
三种藤本药食同源植物光响应模型拟合及光合特性研究
2023-11-16 03:54:30肖妮洁史艳财韦霄熊忠臣邓丽丽
热带农业科学 2023年9期
肖妮洁 史艳财 韦霄 熊忠臣 邓丽丽
三种藤本药食同源植物光响应模型拟合及光合特性研究
肖妮洁1,2史艳财1韦霄1熊忠臣1邓丽丽1
(1. 广西壮族自治区中国科学院广西植物研究所 广西桂林 541006;2. 桂林理工大学旅游与风景园林学院 广西桂林 541006)
对牛尾菜、赤苍藤和土茯苓3种藤本药食同源植物的光响应曲线拟合模型与光合特性进行研究,为3种藤本药食同源植物开发利用提供依据。以牛尾菜、赤苍藤及土茯苓为试验材料,采用Li-6400光合仪测定其光合作用参数,用直角双曲线、非直角双曲线、直角双曲线修正模型和指数方程模型对3种植物的光合响应曲线进行拟合,研究3种植物光合日变化规律,并对其与环境因子的相关性进行分析。结果显示:对3种植物拟合效果最好的均是双曲线修正模型;3种藤本药食同源植物光响应曲线趋势较为一致,三者的最大净光合速率大小为牛尾菜>土茯苓>赤苍藤;3种藤本药食同源植物的净光合速率(P)、气孔导度(G)、胞间CO2浓度(C)和蒸腾速率(T)的日变化呈双峰或单峰曲线,且三者峰值出现时间略有差异,其中牛尾菜和赤苍藤的净光合速率日变化曲线都呈明显双峰形,而土茯苓的净光合速率日变化则呈单峰趋势,三者净光合速率达到第一峰值的时间相同,气孔导度日变化趋势与之相似;3种植物的净光合速率都与光合有效辐射呈正相关[表现为=0.478(牛尾菜)、=0.325(赤苍藤)、=0.060(土茯苓)],与空气温度呈负相关[相关性大小表现为赤苍藤(= –0.092)<牛尾菜(= –0.162)<土茯苓(= –0.532)],均未达显著水平;3种藤本药食同源植物利用强光能力弱,都具有一定的阴性植物特征,实际生产中可对3种藤本药食同源植物进行遮阴处理或通过控制上层植物荫蔽度来调整光照,以提高3种植物的光合效率。
牛尾菜;赤苍藤;土茯苓;光响应模型;光响应曲线;日变化
药食同源是人们在长期的生产实践中对药物和食物关系的归纳概括,从字面理解是指药物与食物均来源于自然界,具有成分和理论同源性[1]。药食虽同源但也有一定的界限,在认识药食界限同时,存在着既可以做药品也可以做食品的药食两用植物,诸多古籍医书中对这些植物都进行了详细的论述。随着国家对药食同源文化的大力推广,除已列入药食同源名录的物种外,许多在民间有着悠久使用历史的药食同源植物受到人们的重视[2]。土茯苓(Roxb.)为百合科菝葜属植物,为多年生常绿攀缘状灌木。牛尾菜(A. DC.)是百合科菝葜属多年生草质藤本植物,别名鞭鞘子菜、草菝葜、龙须菜等。赤苍藤(Blume)为铁青树科赤苍藤属多年生常绿大型木质藤本,又名腥藤、七白芍、假黄藤等。以上3种植物是我国华南地区广泛分布的3种药食同源植物,土茯苓在我国中医药应用方面有着悠久的历史,是众多常用中药复方的重要组分之一,同时也是人们常用保健食材的佳品之一[3]。牛尾菜根状茎供药用,是值得深入挖掘筛选的抗癌植物种类之一,嫩茎叶营养价值极高,是我国野菜出口的品种之一[4]。赤苍藤作为喀斯特地貌特有植物,是一种具有喀斯特地貌恢复和水土保持功能的特色经济作物,是新一代植物源痛风治疗药的重点研究对象,也是深受人们喜爱的优异森林蔬菜品种,发展潜力巨大[5]。市场对于这3种药食同源植物的需求持续增加,但目前并未形成大规模的产业化生产,间接导致了野生资源遭受破坏。目前关于这3种藤本药食同源植物的研究主要集中在化学成分、药理、生物学特性及育苗技术等方面[6-9],对于其光合生理生态方面的了解还有限,鉴于此,对牛尾菜、赤苍藤和土茯苓这3种藤本药食同源植物的光响应曲线模型与光合特性进行研究,以期为这3种植物的栽培管理提供技术支持,为其产业化生产奠定基础。
1 材料与方法
1.1 材料
供试的牛尾菜、赤苍藤及土茯苓植株种植于广西壮族自治区中国科学院广西植物研究所药食同源植物种质圃(N 25°08′29.69″,E 110°31′24.51″)。前期选取3种植物的成年植株放在遮阴度为75%的同一大棚内培养。2021年9月选取3种植物健康植株的叶片进行光合生理测试。试验材料经广西壮族自治区中国科学院广西植物研究所史艳财研究员鉴定为牛尾菜、土茯苓和赤苍藤。
1.2 方法
1.2.1 试验设计 从3种植物中分别选取植株中上部3片健康、完整的向阳叶片,采用便携式光合测定系统Li-6400(LI-COR Inc., Lincoln, USA)进行光响应曲线测定。光响应曲线的光强梯度设定为:1 600、1 400、1 200、1 000、800、600、400、200、150、100、50、20、0 μmol/(m2·s),测定前对被测叶片进行800 μmol/(m2·s)光诱导30 min。测定参数主要有净光合速率(P)、气孔导度(G)、胞间CO2浓度(C)和蒸腾速率(T)等。选取3片叶片测定结果的平均值作为光响应曲线净光合速率的实测值。用直角双曲线模型、非直角双曲线模型、双曲线修正模型和指数方程模型对光响应曲线进行拟合,各模型的具体计算公式和方法参考秦惠珍等[10]的研究。
日变化曲线时间设定为8:30、10:00、11:30、13:00、14:30、16:00、17:30。每种每个时间点取10个值,取其平均值。测定时保持叶片原有方位,记录净光合速率(P)、气孔导度(G)、胞间CO2浓度(C)和蒸腾速率(T)等光合参数以及空气温度()、空气相对湿度()、光合有效辐射()、空气CO2浓度(C)等主要环境因子的日变化。
1.2.2 数据处理 采用Origin 2018软件进行数据可视化整理,使用SPSS 26.0软件进行数据分析和检验,采用单因素方差分析和比较不同种类间各指标的差异性,数据以(平均值±标准差)表示。用光合计算软件光合计算4.1.1(版权所有:井冈山大学生命科学学院/数理学院叶子飘)对光响应曲线进行拟合。
2 结果与分析
2.1 三种药食同源植物不同拟合模型的拟合效果比较
3种植物不同拟合模型的净光合速率拟合效果如图1所示。由图1可知,4种模型对牛尾菜净光合速率的拟合效果均较好,其中非直角双曲线模型、双曲线修正模型及指数方程模型与实测值都比较接近,差异稍明显的是直角双曲线模型;赤苍藤的直角双曲线拟合值整体高于实测值,拟合效果最差,与赤苍藤实测值曲线趋势最接近的是双曲线修正模型;土茯苓的双曲线修正模型拟合曲线与实测值曲线基本重合,在光合有效辐射大于400 μmol/(m2·s)左右时,非直角双曲线模型和指数方程模型的拟合值均保持平缓不变,且曲线几乎重合,这与实测值曲线具有明显差异,而直角双曲线的拟合效果完全偏离实测值曲线趋势。说明双曲线修正模型对3种藤本药食同源植物的净光合速率拟合效果较好。

a. 牛尾菜;b. 赤苍藤;c. 土茯苓。
2.2 四种模型对3种药食同源植物净光合速率的拟合
由表1可知,光合有效辐射为0~1 400 μmol/(m2·s)时,牛尾菜净光合速率实测值随着光合有效辐射的增加而增加,而当光合有效辐射为1600 μmol/(m2·s)时,牛尾菜净光合速率下降。4种不同模型拟合效果差异不明显,且决定系数都达到0.97及以上。其中,直角双曲线模型拟合效果较其他3种模型稍差,决定系数为0.97,且最大净光合速率拟合值大于实测值;但其他3种模型的最大净光合速率均比实测值低,且只有双曲线修正模型在光合有效辐射1 400~1 600 μmol/(m2·s)时,净光合速率呈下降趋势,这与实测值趋势相同。说明牛尾菜最适的光照区间为800~1 400 μmol/(m2·s),且双曲线修正模型对牛尾菜净光合速率的拟合效果最佳。
由表2可知,光合有效辐射为0~600 μmol/(m2·s),赤苍藤净光合速率实测值随着光合有效辐射的增加而增加,在600 μmol/(m2·s)时达到最大净光合速率[2.92 μmol/(m2·s)];随后,实测值随着光合有效辐射的增加而逐渐减小。4种模型中只有双曲线修正模型与实测值趋势接近,直角双曲线和非直角双曲线均呈逐渐上升趋势,而且决定系数小,拟合效果差。指数方程拟合模型在光合有效辐射为400~1 600 μmol/(m2·s),净光合速率拟合值保持在2.50 μmol/(m2·s)不变,拟合效果不符合真实情况。因此,无论从决定系数还是净光合速率变化趋势来看,双曲线修正模型都比其他3种拟合效果更好。综上可知,赤苍藤的最适光照区间为400~600 μmol/(m2·s),且双曲线修正模型对赤苍藤净光合速率的拟合效果最佳。

表1 不同光照强度下牛尾菜净光合速率实测值及其各模型拟合值

表2 不同光照强度下赤苍藤净光合速率实测值及其各模型拟合值
由表3可知,土茯苓净光合速率实测值呈先上升后逐渐下降的趋势,在光合有效辐射为400 μmol/(m2·s)时,土茯苓具有最大净光合速率为4.93 μmol/(m2·s)。从4种拟合模型决定系数来看,除了直角双曲线型拟合效果较差(2=0.85),其他3种模型拟合效果均较理想(2=0.98或2=0.99)。从净光合速率变化趋势来看,非直角双曲线和指数方程模型的拟合值都是随着光合有效辐射的增加而增加,这与实测值趋势不符,双曲线修正模型拟合效果更接近实测值,因此初步判断双曲线修正模型拟合效果更好。说明土茯苓最适光照区间为150~800 μmol/(m2·s),且双曲线修正模型对土茯苓净光合速率的拟合效果最佳。

表3 不同光照强度下土茯苓净光合速率实测值及其各模型拟合值
2.3 三种药食同源植物拟合模型参数比较
3种药食同源植物拟合模型参数比较如表4所示。由表4可知,牛尾菜和土茯苓的真实表观量子效率均为0.4 mol/mol,赤苍藤的真实表观量子效率为0.2 mol/mol,非直角双曲线模型和指数方程的此参数的拟合值与实测值相符。牛尾菜、赤苍藤及土茯苓实测的最大净光合速率分别为7.06、2.92、4.93 μmol/(m2·s),光补偿点分别为0.41、17.92、7.07 μmol/(m2·s),光饱和点分别为162.06、140.61、136.80 μmol/(m2·s)。3种植物光响应曲线拟合效果最差的均为直角双曲线模型,该模型参数值从整体上远高于实测值;牛尾菜和赤苍藤光响应曲线拟合效果最好的均是双曲线修正模型,牛尾菜的直角双曲线模型拟合参数整体上大于实测值,而指数方程拟合参数光饱和点和光补偿点与实测值相比差异较大。从光补偿点参数来看,赤苍藤的直角双曲线模型拟合参数值远高于实测值而指数方程拟合参数稍低于实测值;土茯苓光响应曲线拟合效果较好的是指数方程,其次是双曲线修正模型,但结合2.1及2.2的分析,从模型拟合曲线及净光合速率拟合值来看,指数方程模型的拟合效果不如双曲线修正模型。因此,拟合双曲线修正模型更适用于土茯苓光响应曲线的拟合。从各参数的综合比较可以看出,双曲线修正模型对于3种药食同源植物的光响应曲线拟合效果最优。
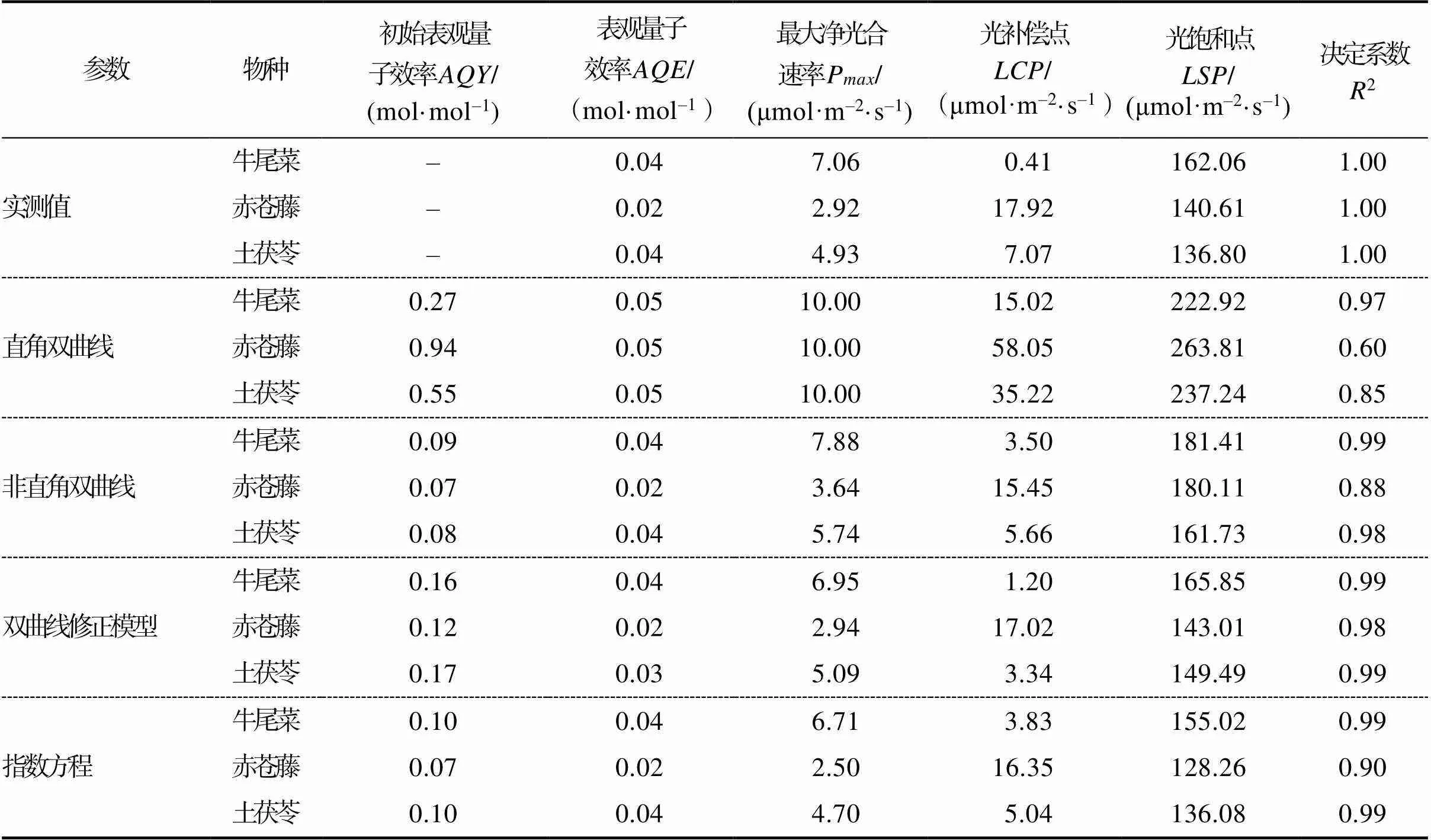
表4 三种药食同源植物拟合模型参数比较
2.4 三种药食同源植物光响应曲线及光响应特征参数
3种植物光响应曲线如图2所示。由图2可知,同一光强环境下,3种药食同源植物的光响应曲线的变化趋势一致,同时在光合有效辐射为200~400 μmol/(m2·s)出现明显的“拐点”,然后P值都随着光合有效辐射的增强而出现下降趋势,但三者的最大净光合速率存在的差异比较明显。其中,牛尾菜的最大净光合速率(P)最高,其次是土茯苓,赤苍藤的最大净光合速率(P)最低。

图2 三种药食同源植物光响应曲线
3种药食同源植物光响应特征参数如表5所示。由表5可知,虽然3种药食同源植物的最大净光合速率(P)在2.9~7.0 μmol/(m2·s),但三者存在显著差异(<0.05),最大的是牛尾菜,最小的是赤苍藤。3种药食同源植物的表观量子效率都在0.02~0.05 mol/mol。光补偿点()除赤苍藤在18 μmol/(m2·s)左右外,牛尾菜和土茯苓的光补偿点都小于10 μmol/(m2·s)。3种药食同源植物光饱和点()都偏低,牛尾菜的光饱和点约为160 μmol/(m2·s),显著高于赤苍藤和土茯苓。三者暗呼吸速率(R)都在1.25~1.34 μmol/(m2·s)。从以上参数来看,3种药食同源植物中,赤苍藤对光能的利用效率较高,光适应能力最好,光和能力最强,利于积累更多的有机物。
2.5 三种药食同源植物光合日变化
2.5.1 三种药食同源植物净光合速率(P)日变化 3种植物净光合速率日变化如图3所示。牛尾菜和赤苍藤的净光合速率(P)日变化曲线都呈“双峰形”,说明具有“光午休”现象。牛尾菜和赤苍藤净光合速率都在上午10:00时出现第一峰峰值,分别为1.91和1.41 μmol/(m2·s);牛尾菜在下午13:00时出现第二峰峰值[1.66 μmol/(m2·s)],但是此时的净光合速率小于第一峰值,而赤苍藤在下午16:00时出现第二峰值,也是最高峰值[1.58 μmol/(m2·s)]。土茯苓的净光合速率日变化呈双峰曲线趋势,在上午10:00时净光合速率达到最大值[1.62 μmol/(m2·s)];随后在11:30~14:30期间内,净光合速率明显下降,在16:00时净光合速率上升至0.64 μmol/(m2·s)后又继续下降至最小值。综合来看,土茯苓对光照的适应能力比另外2种植物更弱。
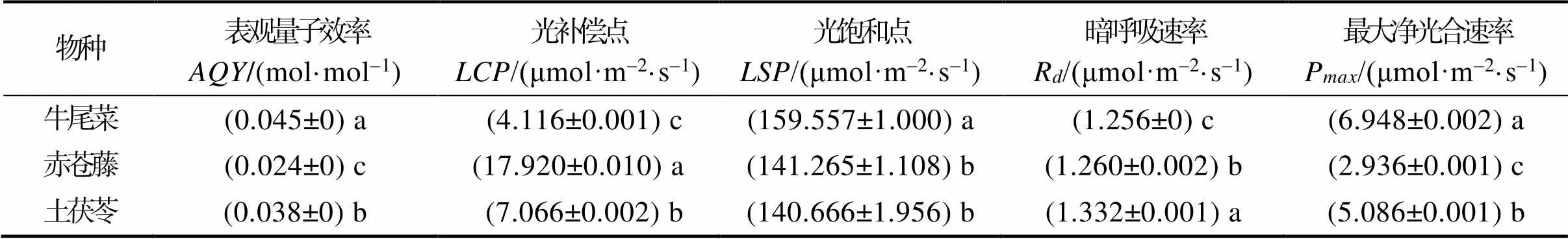
表5 三种药食同源植物光响应特征参数
注:表观量子效率;最大净光合速率;R.暗呼吸速率;光饱和点;光补偿点。不同小写字母表示不同物种间差异显著(<0.05)。

图3 三种药食同源植物净光合速率(Pn)日变化
2.5.2 三种药食同源植物气孔导度日变化曲线 3种药食同源植物气孔导度日变化曲线如图4所示。由图4可知,牛尾菜气孔导度日变化呈明显的双峰曲线,第一个峰值[G为0.033 mol/(m2·s)]出现在上午10:00时,第二个峰值[G为0.029 mol/(m2·s)]出现在下午14:30时,在17:30达到最小值[G为0.004 9 mol/(m2·s)]。赤苍藤与牛尾菜气孔导度日变化曲线趋势相似,都出现了双峰,在10:00和14:30时分别达到峰值,G分别为0.028和0.025 mol/(m2·s)。土茯苓气孔导度日变化呈单峰曲线,在上午10:00时出现最大值[0.024 mol/(m2·s)],随后气孔导度不断下降,直到14:30时开始出现平缓上升趋势,但趋势并不明显。以上表明,随着温度的升高,3种植物的气孔均受到限制,但赤苍藤和牛尾菜在光照减弱,温度降低之后,气孔仍然能够打开,从而进行光合作用,其对光的利用能力相对于土茯苓更强。
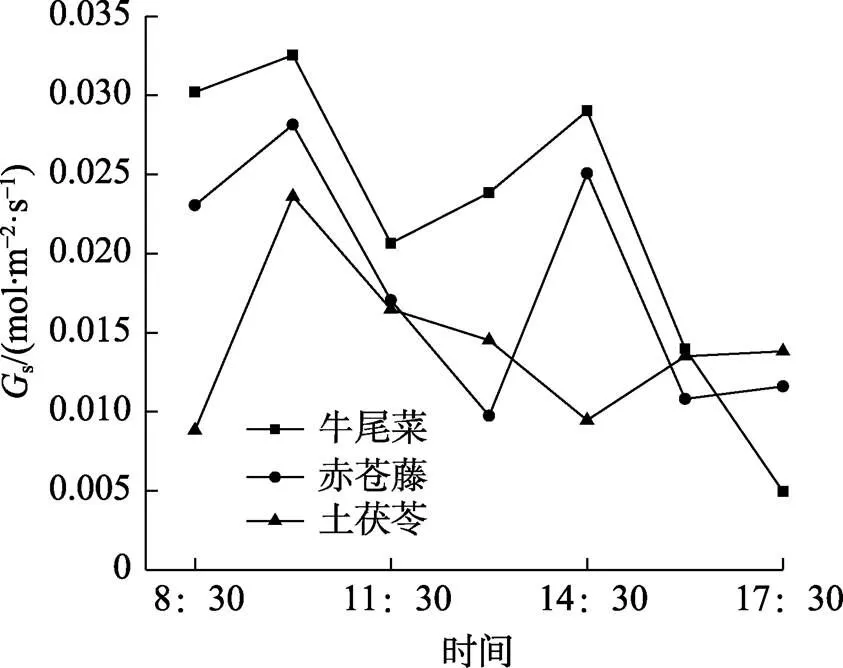
图4 三种药食同源植物气孔导度(Gs)日变化
2.5.3 三种药食同源植物胞间CO2浓度日变化曲线 3种药食同源植物胞间CO2浓度日变化曲线如图5所示。由图5可以看出,牛尾菜和赤苍藤胞间CO2浓度值都在8:30时最大,C分别为401.28和340.16 μmol/mol,牛尾菜胞间CO2浓度变化呈“倒双峰”的趋势,分别在13:00时和16:00时出现最低峰值;而赤苍藤首先在8:30至13:00时呈现下降趋势,胞间CO2浓度下降至274.48 μmol/mol,接着在14:30时增加,随后出现陡降,在16:00出现最低峰值(153.13 μmol/mol),这也许是由于植物的“光合午休”导致气孔关闭,进而导致胞间CO2浓度降低。土茯苓则呈现上升—下降—上升趋势,在11:30时达到最低值(255.40 μmol/mol)。这3种植物的胞间CO2浓度都在16:00后出现明显上升趋势,说明3种植物的气孔在16:00后开始逐渐关闭。
2.5.4 三种药食同源植物蒸腾速率日变化曲线 3种药食同源植物蒸腾速率日变化曲线如图6所示。由图6可知,牛尾菜、赤苍藤蒸腾速率日变化均呈双峰曲线,均在10:00时和14:30时出现峰值,且第二个峰值[牛尾菜的T为1.33 mmol/(m2·s),赤苍藤的T为1.18 mmol/(m2·s)]都明显高于第一个峰值[牛尾菜的T为0.83 mmol/(m2·s),赤苍藤的T为0.91 mmol/(m2·s)]。土茯苓蒸腾速率在13:00时和16:00时具有明显的峰值,分别为0.67和0.70 mmol/(m2·s)。说明高温明显提高了牛尾菜和赤苍藤的蒸腾速率,但是土茯苓的蒸腾速率反而因高温而有所降低。
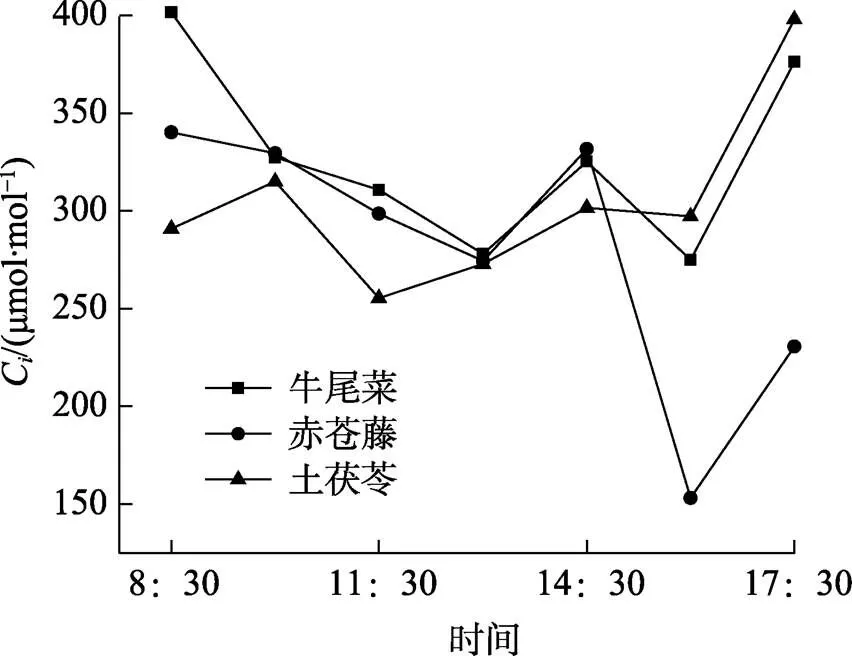
图5 三种药食同源植物胞间CO2浓度(Ci)日变化
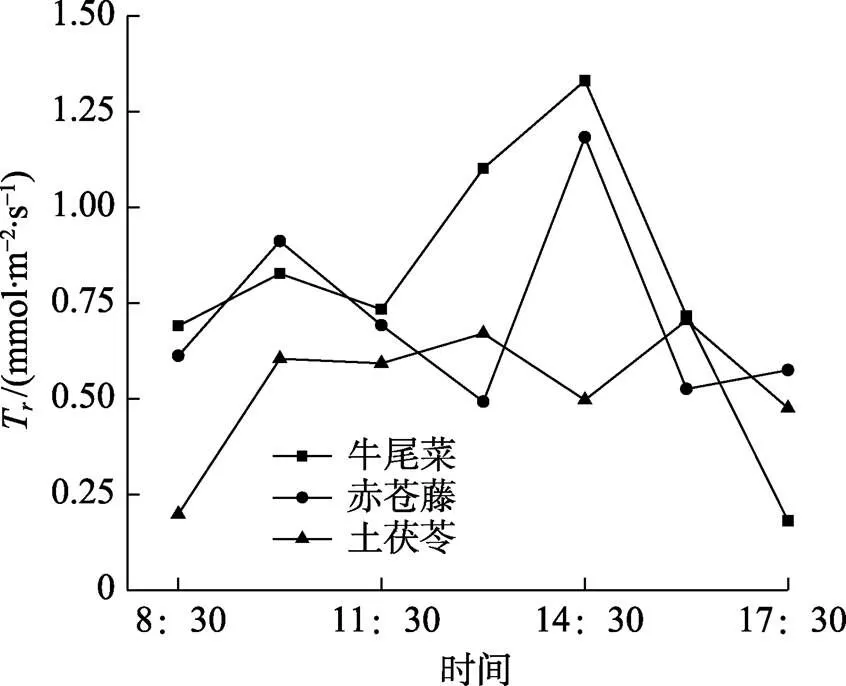
图6 三种药食同源植物蒸腾速率(Tr)日变化
2.6 三种药食同源植物净光合速率与主要环境因子的相关性
3种药食同源植物净光合速率与主要环境因子的相关性如表6所示。由表6可知,主要环境因子中空气温度与空气相对湿度、空气CO2浓度呈极显著的相关性。牛尾菜的净光合速率与空气温度、空气相对湿度及空气中CO2浓度均呈负相关但不显著(= –0.162;= –0.049;= –0.169),其净光合速率只与光合有效辐射呈正相关(= 0.478);赤苍藤的净光合速率与空气相对湿度和光合有效辐射均呈正相关,但相关性不明显(=0.103;=0.325)。土茯苓的净光合速率与空气相对湿度、光合有效辐射及空气CO2浓度均呈正相关(=0.393;=0.060;=0.493),但与空气温度呈负相关(=–0.532)。3种植物的净光合速率与主要环境因子不存在显著的相关性,说明3种植物对环境的适应能力很强,当外界环境变化时,依旧能保证光合作用的正常进行。

表6 净光合速率与主要环境因子的相关性
注:*表示差异显著;**表示差异极显著。
3 讨论与结论
直角双曲线、非直角双曲线、双曲线修正模型及指数方程4种模型是目前国内外常用的光响应模型,采用模型拟合光响应曲线并计算光合参数有利于更好地了解光合作用和不同光照对3种藤本药食同源植物生长的影响[11]。由于植物光合特性存在差异,因此选取光响应模型也有所不同。刘瑞显等[12]根据不同花生品种的光合特性选择了对应的拟合模型;徐德冰等[13]对蓝莓的光合响应曲线拟合模型进行研究发现,模型的选择存在差异性,不同拟合模型选择将直接影响蓝莓的光合特性参数。反之,模型的错误选择也将对植物的光合特性分析产生直接影响,甚至误导日后的栽培管理工作。因此根据不同植物的不同光合特性选择最适宜的模型进行拟合势在必行。本研究通过以上4种模型对牛尾菜、赤苍藤和土茯苓3种藤本药食同源植物光响应曲线进行拟合。从4种拟合模型的拟合效果图形可以看出,4种模型对牛尾菜的拟合效果均较好;与赤苍藤实测值曲线趋势最接近的是双曲线修正模型;土茯苓的双曲线修正模型拟合曲线与实测值曲线基本重合。决定系数可以表现模型拟合的效果,数值越接近1,说明模型拟合的效果越好。4种模型对牛尾菜拟合的决定系数都达到0.97及以上,效果均较好,其中,直角双曲线模型拟合效果较其他3种模型稍差;双曲线修正模型对赤苍藤和土茯苓的拟合效果比其他3种模型好。为了综合判断模型的拟合程度及适用性,不能完全根据光合曲线拟合模型的决定系数及曲线效果来判断拟合效果的好坏,更需要结合具体的模型参数进行评价[14]。本研究结合初始表观量子效率()、表观量子效率()、最大净光合速率(P)、光饱和点()、光补偿点()5种参数对拟合模型进行进一步评价。3种药食同源植物光响应曲线拟合效果最差的均为直角双曲线模型,该模型参数值从整体上远高于实测值;双曲线修正模型相比其他3种模型更适用于3种药食同源植物光响应曲线的拟合。因此,根据以上指标最终可以判断,对3种植物拟合效果最好的均是双曲线修正模型,该模型的拟合曲线符合植物生理生长规律,而且拟合模型参数都较好地反映了植物光抑制的现象。
光合作用是植物最重要的生理过程之一,是植物生长发育的物质和能量来源[15-16]。从光响应曲线来看,3种藤本药食同源植物的光响应曲线变化趋势一致,但三者的最大净光合速率大小存在显著差异:牛尾菜>土茯苓>赤苍藤。最大净光合速率(P)可以反映植物的最大光合能力[17],因此,牛尾菜的最大光合能力强于赤苍藤和土茯苓。光饱和点()是反映植物在强光条件下对光能的利用,其值越高,说明该植物更易适应强光条件。光补偿点()是指光合作用中光合产物与呼吸作用消耗相持平时的光强,也约等于净光合速率(P)为0时的光强[18]。本研究结果表明,牛尾菜的光饱和点明显高于赤苍藤和土茯苓,但与其他植物相比光饱和点明显偏低,因此3种植物利用强光能力弱。牛尾菜的光补偿点明显低于其他二者,说明牛尾菜对弱光的利用率较好,相对来说牛尾菜对光环境的适应能力较强;而赤苍藤的光饱和点低、光补偿点高,因此适宜栽植于遮阴或部分时段荫蔽的环境;土茯苓光补偿点和光饱和点均偏低,因此具有很强的耐阴性。
3种藤本药食同源植物的P、G、C、T的日变化呈双峰或单峰曲线,且三者峰值出现时间略有差异,由于叶绿素在光合作用中起着吸收光能的作用,其含量的高低直接影响到植株光合作用的强弱,这一现象可能与3种植物叶片叶绿素含量的差异有一定关联[19]。从3种藤本药食同源植物净光合速率日变化曲线可知,牛尾菜和赤苍藤的净光合速率日变化曲线都呈明显“双峰”形,说明牛尾菜和赤苍藤具有明显的午休现象,而土茯苓的净光合速率日变化则是呈单峰趋势,三者达到第一峰值的时间相同。气孔导度日变化趋势与净光合速率相似,说明气孔导度大有助于3种植物的光合作用,这一研究现象与2个杨树新品种(系)的光合特性相似[20]。净光合速率与主要环境因子的相关性研究结果表明,植物光合利用率的大小与主要环境因子有一定相关,3种药食同源植物的净光合速率都与光合有效辐射呈正相关,其中,牛尾菜(=0.478)的相关性大于赤苍藤(=0.325)和土茯苓(=0.060),说明适当增加光合有效辐射有利于3种植物的光合生产;而3种植物的净光合速率与空气温度均呈负相关且相关性都不显著,表现为赤苍藤(= –0.092)<牛尾菜(= –0.162)<土茯苓(= –0.532)。因此,与牛尾菜和赤苍藤相比,空气温度对土茯苓影响更大,也说明在一定程度上,高温不利于3种植物的光合作用。综上所述,3种藤本药食同源植物的光合特性存在一定的差异,牛尾菜相对于赤苍藤和土茯苓来说,对光的适应范围较大。因此,在栽培3种药食同源植物的过程中,可以根据3种植物的光合特性对种植地进行选择;同时,在后期管理中,可以通过适当修剪上层乔木来增加牛尾菜的光合有效辐射,使光照强度在800~1 400 μmol/(m2·s);而对赤苍藤和土茯苓可以进行适当遮阴,保持赤苍藤有效光照辐射在400~600 μmol/(m2·s),土茯苓有效光照辐射在150~800 μmol/(m2·s),以此来提高3种植物的生产效率。
[1] 谢果珍, 唐雪阳, 梁雪娟, 等. 药食同源的源流内涵及定义[J]. 中国现代中药, 2020, 22(9): 1 423-1 427+1 462.
[2] 唐雪阳, 谢果珍, 周融融, 等. 药食同源的发展与应用概况[J]. 中国现代中药, 2020, 22(9): 1 428-1 433.
[3] 于坚, 夏邦恩, 王廷玮, 等. 土茯苓的功能性分析及其在功能性食品中的应用进展[J]. 食品研究与开发, 2021, 42(8): 166-171.
[4] 董升, 李博腾, 张淑华, 等. 牛尾菜化学成分与药理作用研究进展[J]. 沈阳药科大学学报, 2021, 38(3): 304-313.
[5] 黄诗宇, 张向军, 李婷, 等. 广西新兴药食同源蔬菜赤苍藤产业发展现状与发展对策[J]. 中国瓜菜, 2021, 34(8): 109-115.
[6] 龚韦凡, 邹大江, 高瑞锡, 等. 牛尾菜根茎的化学成分研究[J]. 中药材, 2017, 40(7): 1 595-1 599.
[7] 冯颖, 张金秋, 顾地周, 等. 牛尾菜种子后熟调控[J]. 江苏农业科学, 2018, 46(2): 85-87.
[8] 张尚文, 李婷, 石前, 等. 药食同源蔬菜赤苍藤新品种桂赤苍藤1号和桂赤苍藤2号的选育[J]. 中国蔬菜, 2020(10): 92-95.
[9] 李慧, 张林. 土茯苓治疗类风湿关节炎的网络药理学研究[J]. 时珍国医国药, 2020, 31(12): 2 854-2 857.
[10] 秦惠珍, 韦霄, 唐健民, 等. 东兴金花茶和长尾毛蕊茶光合响应曲线拟合模型比较研究[J]. 江苏农业科学, 2020, 48(15): 165-170.
[11] 杨雄, 白学慧, 马关润, 等.小粒种咖啡光强和CO2响应曲线拟合模型筛选[J]. 热带农业科学, 2022, 42(2): 69-76.
[12] 刘瑞显, 王晓婧, 杨长琴, 等. 花生不同光响应曲线拟合模型的比较[J]. 花生学报, 2018, 47(4): 55-59+65.
[13] 徐德冰, 王明月, 祖蕾, 等. 适合蓝莓的光合响应曲线拟合模型研究[J]. 中国农机化学报, 2017, 38(10): 33-37.
[14] 李雪琴, 卢艺苗, 黄爱梅, 等. 亚热带10种蕨类植物的光响应模型拟合及光合特性研究[J]. 生态学报, 2022, 42(8): 3 333-3 344.
[15] 许大全. 回忆光合作用研究五十年[J]. 植物生理学报, 2016, 52(11): 1 593-1 608.
[16] 赵玉文, 段少荣, 郑雨, 等. 不同种源砂生槐幼苗的生长及光合特征[J]. 浙江大学学报(农业与生命科学版), 2019, 45(2): 164-174.
[17] 吴玲利, 熊利, 柯镔峰, 等. 3个木通品种光合特性的比较[J]. 经济林研究, 2015, 33(2): 46-49+100.
[18] 沈立明, 戴中武, 钟惠, 等. 3种独蒜兰属植物的光合特性[J]. 热带作物学报, 2021, 42(5): 1 355-1 360.
[19] 柴胜丰, 庄雪影, 韦霄, 等. 光照强度对濒危植物毛瓣金花茶光合生理特性的影响[J]. 西北植物学报, 2013, 33(3): 547-554.
[20] 王兴胜, 李斌, 陈勇, 等. 不同派系杨树品种(系)光合特性评价[J]. 南方农业学报, 2018, 49(12): 2 476-2 485.
Light Response Model Fitting and Photosynthetic Characteristics of Three Vines of Medicinal and Edible Homologous Plants
XIAO Nijie1,2SHI Yancai1WEI Xiao1XIONG Zhongchen1DENG Lili1
(1. Guangxi Institute of Botany, Chinese Academy of Sciences, Guilin, Guangxi 541006, China; 2. College of Tourism & Landscape Architecture, Guilin University of Technology, Guilin, Guangxi 541006, China)
The light response curve fitting model and photosynthetic characteristics of three vine medicinal and food plants, namely,,, and, were investigated to provide a basis for the development and utilization of these three vine medicinal and food plants. Taking,, andas experimental materials, a Li-6400 photosynthesizer was used to measure their photosynthetic parameters, and the photosynthetic response curves of the three plants were fitted with right-angle hyperbola, non-right-angle hyperbola, right-angle hyperbola modified model and exponential equation model to study the changing pattern of the photosynthetic daily rule of the three plants and analyze their correlation with the environmental factors. The results showed that the hyperbolic correction model had the best fitting effect on the three plants; the trends of the light response curves of the three rattan medicinal and food plants were relatively consistent, and the maximum net photosynthetic rates of the three plants were>>; the daily changes in,,, andof the three rattan medicinal and food plants showed a bimodal or unimodal curve, and the time of the peaks of the three plants was slightly different, among which the daily net photosynthesis rate curves ofandboth showed a clear bimodal shape, whereas the daily net photosynthesis rate ofshowed a single-peak trend, with all three net photosynthesis rates reaching the first peak at the same time, and the daily stomatal conductance trend was similar to that of. The net photosynthetic rates of all three plants were positively correlated with photosynthetically active radiation [= 0.478 (),= 0.325 (),= 0.060 ()] and negatively correlated with air temperature [the magnitude of the correlation was(= –0.092) <(= –0.162) <(= –0.532)], and none of them reached a significant level. The three rattan medicinal and food plants were weak in utilizing strong light, and all of them had certain characteristics of shade plants. In actual production, the three rattan medicinal and food plants can be shaded, or the light can be adjusted by controlling the degree of concealment of the upper plants to improve the photosynthetic efficiency of the three plant species.
;;; light response models; light response curve; daily change
Q949.95
A
10.12008/j.issn.1009-2196.2023.09.003
2023-02-08;
2023-02-23
国家林业和草原局重点研发项目(No.GLM [2021]037号);广西植物所基本业务费项目(No.桂植业21012,No.桂植业21013)。
肖妮洁(1998—),女,硕士研究生,研究方向为园林植物应用与景观设计,E-mail:1208141365@qq.com。
邓丽丽(1992—),女,硕士,助理研究员,研究方向为药用植物良种选育及栽培技术,E-mail:denglilimini@163.com。
(责任编辑 林海妹)
猜你喜欢
基层中医药(2021年5期)2021-07-31 07:58:34
基层中医药(2021年12期)2021-06-05 06:56:40
基层中医药(2020年10期)2020-11-27 01:58:58
基层中医药(2020年9期)2020-11-27 01:55:28
吉林中医药(2020年3期)2020-02-14 03:59:27
特别健康(2018年2期)2018-06-29 06:13:50
食品与健康(2018年6期)2018-06-14 02:58:08
时代青年(上半月)(2017年11期)2017-12-06 06:24:46
故事会(2017年15期)2017-08-08 19:07:34
饮食与健康·下旬刊(2016年12期)2017-01-05 16:11:43
