溶性蛋白质质量分数采用考马斯亮蓝G-250法测定;丙二醛(MDA)质量摩尔浓度采用硫代巴比妥酸(TBA)法测定;POD活性采用愈创木酚显色法测定;SOD活性采用氮蓝四唑(NBT)光化还原法测定[17-18]。
叶片气孔和解剖特征:控水处理2周后,利用气孔印迹法观察叶片气孔密度。在典型晴天的10:00—11:30和15:30—17:30时间段内,选择发育程度相近、大小形态相似叶片进行试验。在叶片背面均匀涂抹无色指甲油,待干燥后压上双面胶带,之后将载玻片压在双面胶带上,撕下双面胶,盖上盖玻片,用OLYMPUS显微镜在不同视野下观察记录各树种叶片气孔密度。
叶片显微结构观察采用石蜡切片法。控水处理2周后采集各处理叶片。叶片取样后用FAA固定液固定,之后经脱水→透明→浸蜡→包埋→修块→切片→粘片→脱蜡→染色→透明→封片等一系列步骤后,用Phmians 2008 Cs Ver2.2显微镜在不同视野下观察并测量叶片厚度(LT)、栅栏组织厚度(PT)、海绵组织厚度(ST)等指标。
2.2 数据处理
用Excel 2010软件完成全部数据处理,用SPSS 10.0统计软件进行ANOVA分析,检验相应数据的差异显著性。
3 结果与分析
3.1 各树种幼苗生理特性对干旱胁迫的影响
①叶绿素质量分数。表1可知,不同水分条件下各灌木叶片的叶绿素质量分数之间存在显著差异(P<0.05)。细叶小红柳和江孜沙棘变化趋势相同,均在充分供水和轻度干旱条件下叶绿素质量分数相对较高,重度干旱下叶绿素质量分数显著小于其他处理(P<0.05)。香柏叶片叶绿素质量分数在充分供水时最小,仅为重度干旱下的36%。砂生槐在充分供水、轻度干旱、中度干旱下叶片内叶绿素质量分数间差异不显著,重度干旱与其他处理间差异显著(P<0.05)。醉鱼草在中度干旱时叶绿素质量分数达到1.95 mg/g,显著高于其他处理(P<0.05)。

表1 不同水分梯度下各树种幼苗叶绿素、可溶性糖、可溶性蛋白质质量分数的变化
充分供水条件下,江孜沙棘叶绿素质量分数最高,砂生槐和醉鱼草质量分数有所下降,与江孜沙棘差异显著(P<0.05),细叶小红柳和香柏叶绿素质量分数约为江孜沙棘的50%和25%。轻度干旱时江孜沙棘和醉鱼草叶绿素质量分数较大,砂生槐叶绿素质量分数略有下降,细叶小红柳和香柏叶绿素质量分数最小。中度干旱时醉鱼草叶绿素质量分数最大,显著大于其他树种(P<0.05),砂生槐和江孜沙棘叶绿素质量分数差异不显著,香柏叶绿素质量分数约为醉鱼草的50%,细叶小红柳质量分数最小,仅为醉鱼草的30%。重度干旱时,醉鱼草叶绿素质量分数为1.69 mg/g,显著高于其他树种(P<0.05),砂生槐略有下降,细叶小红柳叶绿素质量分数最小,仅为醉鱼草的15%。
②可溶性糖质量分数。不同土壤水分条件下各树种苗木叶片的可溶性糖质量分数不同。细叶小红柳在充分供水时,可溶性糖质量分数最低,重度干旱时达到最大值,约为充分供水时的1.5倍,轻度和中度干旱下,可溶性糖质量分数居中且二者之间无显著差异。江孜沙棘和砂生槐在充分供水、轻度干旱和中度干旱时叶片内可溶性糖质量分数之间无显著差异,在重度干旱时砂生槐下降20%,而江孜沙棘降幅达55%。醉鱼草叶片可溶性糖质量分数在充分供水、轻度干旱和中度干旱条件下无显著差异,重度干旱下增加,约为其他条件下的1.4倍。充分供水条件下,细叶小红柳、砂生槐、醉鱼草叶片可溶性糖质量分数之间无显著差异,江孜沙棘略高于其他3个树种。轻度干旱和中度干旱时,细叶小红柳和江孜沙棘之间均无显著差异,轻度干旱时砂生槐与醉鱼草之间也无显著差异,但中度干旱时砂生槐可溶性糖质量分数约为醉鱼草的88%。重度干旱下,细叶小红柳和醉鱼草叶片内可溶性糖质量分数间无显著差异,但却显著高于砂生槐和江孜沙棘,分别为1.8和2.8倍。
③可溶性蛋白质质量分数。叶片内可溶性蛋白质质量分数在各树种及不同水分条件下均存在差异。细叶小红柳在重度干旱时可溶性蛋白质质量分数达到最大值42.53 μg·g-1,显著高于其他处理(P<0.05)。江孜沙棘在充分供水、轻度干旱和中度干旱下可溶性蛋白质质量分数间差异不显著,重度干旱下显著高于其他处理约30%。砂生槐在轻度干旱和中度干旱下差异不显著,但与充分供水及重度干旱间差异显著(P<0.05)。香柏可溶性蛋白质质量分数在轻度干旱时最小,为25.07 μg/g,充分供水、中度干旱和重度干旱较轻度干旱分别增加33.90%,30.31%和38.53%。醉鱼草在充分供水、轻度干旱、中度干旱下可溶性蛋白质质量分数间无显著差异,但在重度干旱下可溶性蛋白质质量分数增加25%,与其他水分条件差异显著(P<0.05)。
充分供水、轻度干旱和中度干旱下细叶小红柳和江孜沙棘叶片的可溶性蛋白质质量分数较大,显著高于其他树种(P<0.05),轻度干旱下香柏和砂生槐叶片的可溶性蛋白质质量分数相差不大,但与醉鱼草差异显著(P<0.05),充分供水和中度干旱时香柏、砂生槐和醉鱼草叶片的可溶性蛋白质质量分数间均存在显著差异(P<0.05),由大到小表现为香柏、砂生槐、醉鱼草。重度干旱下砂生槐和醉鱼草叶片的可溶性蛋白质质量分数间差异不显著,但他们与其他3个树种之间均存在显著差异(P<0.05),其他3个树种可溶性蛋白质质量分数从大到小依次为江孜沙棘、细叶小红柳、香柏。
④丙二醛质量摩尔浓度的变化。表2可知,充分供水条件下,细叶小红柳叶片的丙二醛质量摩尔浓度最小,为13.38 mmol/g,轻度干旱时较充分供水显著提高50%,中度干旱和重度干旱下为充分供水时的近2倍。江孜沙棘在4个水分条件下丙二醛质量摩尔浓度间均差异显著(P<0.05),其中轻度干旱下最小,重度干旱下最大。轻度干旱下香柏叶片丙二醛质量摩尔浓度最低,与轻度干旱相比,对照、中度干旱、重度干旱下香柏叶片丙二醛质量摩尔浓度分别增加了52.18%,40.15%和52.27%。充分供水和轻度干旱下砂生槐丙二醛质量摩尔浓度间差异不显著,随干旱加剧,丙二醛质量摩尔浓度增加,中度干旱和重度干旱下丙二醛质量摩尔浓度约增加30%。醉鱼草在各水分梯度下丙二醛质量摩尔浓度变化趋势与细叶小红柳相同,中度干旱和重度干旱下约为充分供水时的2.4倍。

表2 不同水分梯度下各树种幼苗丙二醛摩尔质量分数、过氧化物酶和超氧化物歧化酶活性的变化
4个水分条件下,砂生槐叶片丙二醛质量摩尔浓度均显著高于其他树种(P<0.05)。水分充足时,江孜沙棘和香柏丙二醛质量摩尔浓度约为砂生槐的60%,细叶小红柳和醉鱼草丙二醛质量摩尔浓度更低,仅为砂生槐的1/2。轻度干旱下细叶小红柳和醉鱼草丙二醛质量摩尔浓度约为砂生槐的70%,但江孜沙棘和香柏叶片的丙二醛质量摩尔浓度显著低于其他树种(P<0.05),仅为砂生槐的35%。中度干旱和重度干旱下各树种丙二醛质量摩尔浓度间均差异显著(P<0.05),中度干旱和重度干旱下均为香柏叶片的丙二醛质量摩尔浓度最小。
⑤POD、SOD活性的变化。各树种叶片POD活性在不同水分条件下差异显著。细叶小红柳和江孜沙棘在4个水分梯度下活性的变化趋势相似,各水分条件下均存在显著差异(P<0.05),其中重度干旱下活性最大,其次为充分供水,轻度干旱下活性最小。香柏幼苗叶片的POD活性在轻度干旱时达到峰值4 565.90 mg·g-1·min-1,与轻度干旱相比较,充分供水、中度干旱和重度干旱胁迫时香柏幼苗叶片POD活性分别降低了48.45%,29.55%和24.51%。砂生槐在中度干旱和重度干旱下POD活性显著高于充分供水和轻度干旱(P<0.05)。醉鱼草在中度干旱和重度干旱下POD活性同样显著高于另外两个水分条件(P<0.05),轻度干旱下活性最小。
同一水分条件下砂生槐叶片POD活性均显著高于其他树种(P<0.05)。充分供水时砂生槐POD活性是细叶小红柳和醉鱼草的5.6倍,是江孜沙棘的4.5倍,是香柏的3.6倍。轻度干旱下细叶小红柳的活性最小,仅为砂生槐的6%,江孜沙棘和醉鱼草约为砂生槐的13%,香柏叶片的POD活性相对较高,约为砂生槐的52%。中度干旱下砂生槐POD活性分别是香柏、醉鱼草、江孜沙棘和细叶小红柳的3.1、5.3、6.0、10.5倍。重度干旱时,细叶小红柳和香柏叶片的POD活性间差异不显著,但其与其他3个树种之间均差异显著(P<0.05),细叶小红柳、香柏、江孜沙棘和醉鱼草活性约为砂生槐的30%、32%、25%和20%。
水分充足时,江孜沙棘SOD活性较高,其次为细叶小红柳,二者间差异显著(P<0.05),且与其他3个树种间存在差异,砂生槐和醉鱼草叶片的SOD活性差异不显著,香柏的SOD活性显著小于其他水分条件(P<0.05)。轻度干旱下砂生槐活性显著高于其他树种(P<0.05),江孜沙棘活性次之,细叶小红柳和醉鱼草活性差异不显著,香柏含量最小。中度干旱时香柏和醉鱼草叶片的SOD活性间差异不显著,与其他3个树种SOD活性之间均差异显著(P<0.05),细叶小红柳最大,江孜沙棘最小。重度干旱下细叶小红柳SOD活性显著高于其他树种(P<0.05),砂生槐活性最小,仅为细叶小红柳的70%,江孜沙棘和香柏叶片的SOD活性间无显著差异。
3.2 干旱胁迫对各树种幼苗叶片结构的影响
①气孔密度。从表3可知,细叶小红柳气孔密度在轻度干旱下最大。江孜沙棘气孔密度在充分供水、轻度干旱和中度干旱时较大,重度干旱时显著小于其余3个水分条件(P<0.05)。香柏气孔密度在轻度干旱时最大,其次为充分供水,中度干旱和重度干旱下差异不显著。砂生槐气孔密度在轻度干旱和中度干旱时显著大于充分供水和重度干旱条件(P<0.05),充分供水和重度干旱下差异不显著。醉鱼草气孔密度在充分供水和轻度干旱时较大,且二者差异不显著,中度干旱和重度干旱下差异不显著。

表3 不同水分梯度下各树种幼苗叶片气孔密度的变化
充分供水时,细叶小红柳的气孔密度最大,其次为江孜沙棘和醉鱼草,二者差异不显著,砂生槐和香柏的较小。轻度干旱时细叶小红柳的仍最大,其次为砂生槐、香柏和江孜沙棘,三者无显著差异,醉鱼草小于以上3个树种。中度干旱下5个树种中细叶小红柳、砂生槐和江孜沙棘的较大,三者差异不显著,香柏最小。重度干旱下,5个灌木树种气孔密度间差异不显著。
②解剖结构。不同水分梯度下各树种叶片解剖结构的变化如表4所示。随干旱加剧,细叶小红柳和醉鱼草叶片厚度和海绵组织厚度均逐渐减小,江孜沙棘在中度干旱时叶片厚度最大,重度干旱下最小,砂生槐在轻度干旱下叶片厚度最大,后随干旱加剧厚度减小。砂生槐栅栏组织厚度在各水分条件下均显著大于其他树种,且其在各水分梯度下差异不显著。随干旱胁迫加剧,细叶小红柳和醉鱼草叶片栅栏组织厚度递减,重度干旱下其显著小于其他水分条件(P<0.05),江孜沙棘在中度干旱时栅栏组织厚度显著高于其他水分条件(P<0.05)。随着干旱加剧,各树种海绵组织厚度逐渐减小,在重度干旱时达到最小值。香柏叶片厚度、栅栏组织厚度、海绵组织厚度、组织紧密度在4个水分条件下均无显著差异,组织疏松度随干旱胁迫加剧逐渐降低,但中度干旱和重度干旱下差异不显著。组织紧密度砂生槐远大于其余树种,醉鱼草和江孜沙棘次之,二者差异不显著,香柏最小,但与细叶小红柳差异不显著;组织疏松度香柏最大,其次为砂生槐,其余3个数种差异不显著。
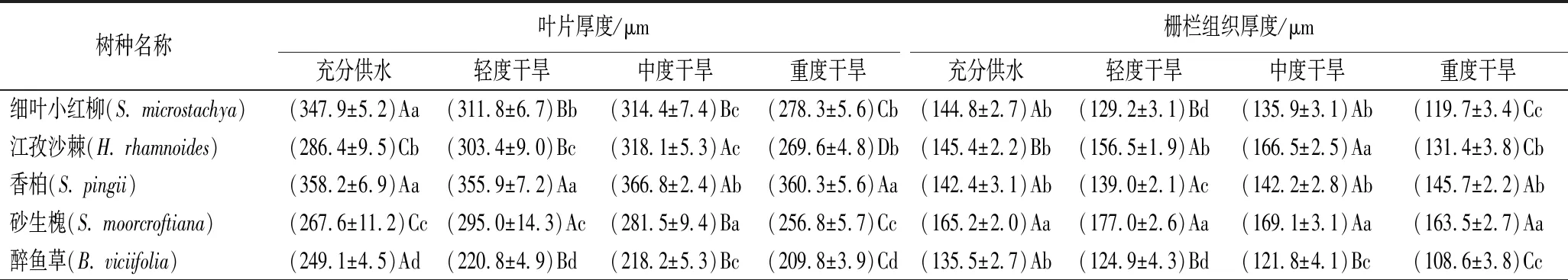
表4 不同水分梯度下各树种幼苗叶片解剖结构的变化
3.3 各树种抗旱性综合评价
干旱胁迫对灌木叶片生理及解剖影响的主成分分析结果(表5)表明,主成分1、2、3、4、5分别能够解释的方差分别占总方差的31.80%、19.97%、15.68%、11.39%、7.19%,其累计贡献率达到86.03%,也就是说,其可解释原始14个变量的86.03%。第1主成分与栅栏组织厚度及组织紧密度呈较强的正相关关系,而与海绵组织厚度及组织疏松度呈较强负相关。第2主成分与丙二醛质量摩尔浓度、POD活性、叶片厚度、海绵组织厚度呈较强正相关,而与可溶性糖、SOD呈较强负相关。第3主成分与可溶性蛋白质质量分数、叶片厚度、栅栏组织厚度呈较强正相关。第4主成分与叶片气孔密度正相关关系较强,其次为SOD活性和可溶性蛋白质质量分数,与叶绿素质量分数呈现较强的负相关关系。第5主成分与叶绿素质量分数、气孔密度呈较强正相关,而与可溶性蛋白质的负相关关系较强。

表5 干旱胁迫对各树种叶片生理及解剖影响的主成分分析结果
将各树种生理及解剖特征值与主成分的特征向量的乘积累加,并用贡献率与累加之和相乘进行加权,将同一树种的各个生理及解剖指标加权值求和,计算得出各树种在不同水分条件下的综合得分(见表6),得分越高,说明该树种的抗旱性越强[19]。砂生槐在不同水分条件下综合得分的平均值显著高于其他树种(P<0.05),达到0.997,江孜沙棘的综合得为0.462,明显低于砂生槐综合得分。而醉鱼草、细叶小红柳和香柏的综合得分均为负值,分别为-0.095、-0.29、-1.07,由此可知5个灌木树种的抗旱性从大到小的排序为:砂生槐、江孜沙棘、醉鱼草、细叶小红柳、香柏。
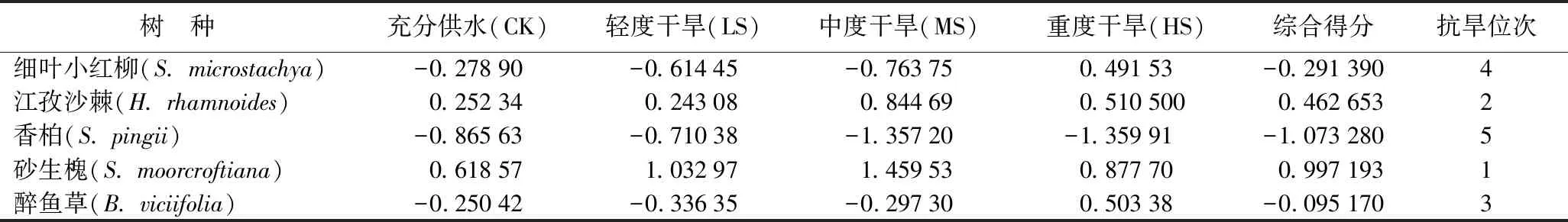
表6 5种灌木树种抗旱性综合评价结果
4 讨论与结论
随土壤含水量的减少,本研究所选择的5个树种叶片的叶绿素质量分数除香柏以外均表现为先上升后下降的趋势,说明轻度干旱胁迫下植物能维持正常的光合作用,但重度干旱时,叶绿素质量分数降低,是因为干旱会使植物原生质遭到破坏,叶绿素合成受到抑制,分解速度加快,所以质量分数下降。研究结果与其他苗木研究结果一致[13,20]。干旱胁迫下植物会通过增加渗透调节物质以增强细胞渗透调节能力。随干旱胁迫加剧,细叶小红柳和醉鱼草叶片的可溶性糖质量分数逐渐增加,在重度干旱下可溶性糖质量分数上升,可能是叶片为应对干旱,加速分解累积的糖分以提高呼吸速率所致[21]。其余树种随干旱加剧叶片可溶性糖呈现先上升后下降的趋势,说明干旱胁迫过于严重时,可溶性糖的渗透调节能力已丧失。叶片蛋白质质量分数变化随干旱加剧呈现持续增加。可见,不同树种在不同程度干旱胁迫下其可溶性糖和可溶性蛋白质质量分数变化存在差异,质量分数的高低与各树种的渗透调节能力有关,也反映了叶片渗透调节作用的有限性[22]。MDA是膜脂过氧化产物,其质量摩尔浓度的多少可以反映脂质过氧化作用的强弱和植物膜受伤害的程度[23-24]。本研究中,随干旱胁迫加剧,各树种均在重度干旱时MDA质量摩尔浓度达到最大,表明严重的干旱胁迫均造成幼苗叶片膜脂过氧化作用加强,细胞膜受到破坏。江孜沙棘和香柏在轻度干旱下MDA质量摩尔浓度有所下降,说明此水分条件比较适合各自生长。植物在正常代谢过程中,其活性氧的积累和清除处于动态平衡状态,而当植物处于逆境时,则会增加SOD和POD等保护酶活性以清除活性氧,保护细胞膜系统,使植物免受逆境伤害[25]。随干旱胁迫的变化,各树种SOD和POD活性变化不同,说明不同树种其抵抗干旱的防御系统不尽相同,单一的抗氧化酶不足以抵制植物所遭受的氧化胁迫[26],本研究中砂生槐叶片在各水分条件下均具有极高的POD酶活性,随干旱胁迫加剧,并未表现出明显的诱导反应,且其值远远的大于其余各树种,说明砂生槐能够较为稳定的保持其清除能力,受干旱胁迫的影响相对较小,这可能是其具有较强抗旱性和较高水分利用效率的原因。
叶片是植物感受干旱胁迫最敏感的器官,叶片解剖结构的变化是植物在遭受逆境胁迫后产生的适应性反应,但在干旱环境下不同植物叶片的解剖结构变化也是千差万别的。细叶小红柳的气孔密度在充分供水、轻度干旱及中度干旱下相对较大,随干旱胁迫加剧,细叶小红柳、香柏、砂生槐气孔密度呈现先上升后下降的变化趋势,而江孜沙棘和醉鱼草呈现逐渐下降的趋势,重度干旱下,除香柏以外,其余灌木树种叶片气孔密度均有不同程度的下降。可见植物叶片气孔密度不仅与植物种类有关,而且与干旱胁迫程度有关[27],一定程度的干旱胁迫会使4个乔木树种叶片气孔密度增加,但干旱胁迫程度加剧,特别是达到重度干旱时,气孔密度会减小。这与一些研究者发现的过度干旱是叶片气孔密度下降一致[28],本研究中,细叶小红柳和香柏在中度干旱下叶片气孔密度有明显下降,而江孜沙棘、砂生槐、醉鱼草在重度干旱下气孔密度下降幅度明显,而相关研究表明,植物受到严重干旱胁迫时,叶片严重受损,抑制了气孔发生,气孔数目显著减少,最终表现为气孔密度急剧下降[29],由此可见,干旱胁迫的严重程度对不同的植物而言是不同的,这在一定程度上也反映了植物的抗旱性。干旱状况下,植物叶片结构主要向降低蒸腾、增强保水和提高光合作用等方面发展[30]。叶片厚度随干旱胁迫加剧逐渐变薄的是细叶小红柳和醉鱼草。此结果与王顺才等[31]对苹果属植物叶片厚度的结果一致。也有结果表明干旱胁迫下苹果成龄叶厚度变薄,而幼叶厚度增加[32]。江孜沙棘和砂生槐叶片厚度随干旱加剧表现出增大的变化趋势,但在重度干旱下叶片厚度显著减小(P<0.05)。这是否是由于叶龄所致还有待于进一步研究。随干旱胁迫程度增加,叶片的栅栏组织厚度和海绵组织厚度表现各异,砂生槐和醉鱼草的栅栏组织厚度呈现先减小后增加的变化,而江孜沙棘和香柏则表现为先增加后减少。随干旱加剧,江孜沙棘和砂生槐海绵组织厚度先增加后减小,其余树种海绵组织厚度逐渐减小。随干旱加剧,组织疏松度各树种表现出减小趋势。由此可见,植物叶片对干旱胁迫的响应因树种差异较大,不同的植物在遭遇干旱胁迫时由于其耐旱性的差异而表现出不同的解剖特征。
各树种在不同水分条件下的渗透调节物质、抗氧化物酶活性、叶片厚度以及叶片解剖结构等表现各异,综合各树种生理及解剖特征值的主成分分析结果,砂生槐在不同水分条件下综合得分的平均值显著高于其他树种(P<0.05),达到0.997,江孜沙棘的综合得为0.462,明显低于砂生槐综合得分。而醉鱼草、细叶小红柳和香柏的综合得分均为负值,由此可知5个灌木树种中砂生槐的抗旱性最强,江孜沙棘也具有较为明显的抗旱性,醉鱼草、细叶小红柳、香柏的综合得分为负值,说明其抗旱性相对较弱。其中,砂生槐在各水分条件下的水分利用效率和POD酶活性显著高于其他树种(P<0.05),说明大幅度提高POD酶活性是其应对干旱的重要表现。
猜你喜欢
小学生学习指导(高年级)(2023年4期)2023-05-09 01:09:24
音乐天地(音乐创作版)(2020年12期)2021-01-11 01:13:42
东坡赤壁诗词(2020年3期)2020-07-04 02:50:05
中华建设(2017年3期)2017-06-08 05:49:29
西藏人文地理(2017年1期)2017-01-19 20:49:29
甘肃林业(2016年2期)2016-11-07 08:56:05
西藏科技(2016年10期)2016-09-26 09:01:35
广西林业科学(2016年3期)2016-03-16 05:43:39
岷峨诗稿(2014年3期)2014-11-15 04:13:24
华东师范大学学报(自然科学版)(2014年6期)2014-02-27 13:41:01
