
云南省三江并流区混种鸟群秋季时空分布格局
2023-11-15 02:35段廷璐杨学俊沈秀英郭龙洁李迎春谭坤张淑霞王荣兴李娜肖文
四川林业科技 2023年5期
段廷璐, 杨学俊, 沈秀英, 郭龙洁, 李迎春, 谭坤,5,6,7,张淑霞,5,6,7, 王荣兴, 李娜,5,6,7*, 肖文,5,6,7
1. 大理大学东喜玛拉雅研究院, 云南 大理 671003;
2. 高黎贡山国家级自然保护区福贡管护分局, 云南 怒江 673400;
3. 高黎贡山国家级自然保护区怒江管护分局, 云南 怒江 673200;
4. 高黎贡山国家级自然保护区贡山管护分局, 云南 怒江 673500;
5. 中国三江并流区域生物多样性协同创新中心, 云南 大理 671003;
6. 大理大学三江并流区域生物多样性保护与利用云南省创新团队, 云南 大理 671003;
7. 国际生物多样性与灵长类保护中心, 云南 大理 671003
当前生物多样性丧失的速度加快[1-2],生物多样性空间格局及形成机制是宏观生态学和生物地理学研究中最为基础的科学问题之一,而物种多样性则是生物多样性的重要组成部分。海拔和纬度梯度被认为是影响生物多样性分布格局的关键因素[3-4],纬度和海拔梯度的共同作用,几乎决定了物种的空间分布。研究物种多样性的纬度梯度、海拔梯度分布格局对物种多样性的监测和合理保护措施的制定有着重要的意义。
混种鸟群是指两种及其以上的鸟类出现在一个相对集中的空间范围,并大致朝着同一个方向移动、觅食,在热带、亚热带森林中普遍存在的现象,全球有超过1 000种鸟类参与混种群[5-6]。混种鸟群是鸟类群聚中特别的社会行为表现,且存在于各种不同地理区域、海拔及栖息地类型[7-8]。混种鸟群的形成可以增加觅食效率[7]和降低被捕食风险[9-11],从而增加混群物种的生存率具有重要意义[12-13]。同时,因混种鸟群所含的鸟类物种丰富且个体数众多,比单独的个体更容易被观测、更具有可观赏性,深受科研工作者和自然爱好者的青睐。目前关于混种鸟群的研究主要涉及群落结构和生态功能[14-15]、对人为干扰的反应[16-17]和生态位的分化[18-19]等。且过去针对混种鸟群的研究大都聚焦于热带地区[15,20-22],而对温带[23-24]及亚热带[19,25-26]地区的混种鸟群研究较少。在时间梯度上,大都针对单一物种的日活动节律[27]和季节变化导致混种鸟群发生情况的不同[24,28-29]。
关于混种鸟群在较大纬度梯度和海拔梯度上的时空分布格局研究较少。在纬度梯度上,Hutto指出,随着纬度降低,混种鸟群组成结构的复杂度也会随之提高[28];Zou F. S的研究表明纬度梯度的变化对混种鸟群的影响较大,热带地区的鸟类物种丰富度均高于温带[30]。在海拔梯度上,国内关于混种鸟群的研究中,台湾相对较多,且主要集中在中、低海拔山区[31-33],仅有王颖和陈炤杰于太鲁阁中、高海拔地区进行过混群组成的初步探讨[34];大陆在西南地区,Liping Z等人研究得出在中海拔(1 400 m以上)森林中的混种鸟群多于森林与农作物种植区之间的缓冲区[26];Zhang等人在南岭进行的研究表明混群是影响群落结构的重要因素,而海拔高度对其的影响则相对较弱[25]。但同时讨论海拔和纬度两种梯度上混种鸟群的发生规律的研究较少,多数研究为海拔或纬度某一种梯度的讨论[26,29,32]。
因此,本研究通过对云南省三江并流区的鸟类进行监测,旨在阐明混种鸟群在纬度和海拔梯度上的时空分布规律。研究结果将有助于了解三江并流区混种鸟群分布的时空规律,为科学的保护三江并流区域鸟类物种多样性提供依据;同时也有助于滇西北地区生态旅游设计观鸟路线,为观鸟生态旅游规划提供参考和支撑[35]。最后,本研究可为较大环境梯度上混种鸟群形成的驱动机制提供基础。
1 区域概况
“三江并流”自然遗产地主要位于云南西北部,是印度板块与欧亚板块碰撞、缝合的地质遗迹区,毗邻西藏自治区和四川省,为青藏高原南延部分,以怒江、澜沧江、金沙江等大型河流和高黎贡山、碧罗雪山、云岭等横断山脉为主体,构成独特的纵向岭谷区,被称为三江并流区[36-37]。三江并流区是世界生物多样性最丰富的地区之一,是北半球生物景观的缩影,名列中国生物多样性保护十七个“关键地区”的第一位,是世界级物种基因库,是中国三大生态物种中心之一。这里集中了北半球南亚热带、中亚热带、北亚热带、暖温带、温带、寒温带、温带、寒带的多种气候和生物群落,是地球最直观的体温表和中国珍稀濒危动植物的避难所。因其十分丰富的生物多样性而受到广泛关注,是全球25个生物多样性保护的热点地区之一[38]。三江并流形成世界上“江水并流而不交汇”的奇特自然地理景观,其鸟类种类多,种群数量丰富,分布集中[39],为开展混种鸟群的时空分布格局研究提供了条件。
2 研究方法
2.1 样线布设
2019年4月野外踏查,了解区域整体的纬度、海拔、植被、生境分布情况及路况,选定了4个纬度梯度,分别为姚家坪片区、知子罗片区、石月亮片区(27.14°N)独龙江片区,海拔范围均为1 500~3 300 m。设置样线6条,总长度为84 km,各路线上海拔从低到高每上升200 m设置3个样点,共计155个样点(见表1)。9月13日—10月5日开展调查。
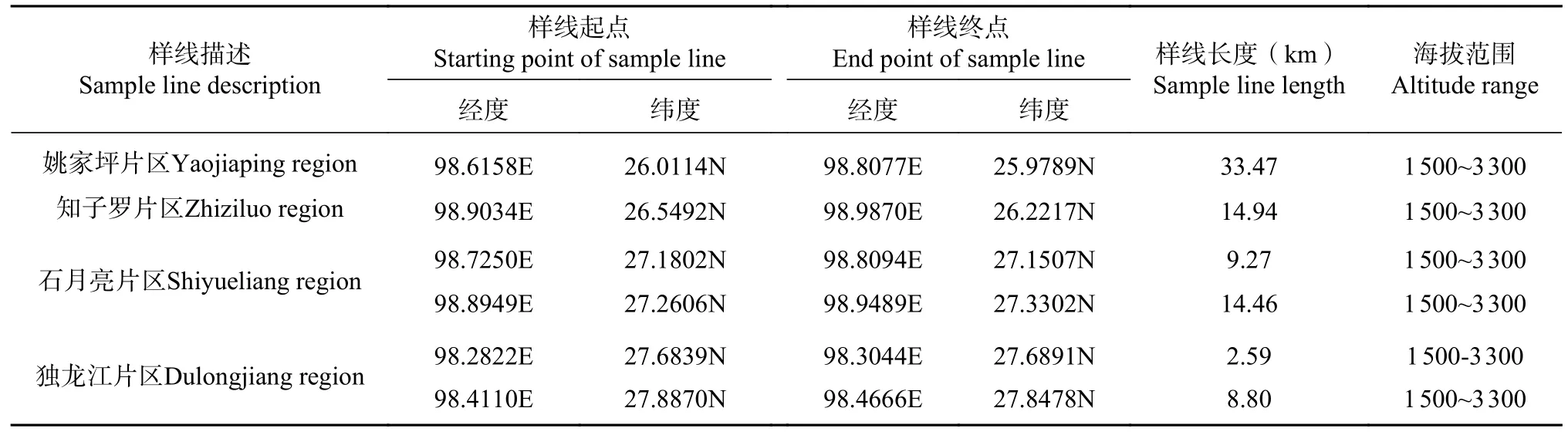
表1 秋季混种鸟群调查样线设置Tab. 1 Transect lines for the research of mixed-species bird flocks in Autumn
2.2 混种鸟群调查
调查期间使用样线和样点结合法进行调查,调查时间为每天7:00—13:00及15:00—19:30两个时间段,遇下雨和阴天天气根据实际情况调节调查时间。调查人员4~5人,持双筒望远镜、单反相机及录音笔,对观察到的所有鸟类进行识别和记录。记录的信息包括:鸟类物种、数量、所处生境类型、行为、是否参加混种鸟群、混种鸟群发生时间、混种鸟群持续时间、混种鸟群物种组成、混种鸟群发生的生境类型和植被组成。
2.3 数据分析
(1)在时间梯度上,分析混种鸟群的遇见次数和不同时间段发生频率的高低;
(2)在纬度梯度上,分析混种鸟群的遇见次数和频率,以及鸟类物种丰富度和丰度的变化规律;
(3)在海拔梯度上,分析混种鸟群遇见次数和频率,以及鸟类物种丰富度和丰度变化规律;
(4)综合分析混种鸟群纬度和海拔梯度上的变化规律,以及纬度和时间梯度上的变化规律;
(5)对混种鸟群中混群物种的食性结构进行分析。
鸟类物种鉴定参考的书目是《中国鸟类图鉴》(第一版 赵欣如等编著)、《云南鸟类志》(第一版 杨岚等编著),食性信息来自《中国动物志·鸟纲》(第一版 中国科学院动物所编著)、《云南鸟类志》(第一版 杨岚、杨晓君等编著)和《鸟类分类与识别》(第一版 鲁长虎等编著)等。
3 结果与分析
调查共记录混种鸟群83次,鸟类物种数为66种。混种鸟群中以各种柳莺类居多,如橙斑翅柳莺(Phylloscopus pulcher),西南冠纹柳莺(Phylloscopus reguloides)等,其中橙斑翅柳莺(Phylloscopus pulcher)在混种鸟群中出现22次,占混种鸟群总记录次数的26.5%。较低纬度的知子罗片区单位距离长度(每一千米)鸟类物种数和混种鸟群遇见次数均较高;中纬度的石月亮片区虽然鸟类物种数较高,但是单位距离长度鸟类物种数较低;高纬度的独龙江片区虽然鸟类物种数较低,但是单位距离长度鸟类物种数较高(见表2)。
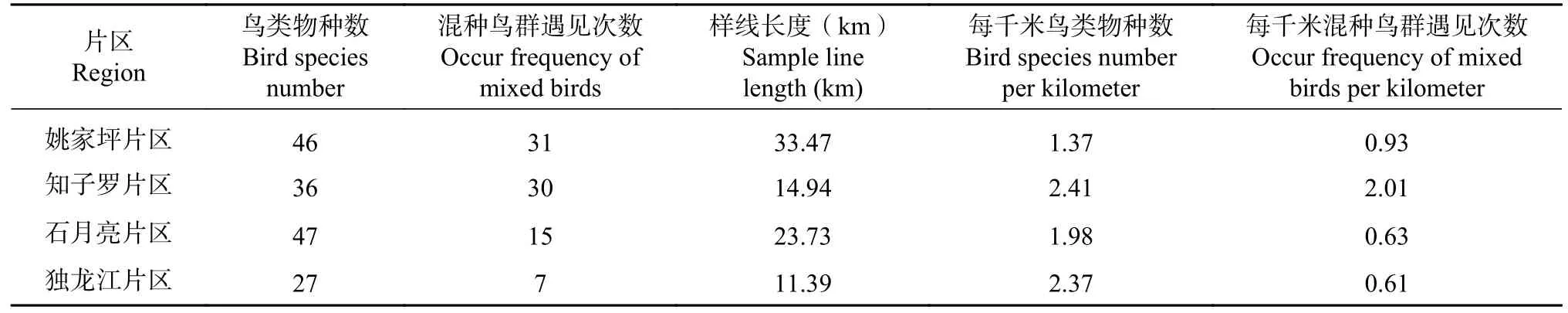
表2 鸟类物种多样性及混种鸟群调查结果Tab. 2 Mixed-species bird flocks diversity and mixed species survey results
3.1 混种鸟群在时间上的发生规律
在时间梯度上,一天内的混种鸟群发生频率呈现双峰型,即8:00—11:00以及18:00—19:00 发生率较高,尤其是8:00—9:00时段内达到顶峰(见图1)。
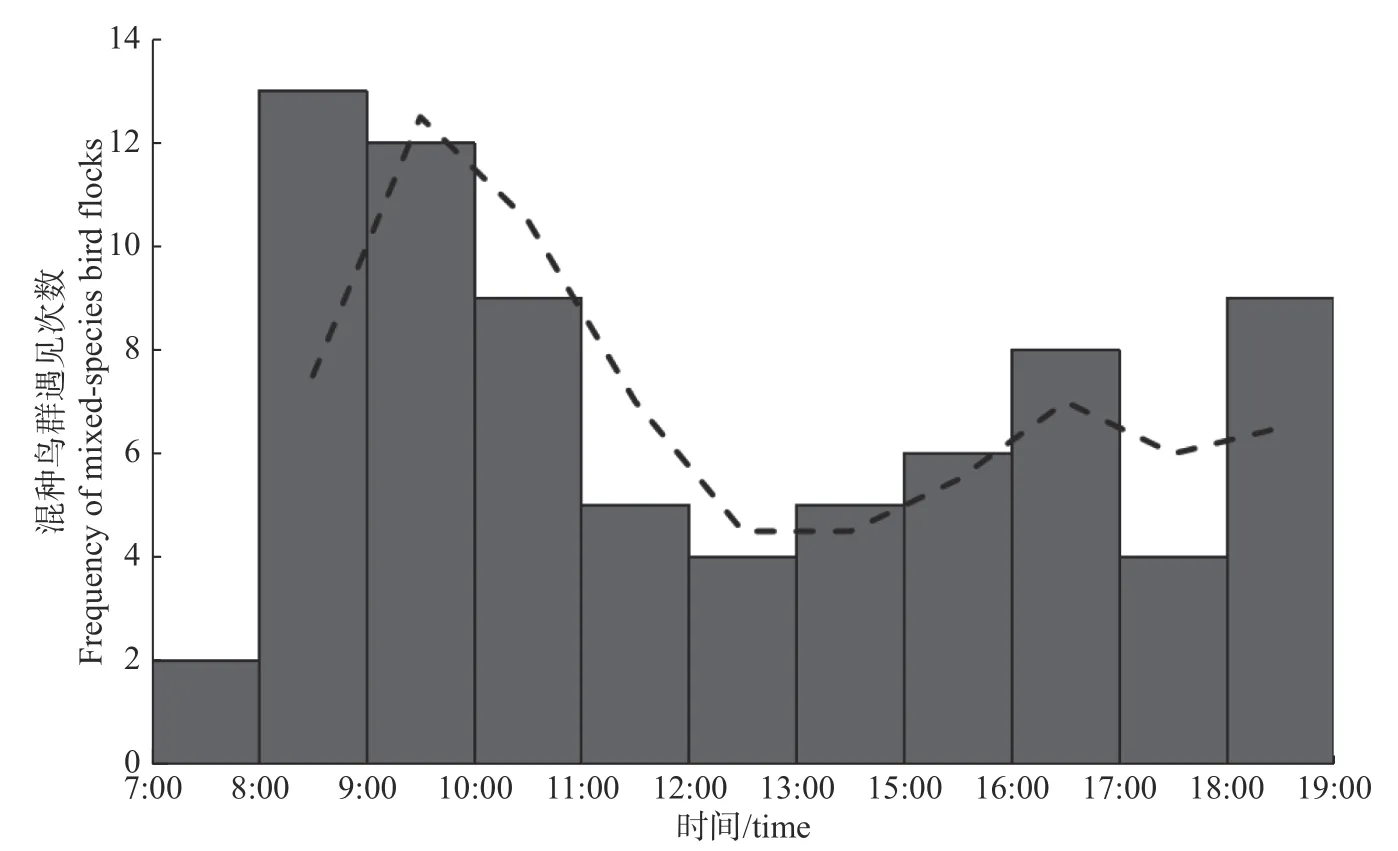
图1 混种鸟群遇见次数随时间的变化Fig. 1 Variation of occur frequency of mixed-species bird flocks within one day
3.2 调查区域内混种鸟群在纬度梯度上的发生规律
在纬度梯度上,(1)随着纬度的增加,混种鸟群的发生率降低(见图2A);同时随着纬度的增加,混种鸟群中的鸟类物种丰富度、丰度下降(见图2B和2C);(2)随着纬度的增加,混种鸟群在时间梯度上由单峰分布向双峰分布、偏峰分布和低频分布转变,且同样在早上8:00—11:00以及下午6:00—7:00 发生频率较高(见图3)。
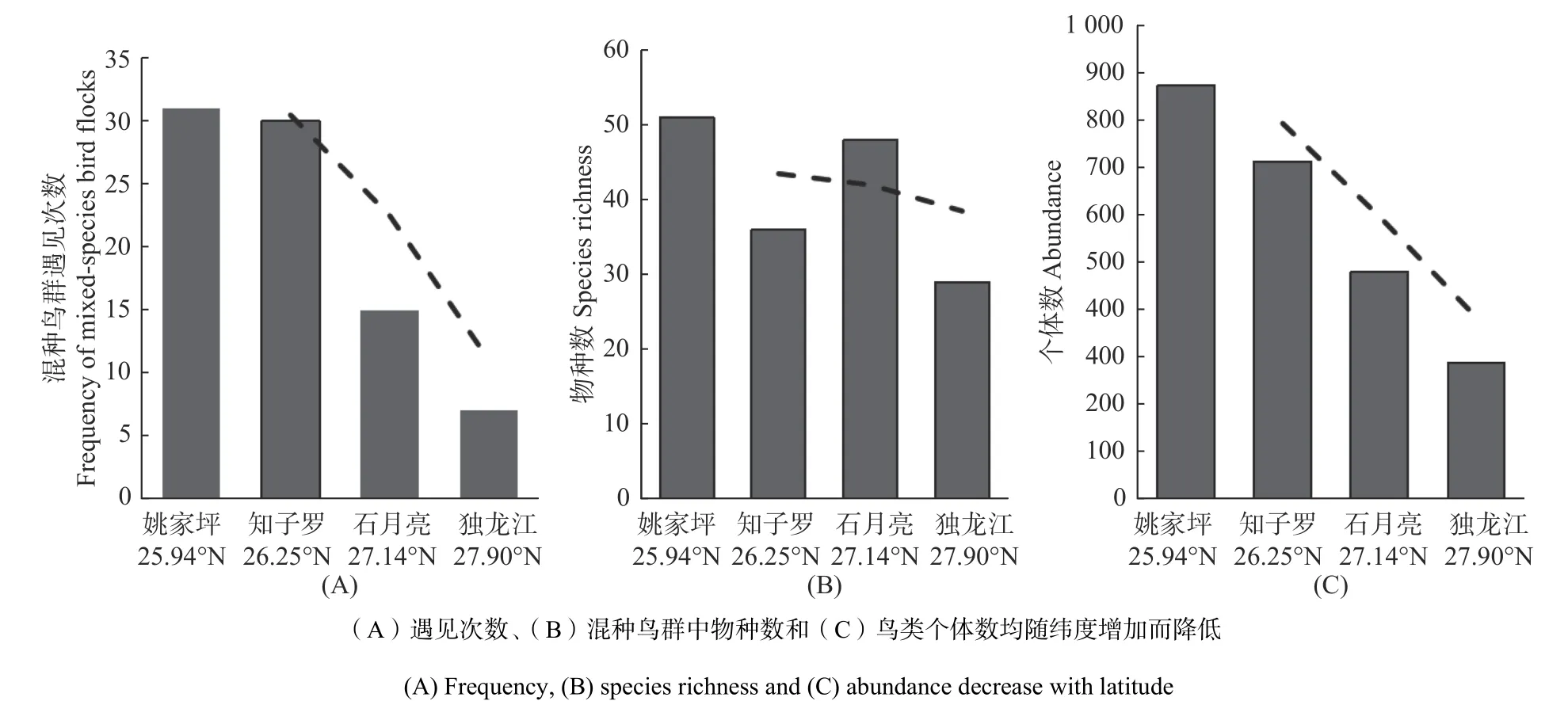
图2 混种鸟群发生情况随纬度的变化.Fig. 2 Variation of the occurrence of mixed bird populations with latitude
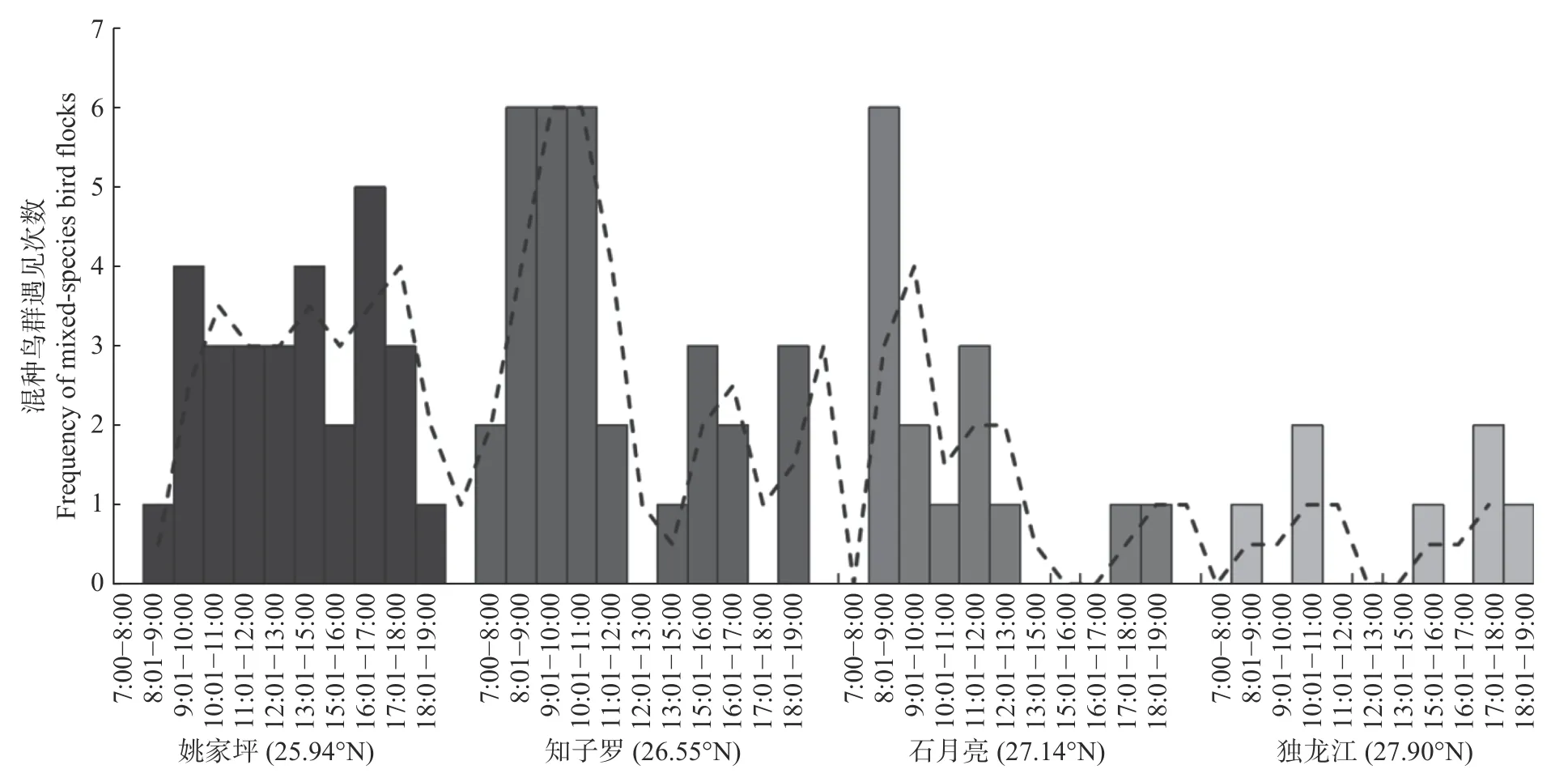
图3 混种鸟群发生的时间分布以及在纬度上的变化Fig. 3 Temporal distribution pattern and their changes of mixed-species bird flocks with latitude
3.3 混种鸟群在海拔梯度上的发生规律
在海拔梯度上,整个调查区域内混种鸟群发生率较高的范围为2 400~2 700 m,为中海拔区域(见图4),混种鸟群中的鸟类物种丰富度和丰度呈单峰分布,在中海拔上具有较高的鸟类物种丰富度(见图5)。
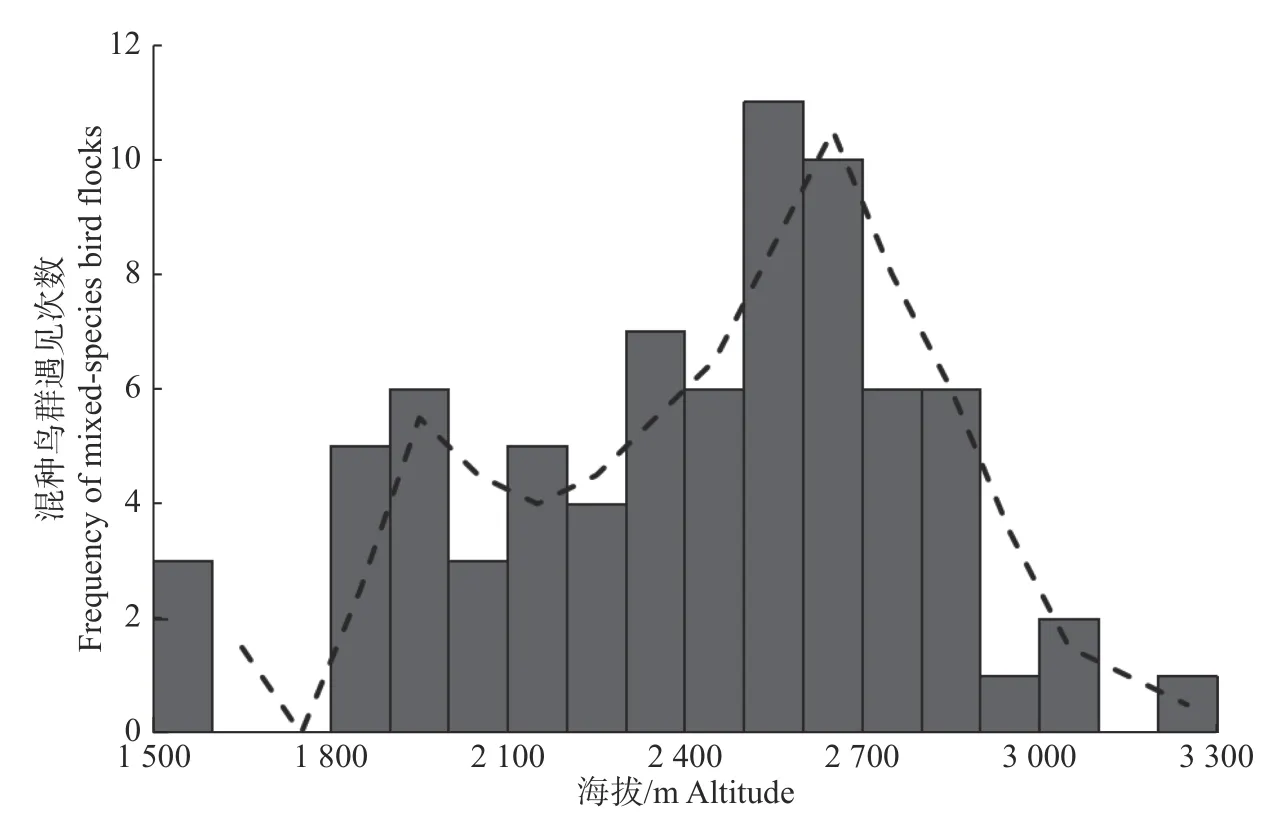
图4 混种鸟群发生情况随海拔的变化Fig. 4 Variation of the occurrence of mixed-species bird flocks with altitudinal gradient
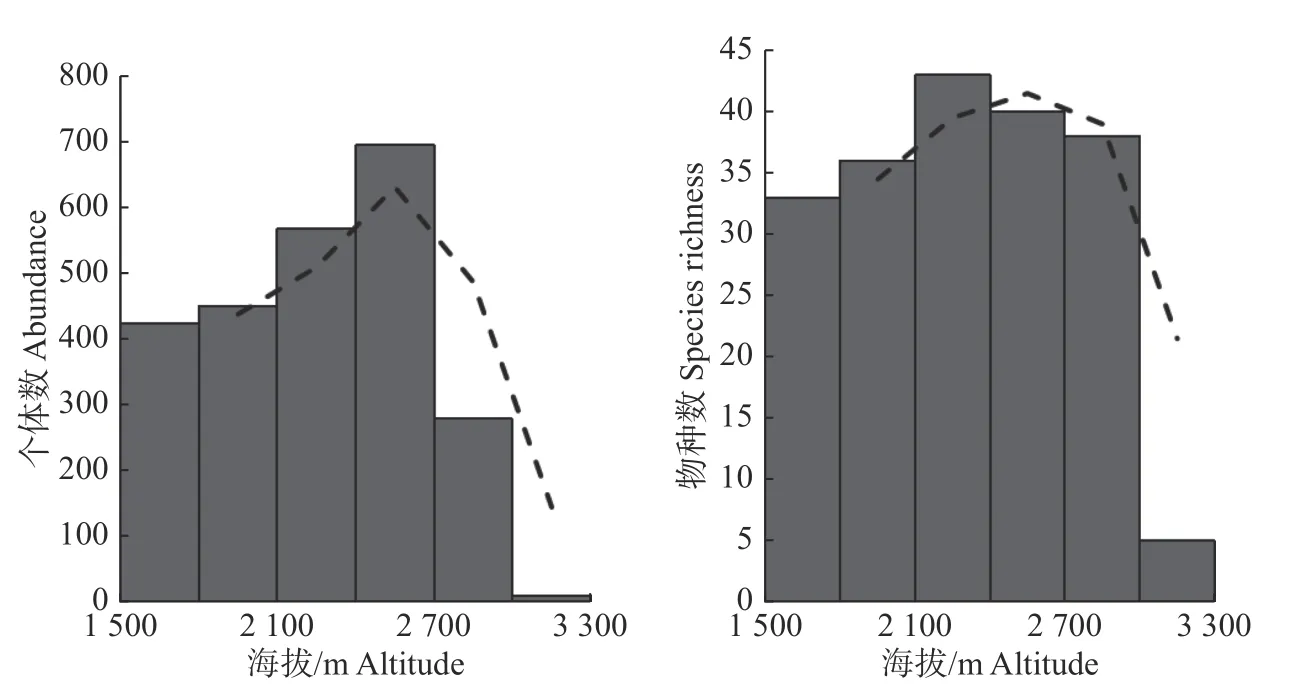
图5 混种鸟群中鸟类个体数和物种数随海拔的变化Fig. 5 Variation of the abundance and species richness of mixed-species bird flocks along altitude gradient
3.4 混种鸟群在海拔梯度上的发生情况随纬度的变化
将四个纬度地区的混种鸟群发生情况单独分析,发现混种鸟群在低纬度中海拔(2 400 m~2 700 m)地区呈现较高的发生频率,同时随着纬度的增加,整体海拔上的发生频率均降低(见图6)。
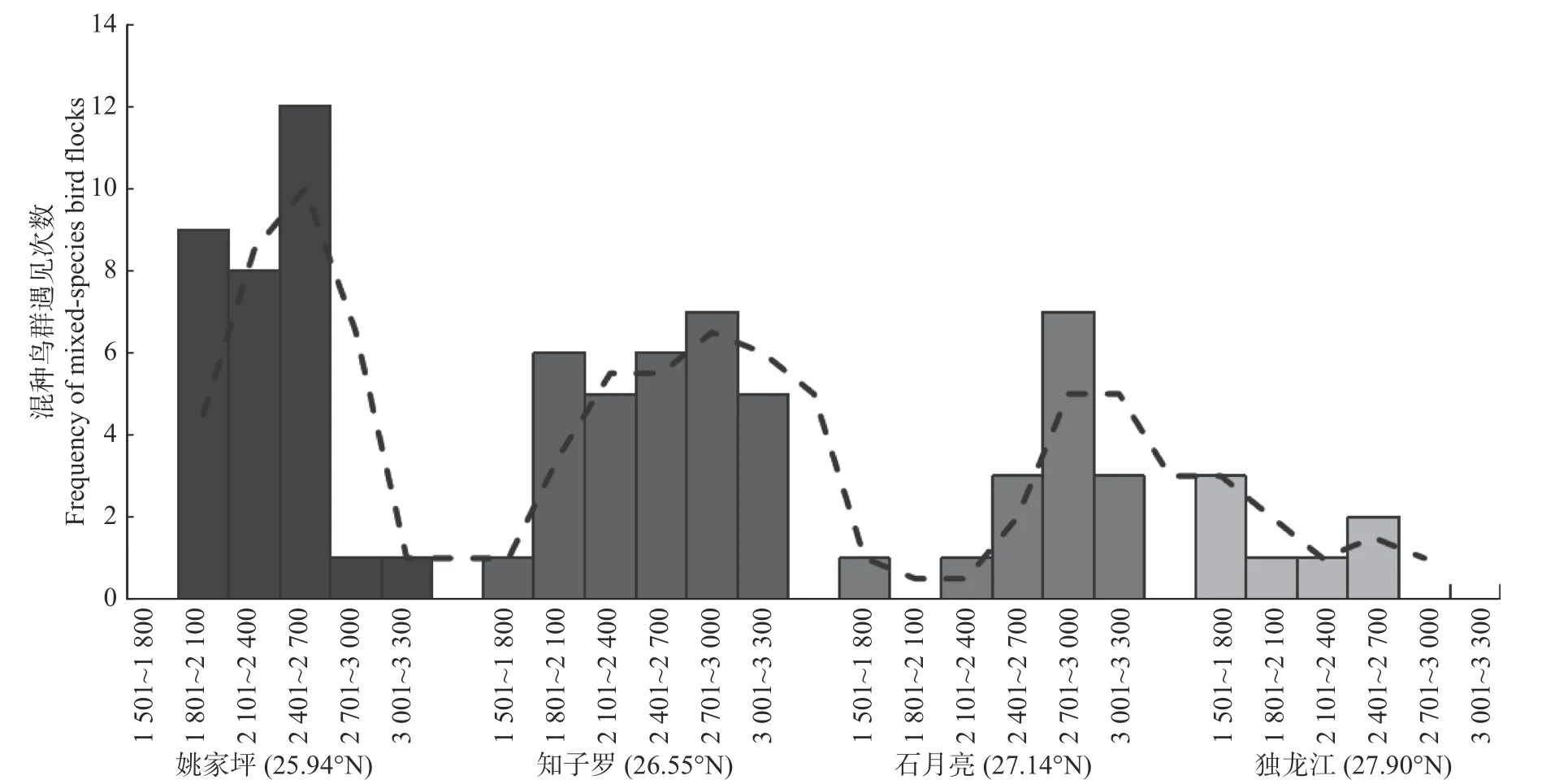
图6 混种鸟群在海拔梯度上的发生情况随纬度的变化Fig. 6 Occurrence of mixed-species bird flocks along altitude gradient
3.5 混种鸟群中鸟类物种的食性结构
三江并流区域秋季混种鸟群混群的66种鸟类中,包括食虫性(17种)和杂食性(49种)鸟类,仅喜山白眉朱雀(Carpodacus thura)为植食性,且杂食性鸟类主要以昆虫为食。
4 讨论与结论
研究探讨了在纬度和海拔梯度共同作用下,混种鸟群发生的时空规律。在时间梯度上,混种鸟群在8:00—11:00以及18:00—19:00 发生率较高,这与鸟类活动觅食的活跃时间点一致[40]。在纬度梯度上,随着纬度的增加,混种鸟群的发生率降低,同时随着纬度的增加,混种鸟群中的鸟类丰度下降,丰富度在不同地区存在差异,这可能与当地的生境差异有关[41]。在海拔梯度上,本次研究发现混种鸟群发生率较高的海拔范围为2 400~2 600 m,为中海拔区域,鸟类个体数和物种数呈单峰分布,支持高山物种多样性分布的“中间区域效应”(Mid-domain Effeet)。综合分析发现混种鸟群发生的时间随着纬度的增加由单峰分布向双峰分布、偏峰分布和低频分布转变,这可能与生境变化等因素有关;将四个纬度地区的混种鸟群发生情况单独分析,发现混种鸟群在低纬度中海拔(2 400 m~2 700 m)地区呈现较高的发生频率,随着纬度的增加在整体海拔上的发生频率降低。这与不同山区的鸟类物种数量跟海拔跨度和其所处的纬度呈显著性相关[42]。
环境的不同导致生物不同的空间分布格局,作为生物多样性热点地区之一,三江并流区域拥有复杂多样的地理特征,在海拔和纬度共同作用下,混种鸟群的时空分布格局也存在一定差异。张义荣于台湾溪南山中海拔地区的研究发现混种鸟群出现的时段并不平均,以早上7:00—9:00及下午2:00—4:00两个时段最多[43],与本研究下午混种鸟群出现的时间为18:00—19:00的结果存在差异,混群出现时段存在差异的现象可能与当地鸟类活动模式有关,在不同地理区域或栖息地间可能也会有差异。Poulsen于厄瓜多尔高海拔次生林研究混种鸟群的组成及领域关系时,发现当地混种鸟群的参与鸟种数反而比中、低海拔的混群鸟种数来的高[44],这也与本研究存在一定差异,混群的鸟种数可能会受到植层高度影响,而植层高度又会受到不同的地理差异的影响。Liping Z指出土地利用对混种鸟群物种的组成的影响比海拔对其的影响更显著[26]。因此,未来在进行混种鸟群相关研究时,除了需考虑较大空间尺度导致的差异外,不同地区生境特性也是需要注意的影响因素。
调查发现,在具有较大树龄的高大乔木、乔木下有灌丛且较开阔的生境,混种鸟群发生率较高,尤其是生境中灌木结有果实时,鸟类停留时间较长。据不完全调查,在此种生境类型中常可持续1.5小时有数十种鸟类陆续觅食活动。考虑到样线长度的影响,计算了单位样线长度(每1 km)上混种鸟群的遇见次数,结果表明知子罗和独龙江片区有较高的遇见率(见表2)。所以观鸟路线设计可以从生境类型、鸟类物种丰富度、鸟浪遇见率等方面考虑,知子罗为可选择的较好的观鸟点。虽然在不同地点考虑了海拔跨度的问题,但因为地形影响各样线设置的长度不一,在一定程度上也可能影响鸟类调查情况。
猜你喜欢
军事文摘(2023年20期)2023-10-31
散文诗世界(2022年12期)2022-08-15
小哥白尼(野生动物)(2022年3期)2022-06-16
小天使·三年级语数英综合(2019年9期)2019-11-09
小天使·三年级语数英综合(2019年4期)2019-10-06
民族音乐(2018年1期)2018-04-18
齐鲁周刊(2017年29期)2017-08-08
山东青年(2016年1期)2016-02-28
技术经济与管理研究(2014年11期)2014-03-11
测绘学报(2014年2期)2014-01-11
