
甜菜块根生长发育中呼吸代谢的特性研究
2023-11-15 12:10:22李国龙孙亚卿李宁宁张少英
作物学报 2023年12期
于 超 李国龙 孙亚卿 李宁宁 张少英
甜菜块根生长发育中呼吸代谢的特性研究
于 超 李国龙 孙亚卿 李宁宁 张少英*
内蒙古农业大学农学院, 内蒙古呼和浩特 010019
呼吸作用是作物生长发育的重要代谢基础。本研究以4个不同基因型甜菜品种为研究材料, 比较不同基因型甜菜各生育时期块根生长发育特性、呼吸速率、呼吸关键酶活性及其基因表达和能量差异, 分析各呼吸生理指标与甜菜块根增长和糖分积累的关系。研究结果表明, 不同基因型甜菜呼吸代谢水平存在显著差异, 块根呼吸速率、ATP合酶活性、ATP含量与块根重量呈极显著正相关, 表明较强的呼吸与能量代谢可促进甜菜块根的生长。联合分析呼吸代谢途径关键酶(HK、PK、IDH、SDH、6PGDH和G6PDH)活性及其基因表达和呼吸途径抑制剂效果表明, EMP-TCA是甜菜块根生长的主呼吸代谢途径, 抑制各条呼吸途径都降低甜菜的块根重量和直径, EMP、TCA和PPP途径均对甜菜块根膨大具有促进作用。研究结果为进一步深入研究甜菜块根生理机制奠定了理论基础, 为选育和鉴定高产甜菜提供了生理指标。
甜菜; 呼吸速率; 呼吸酶活性; 基因表达; ATP
呼吸作用既为植物生长提供能量, 又为植物体内有机物的形成提供原料和还原剂, 从而把植物的代谢都带动起来, 成为植物代谢的一个中心[1]。因此, 呼吸作用影响到植物的生长发育[2-3], 关系着农作物的产量和品质, 是植物新陈代谢的重要的组成部分[4-5]。
植物体内存在多条呼吸代谢途径, 不同植物、同一植物不同器官在不同环境条件、不同生育时期呼吸代谢可以有不同的途径。糖酵解(glycolysis, EMP)在植物中普遍存在, 是有氧和无氧呼吸的共同途径[6-9]。研究证实, EMP途径由己糖激酶(hexokinase, HK)、丙酮酸激酶(pyruvate kinase, PK)和磷酸果糖激酶(phosphofructokinase, PFK) 3个关键酶进行调控[10]。EMP途径的终产物在有氧条件下可进入线粒体中, 通过三羧酸循环(tricarboxylic acid cycle, TCA)途径彻底氧化分解, 该途径是脂肪、蛋白质和糖类三大类物质的共同氧化途径, 也是植物体生命活动所需能量的最主要来源[11-15], 调控TCA途径的关键酶也有3种, 分别是琥珀酸脱氢酶(succinate dehydrogenase, SDH)、异柠檬酸脱氢酶(isocitric dehydrogenase, IDH)和α-酮戊二酸脱氢酶(α-oxoxglutarate dehydrogenase, OGDC)[16-19]。戊糖磷酸途径(pentoso phosphate pathway, PPP)是植物另一呼吸代谢途径, 与植物生长发育和各种环境胁迫存在密切关系[20-22], 调控该途径的2个限速酶是6-磷酸葡萄糖酸脱氢酶(6-phosphogluconate dehydrogenase, 6PGDH)和葡萄糖-6-磷酸脱氢酶(glucose-6- phosphate dehydrogenase, G6PDH), 当EMP-TCA途径受阻时, 可启动PPP途径来代替正常的有氧呼吸, 维持作物的生长发育及对环境的适应。
甜菜(L.)是我国重要的糖料作物之一, 以收获块根榨取糖分为生产目的, 主要种植于我国北方地区。甜菜块根生长发育是其增加重量和积累糖分的过程, 因此, 研究甜菜块根生长发育机理对提高甜菜产量和含糖具有重要理论意义。对甜菜块根发育的生理和分子基础已有一些研究[23], 而有关呼吸代谢对甜菜块根发育的作用机理研究鲜见报道。本研究以高糖和丰产2种类型甜菜品种为研究材料, 通过比较不同基因型甜菜各生育时期块根呼吸速率、发育特性、呼吸关键酶活性及其基因表达和能量代谢等生理代谢差异及其与甜菜块根生长发育的关系, 阐明块根膨大的主要呼吸途径和呼吸代谢生理基础, 为甜菜高产栽培调控和遗传改良提供理论依据。
1 材料与方法
1.1 试验材料
大田试验选择块根重量和含糖不同的丰产型和高糖型品种(系)各2个, 其中丰产型品种: ST13092、SD13829 (由德国斯特儒博公司提供), 高糖型品种: TY0603、04BS02 (由内蒙古农业大学甜菜生理研究所提供)。
大田试验在内蒙古农业大学教学农场(40º51¢53¢¢N, 111º42¢52¢¢E)进行, 各品种小区面积30 m2, 3次重复, 田间管理条件一致。
为进一步检验甜菜块根发育的主呼吸代谢途径,开展了呼吸抑制剂处理盆栽试验。试验材料选用了丰产和含糖性状较突出的丰产型品种SD13829和高糖型品种04BS02。
将甜菜播种于蛭石和营养土(体积比为1∶3)的苗盘中, 在空气湿度40%、昼夜温度22℃/16℃的人工气候室内培养至10片叶进行处理。
1.2 试验方法
1.2.1 取样时期与方法 在甜菜4个生育时期各取样1次, 生育时期划分参照邵金旺《甜菜生理学》[24]。分别是苗期(Seedling stage, SS) 6月16日、叶丛快速生长期(Rapid foliage growth stage, RFGS) 7月16日、块根及糖分增长期(Root and sugar growth stage, RSGS) 8月16日和糖分积累期(Sugar accumulation stage, SAS) 9月22日取样。
取样方法: 选取长势一致的甜菜块根5株, 在块根第一个叶痕处一刀切后, 称量根重。
1.2.2 含糖率测定 用锤度计Refractometer PAL-1测定甜菜块根锤度值, 含糖率(%)=锤度值×0.80[25]。
1.2.3 块根呼吸速率测定 采用红外线气体分析仪进行呼吸速率测定[26]。
计算公式如下: 呼吸速率 =/×。
式中,为测定时CO2释放量(mg)减去初始测定CO2释放量(mg),为甜菜块根鲜重(g),为测定时间(h)。
1.2.4 酶活性的测定 本试验采用酶联免疫(enzyme linked immunosorbent assay, ELISA)试剂盒测定三磷酸腺苷合成酶(adenosine triphosphate syntherase, ATPase)、PK、HK、IDH、SDH、6PGDH、G6PDH的酶活性和三磷酸腺苷(adenosine triphosphate, ATP)含量。
1.2.5 呼吸抑制剂处理 对长势一致的10片叶龄甜菜分别喷施500 mg L–1正磷酸钠, 125 mg L–1丙二酸, 0.5 mg L–1碘乙酸和水各200 mL, 各处理分别喷施30株。处理16 h后选取5株测定呼吸速率, 处理15 d后测定甜菜块根形态指标, 3次重复。
1.2.6 呼吸代谢途径关键酶基因表达分析 利用TransZol Plant试剂盒提取各生育时期甜菜块根总RNA, 并使用First-Strand cDNA Synthesis Super Mix试剂盒反转录为cDNA。用Premier 5.0设计引物, DNAman检测, 以甜菜基因为内参基因(表1), 由TaKaRa合成引物。利用罗氏LightCycler480 II实时荧光定量PCR进行荧光定量测定。
1.3 数据处理
采用SAS 9.0进行方差分析, GraphPad进行作图, SPSS V19.0进行相关性分析。

表1 引物设计
2 结果与分析
2.1 不同基因型甜菜各生育时期块根重量和含糖率的差异
田间试验结果表明, 供试4个基因型甜菜的块根鲜重都随生育进程而逐渐增加(图1和图2-A)。苗期不同基因型品种间块根重量无显著差异, 后3个生育时期2个丰产型品种SD13829和ST13092块根重量均显著高于2个高糖型品种04BS02和TY0603。
由图2-B看出, 不同基因型甜菜的块根含糖率均随生育进程而提高, 到糖分积累期达到最高。在甜菜整个生育期高糖型品种的含糖率均高于丰产型品种, 在块根及糖分增长期和糖分积累期丰产型品种与高糖型品种含糖率差异达到显著水平(≤0.05)。
2.2 不同基因型甜菜各生育时期块根呼吸速率的差异
由图3看出, 不同基因型甜菜块根呼吸速率均表现为生育前期较低, 中期较高, 后期又降低的变化规律。不同基因型甜菜块根呼吸速率大小依次为SD13829>ST13092>TY0603>04BS02, 在叶丛快速生长期和块根及糖分增长期, 丰产型品种块根呼吸速率均显著高于高糖型品种(≤0.05), 其中在块根及糖分增长最快的时期2类品种间呼吸速率差异达极显著水平(≤0.01)。
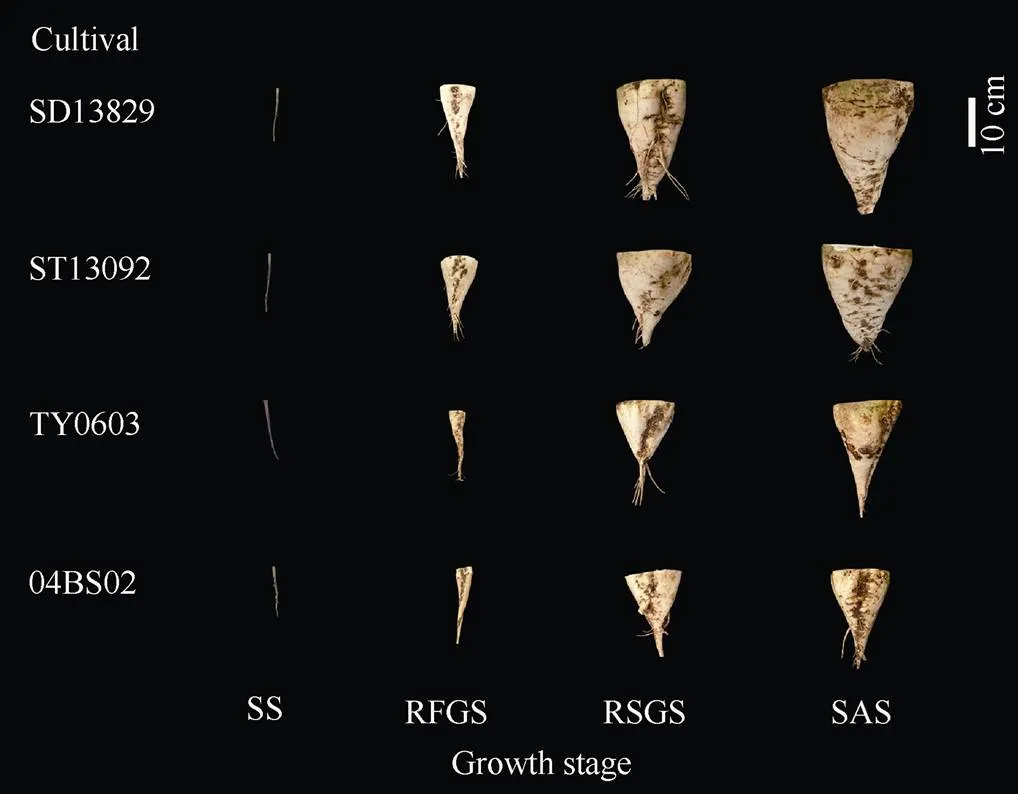
图1 不同基因型甜菜各生育时期块根形态的差异
SS、RFGS、RSGS和SAS分别代表苗期、叶丛快速生长期、块根及糖分增长期和糖分积累期。
SS, RFGS, RSGS, and SAS indicate seedling stage, rapid foliage growth stage, root and sugar growth stage, and sugar accumulation stage, respectively.
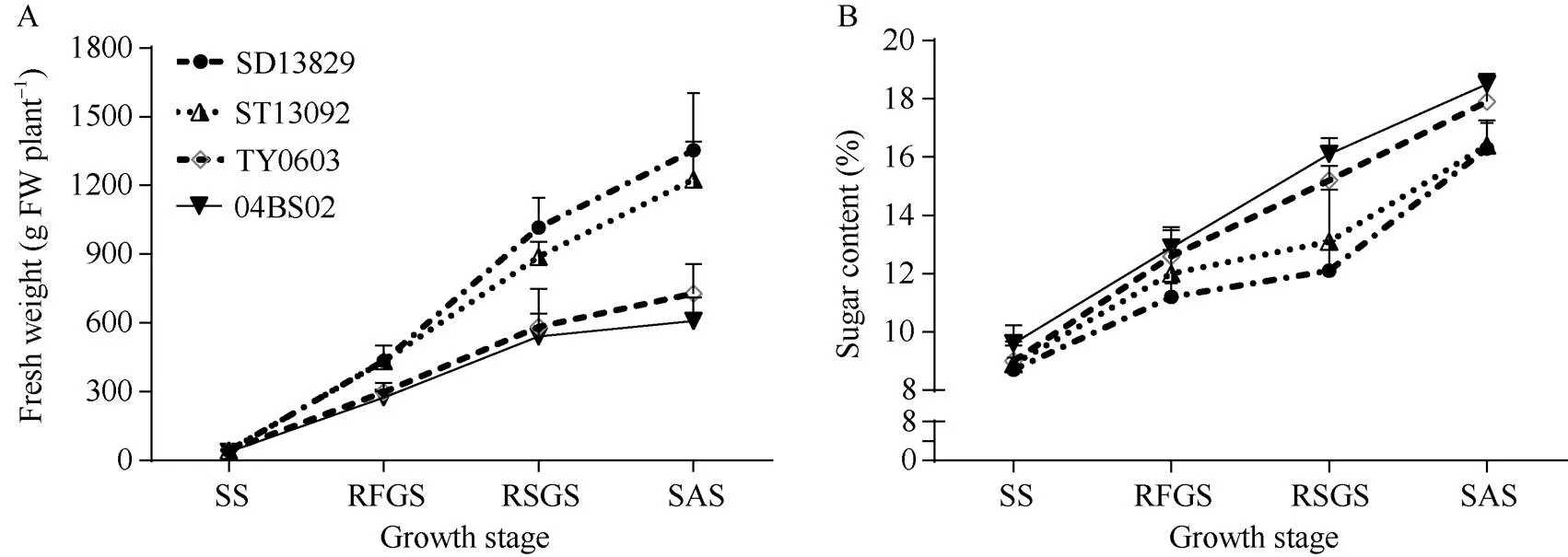
图2 不同基因型甜菜各生育时期块根鲜重和含糖率的差异
不同生育时期的缩写同图1。
Abbreviations for the different growth stages are the same as those given in Fig. 1.
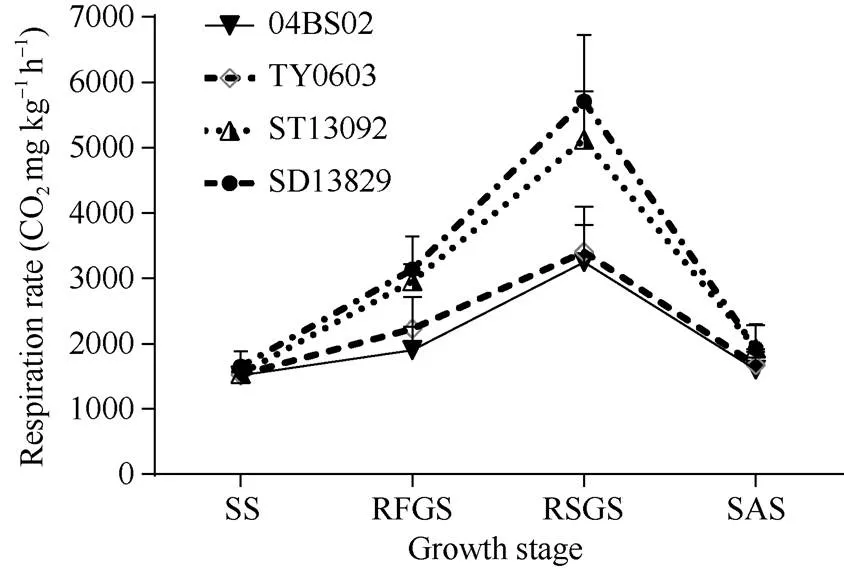
图3 不同基因型甜菜各生育时期块根呼吸速率的差异
不同生育时期的缩写同图1。
Abbreviations for the different growth stages are the same as those given in Fig. 1.
由表2中可以看出, 甜菜苗期块根根重、含糖与呼吸速率呈正相关, 但差异不显著。在甜菜生育期的后3个时期块根根重与呼吸速率成极显著正相关, 表明呼吸速率是影响甜菜块根生长的重要生理代谢; 而含糖率与呼吸速率呈负相关, 表明旺盛的呼吸代谢在增加根重的同时, 相对降低了根中糖分的积累。
2.3 甜菜块根生长过程呼吸代谢途径分析
2.3.1 不同基因型甜菜各生育时期呼吸代谢途径关键酶活性差异 从EMP、TCA、PPP 3条呼吸代谢途径中各选取2个关键酶, 对其在不同基因型甜菜生育过程块根中的酶活性分析。从图4可以看出, 甜菜各发育时期块根中EMP途径中的PK、TCA中的IDH和PPP中的G6PDH活性高于其他酶活性; 在叶丛快速生长期丰产型品种块根中IDH的活性显著高于高糖型品种, 而PK和G6PDH活性在高糖型品种中显著高于丰产型品种, 在块根及糖分增长期和糖分积累期各呼吸关键酶活性均是丰产型品种高于高糖型品种, 除丰产型品种的IDH活性有所增强外, 其他酶活性较上一生育时期有所下降。以上结果分析看, 无论什么品种, 3条呼吸代谢途径均参与了甜菜块根生长过程, 整个生育期呼吸代谢途径以EMP-TCA为主, 但高糖型品种在叶丛快速生长期有较高比例的PPP途径参与呼吸, 糖分积累期丰产型品种的TCA代谢增强。

表2 甜菜呼吸速率与块根重量和含糖率的相关系数
*、**分别表示在0.05和0.01概率水平显著相关。
*and**mean significant correlation at the 0.05 and 0.01 probability levels, respectively. SS, RFGS, RSGS, and SAS indicate seedling stage, rapid foliage growth stage, root and sugar growth stage, and sugar accumulation stage, respectively.

图4 不同基因型甜菜各生育时期块根中呼吸代谢途径关键酶活性差异
误差线上不同小写字母表示在0.05概率水平的差异显著。不同生育时期的缩写同图1。
Bars superscripted by different lowercase letters indicate significantly different at the 0.05 probability level. Abbreviations for the different growth stages are the same as those given in Fig. 1.
进一步对呼吸代谢关键酶活性与甜菜根重和含糖率的相关性分析(表3)表明, 甜菜块根生长发育过程TCA关键酶IDH、SDH活性与根重呈极显著或显著正相关, 与含糖率呈负相关, 生育中后期PK活性也是与根重呈极显著正相关, 与含糖率呈极显著负相关, 说明EMP-TCA呼吸代谢旺盛, 有利于块根生长。
2.3.2 不同基因型甜菜各生育时期呼吸代谢途径关键酶基因表达量差异 由图5可知, 不同基因型甜菜块根中各生育时期TCA关键酶、表达量最高, 且丰产型品种显著高于高糖型品种, 与酶活性结果相一致。
2.3.3 呼吸代谢途径抑制剂处理对甜菜块根呼吸速率及生长的影响 碘乙酸通过对3-磷酸甘油醛脱氢酶的不可逆抑制作用抑制EMP的进行; 丙二酸作为琥珀酸脱氢酶的竞争性抑制剂, 可阻断TCA的进行; 正磷酸钠为葡萄糖-6-磷酸脱氢酶的非竞争性抑制剂抑制戊糖磷酸途径[27]。
用3种呼吸抑制剂喷施甜菜, 由图6-A可以看出, 块根呼吸速率各处理显著低于对照, 2个品种的呼吸抑制剂对呼吸速率的影响均呈现: 丙二酸>碘乙酸>正磷酸钠, 因此, 再次证明不论在丰产品种中还是在高糖品种中, TCA是甜菜块根呼吸作用的主呼吸途径。
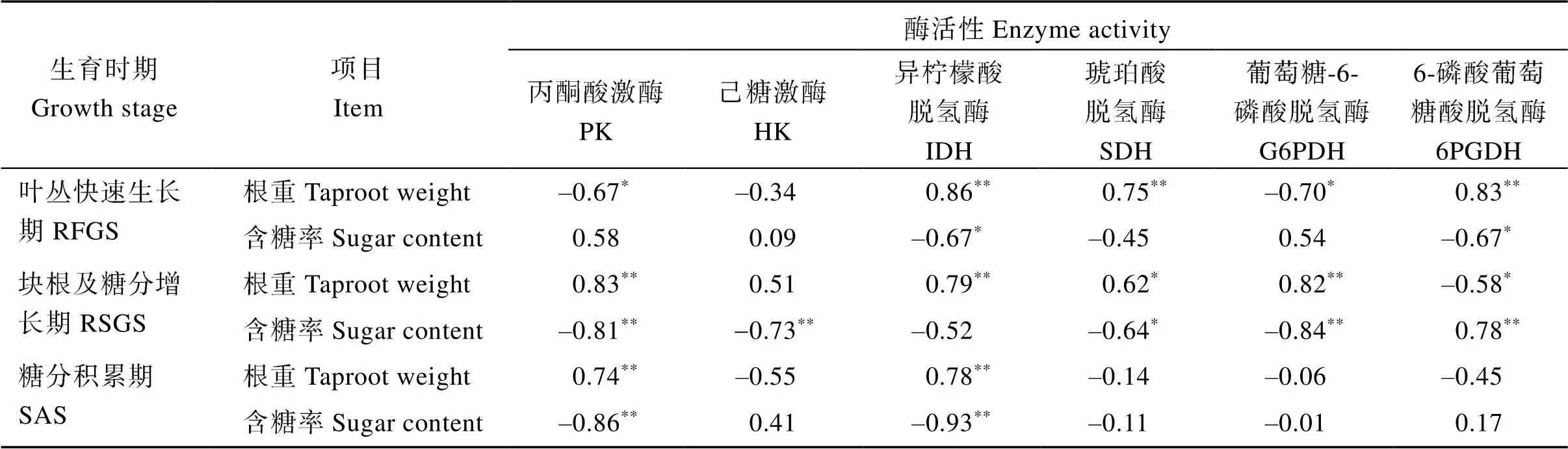
表3 甜菜呼吸代谢途径关键酶活性与块根重量和含糖率的相关系数
*、**分别表示在0.05和0.01概率水平显著相关。
*and**mean significant correlation in the 0.05 and 0.01 probability levels, respectively. SS, RFGS, RSGS, and SAS indicate seedling stage, rapid foliage growth stage, root and sugar growth stage and sugar accumulation stage, respectively.
喷施呼吸抑制剂均显著降低了甜菜块根的重量(图6-B)和直径(图6-D), 对根长无影响(图6-C)。对根重和根直径的降低程度也表现为丙二酸>碘乙酸>正磷酸钠, 说明呼吸作用主要通过促进块根膨大进而促进根的生长, 其中TCA呼吸途径在甜菜块根生长发育中起到重要作用。
2.4 不同基因型甜菜各生育时期块根能量代谢的差异
不同基因型甜菜各生育时期块根ATP合酶活性和ATP含量见图7, 各品种块根ATP合酶活性在块根及糖分增长期即块根生长最快速的时期达到峰值, ATP含量在叶丛快速生长期和块根及糖分增长期一直维持在较高水平, 且无论是ATP合酶活性还是ATP含量, 均是丰产型品种显著高于高糖型品种。进一步分析ATP合酶活性和ATP含量与甜菜块根重量与含糖的关系(表4)表明, 块根生长发育的主要3个生育时期ATP合酶活性和ATP含量与甜菜块根重量均呈正相关, 而与含糖率均呈负相关。说明呼吸作用为甜菜块根生长提供了能量基础, 而随着呼吸的加强, 也消耗了作为呼吸基质的糖类物质, 从而降低了甜菜块根的含糖率。

图5 不同基因型甜菜各生育时期呼吸代谢途径关键酶基因表达量差异
误差线上不同小写字母表示在0.05概率水平差异显著。不同生育时期的缩写同图1。
Bars superscripted by different lowercase letters are significantly different in the 0.05 probability level. Abbreviations for the different growth stages are the same as those given in Fig. 1.

图6 呼吸抑制剂对不同基因型甜菜块根呼吸速率和表型的影响
误差线上不同小写字母表示在0.05概率水平差异显著。
Bars superscripted by different lowercase letters indicate significantly different in the 0.05 probability level.
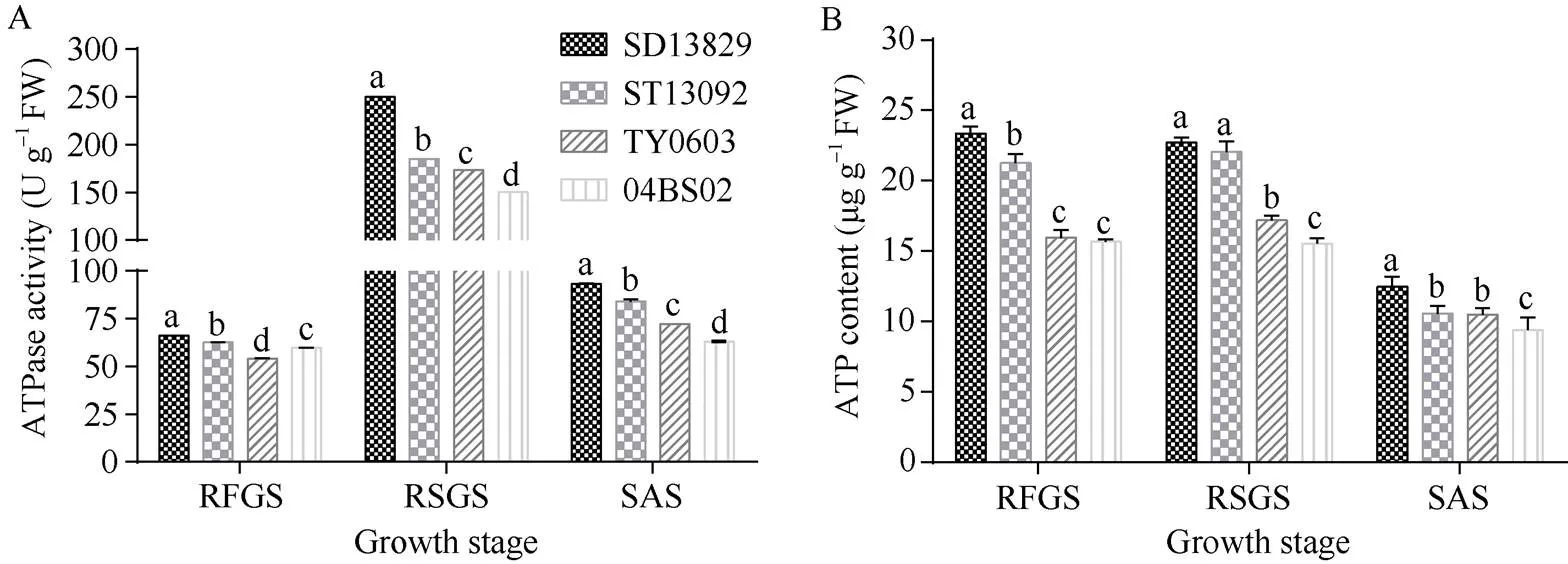
图7 不同基因型甜菜各生育时期块根能量代谢差异
误差线上不同小写字母表示在0.05概率水平差异显著。
Bars superscripted by different lowercase letters are significantly different at the 0.05 probability level.

表4 甜菜能量代谢与块根重量和含糖率的相关系数
*、**分别表示在0.05和0.01概率水平显著相关。
*and**mean significant correlation in the 0.05 and 0.01 probability levels, respectively. SS, RFGS, RSGS, and SAS indicate seedling stage, rapid foliage growth stage, root and sugar growth stage, and sugar accumulation stage, respectively.
3 讨论
3.1 不同基因型甜菜块根发育特性
本试验所选4个基因型甜菜块根鲜重和含糖率均随甜菜生育进程而增加, 除苗期外, 丰产型品种块根鲜重增长显著高于高糖型品种, 在块根及糖分增长期和糖分积累期丰产型品种块根含糖率显著低于高糖型品种。各类型品种根重和含糖增长规律与前人研究结果一致[28-29]。
3.2 呼吸代谢和能量代谢与甜菜块根生长的关系
呼吸作用的大小可反映植物体内代谢活动的强弱。不同植物、同一植物不同器官及不同发育时期的呼吸速率不同。较强的呼吸速率可以使植物产生的能量增加, 使植物在逆境条件下有足够能量来维持生长[30-31]。本研究发现不同基因型甜菜块根呼吸速率、ATP合酶和ATP含量呈生育前期较低, 中期较高, 后期又降低的变化规律, 并与甜菜块根膨大速率相一致。该结果与Moore等在木本植物上发现树干的总呼吸随着直径增长量的提高而提高, 并在达到高峰后, 随着生长量的降低而降低, 树干的生长呼吸与生长速率成正比的结论一致[32-33]。
3.3 呼吸代谢途径与甜菜块根生长的关系
植物体内主要有3种呼吸途径: EMP、TCA和PPP途径。本研究在每条呼吸途径中选用2个呼吸关键酶(EMP途径种的PK和HK; TCA循环中的IDH和SDH、PPP途径中6PGDH和G6PDH)的酶活来表示3种途径的代谢水平, 并利用呼吸抑制剂处理进行验证。结果证明EMP、TCA和PPP 3条呼吸代谢途径均参与不同基因型甜菜块根生长发育过程, 其中EMP-TCA途径为甜菜块根呼吸代谢主途径。李春萍[34]对核桃细根呼吸速率研究和白书农等[35]对水稻幼根进行呼吸代谢研究得出了TCA所占呼吸比重最高。
而TCA循环中的IDH酶活性和表达量与同时期甜菜块根根重规律一致, 在甜菜生长期酶活与块根重量成极显著正相关,基因相对表达量与其酶活性规律一致, 说明IDH酶活的升高是由转录水平直接调控的, TCA循环中的IDH对甜菜块根的生长发育起着重要作用。王雪峰[36]发现不同基因型甜菜的异柠檬酸脱氢酶相关蛋白存在差异表达说明异柠檬酸脱氢酶可以促进甜菜块根的膨大。时鹏涛[37]对转基因抗虫棉花生育前期叶片蛋白质组的比较分析也得出IDH蛋白参与TCA循环、蛋白质合成和碳固定等。还有学者通过究烟草硝酸还原酶缺失突变体[38-39]、番茄果实[40]、马铃薯[41]、油菜[42]和水稻[43-44]证明IDH参与氮吸收和氨基酸转运的功能, 是保持植物生长发育的重要因子。在后续研究中, 将对IDH进行生物信息学分析和表达特性分析,发掘在甜菜块根生长发育中起作用的IDHs基因; 并对其进行转化甜菜, 进一步研究这些基因在甜菜块根发育中的功能, 并探索通过基因工程手段改良甜菜。
4 结论
通过研究不同基因型甜菜各生育时期块根发育特性、呼吸速率、呼吸关键酶基因表达和酶活性各代谢水平的差异及其与根重和含糖的关系, 发现在甜菜块根发育过程中, 块根呼吸速率与块根重量呈极显著正相关, 表明呼吸作用在调控甜菜块根生长中具有重要作用。块根中ATP合酶、IDH、G6PDH活性与块根重量呈正相关关系。呼吸代谢相关酶活性及其基因表达分析和呼吸途径抑制剂实验表明, 三羧酸循环呼吸代谢途径对甜菜块根生长发育作用最大。研究结果为提高甜菜产量提供了新的调控策略。
[1] 刘佳, 薛惠云,李倩, 张志勇. 不同年代小麦品种根系形态差异性分析. 山东农业科学, 2021, 53(3): 15–21. Liu J, Xue H Y, Li Q, Zhang Z Y. Difference analysis of root morphology of wheat varieties among different eras., 2021, 53(3): 15–21 (in Chinese with English abstract).
[2] Robinson D. The responses of plants to non-uniform supplies of nutrients., 1994, 127: 635–674.
[3] 曲梦雪, 宋杰, 孙菁, 胡旦旦, 王洪章, 任昊, 赵斌, 张吉旺, 任佰朝, 刘鹏. 镉胁迫对不同耐镉型玉米品种苗期根系生长的影响. 作物学报, 2022, 48: 2945–2952. Qu M X, Song J, Sun J, Hu D D, Wang H Z, Ren H, Zhao B, Zhang J W, Ren B Z, Liu P. Effects of cadmium stress on root growth of maize (L.) varieties with different cadmium-tolerant at seedling stage., 2022, 48: 2945–2952 (in Chinese with English abstract).
[4] 张秋月. 复合胁迫对东北山樱根系生理生化特性的影响. 沈阳农业大学硕士学位论文, 辽宁沈阳, 2020. Zhang Q Y. Effects on Physiological and Biochemical Characteristics ofRoots under Combined Stress. MS Thesis of Shenyang Agricultural University, Shenyang, Liaoning, China, 2020 (in Chinese with English abstract).
[5] Van Der Merwe M J, Osorio S, Araujo W L, Albo I, Nunes-Nesi A, Maximova E, Carrari F, Bunik V I, Persson S, Fernie A R. Tricarboxylic acid cycle activity regulates tomato root growtheffects on secondary cell wall production., 2010, 153: 611–621.
[6] Howell P G, Levin E A, Springer A L, Kraemer S M, Phippard D J, Schief W R, Smith J D. Mapping a common interaction site used byDuffy binding-like domains to bind diverse host receptors., 2008, 67: 78–87.
[7] 于乔乔. 低温胁迫下玉米幼苗光合及呼吸代谢特性的研究. 东北农业大学硕士学位论文, 黑龙江哈尔滨, 2021. Yu Q Q. Study on the Characteristics of Photosynthesis and Respiratory Metabolism of Maize Seedlings under Low Temperature Stress. MS Thesis of Northeast Agricultural University, Harbin, Heilongjiang, China, 2021 (in Chinese with English abstract).
[8] 王军可. 灌浆初期高温影响粳稻籽粒碳氮代谢变化的机理及其调控. 中国农业科学院硕士学位论文,北京, 2021. Wang J K. Mechanism and Regulation of High Temperature at the Early Stage of Grain Filling on Changes in Carbon and Nitrogen Metabolism ofRice Grains. MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2021 (in Chinese with English abstract).
[9] 任晓松, 王子沐, 焦健, 田礼欣, 刘赵月, 李晶. GA3处理对低温胁迫条件下玉米种子呼吸代谢的影响. 生态学杂志, 2020, 39: 847–854. Ren X S, Wang Z M, Jiao J, Tian L X, Liu Z Y, Li J. Effects of GA3treatment on respiratory metabolism of maize seeds under low temperature stress., 2020, 39: 847–854 (in Chinese with English abstract).
[10] 刘丽杰, 苍晶, 李怀伟, 于晶, 王兴, 王健飞, 黄儒, 徐琛. 外源ABA对冬小麦越冬期呼吸代谢关键酶与糖代谢的影响. 麦类作物学报, 2013, 33: 65–72.Liu L J, Cang J, Li H W, Yu J, Wang X, Wang J F, Huang R, Xu C. Effects of exogenous abscisic acid on key enzymes of respiratory metabolism and sugar metabolism of winter wheat in the wintering period., 2013, 33: 65–72 (in Chinese with English abstract).
[11] 郭家鑫, 鲁晓宇, 陶一凡, 郭慧娟, 闵伟. 棉花在盐碱胁迫下代谢产物及通路的分析. 作物学报, 2022, 48: 2100–2144. Guo J X, Lu X Y, Tao Y F, Guo H J, Min W. Analysis of metabolites and pathways in cotton under salt and alkali stresses., 2022, 48: 2100–2144 (in Chinese with English abstract).
[12] 陈海浪, 陈喜文, 陈德富. NAD+-依赖型异柠檬酸脱氢酶的结构和功能研究进展. 生物技术通讯, 2003, 14: 304–307. Chen H L, Chen X W, Chen D F. Advance on structure and function of NAD+-dependent isocitrate dehydrogenase., 2003, 14: 304–307 (in Chinese with English abstract).
[13] 陈一鸣, 王涛, 张凤姣, 庄维兵, 束晓春,王忠, 杨清. 南方红豆杉野生种与栽培品种‘金锡杉’针叶代谢物的比较分析. 西北植物学报, 2019, 39: 801–807. Chen Y M, Wang T, Zhang F J, Zhuang W B, Shu X C, Wang Z, Yang Q. Comparison of metabolites in the needles ofvar., Mairei andvar., Mairei cv., ‘Jinxishan’.-, 2019, 39: 801–807 (in Chinese with English abstract).
[14] 张旭. 水分亏缺对小麦旗叶与穗部C3, C4光合途径关键酶及其中间产物代谢的影响. 西北农林科技大学硕士学位论文, 陕西杨凌, 2019. Zhang X. Effects of Water Deficit on C3and C4Photosynthesis Enzymes and Primary Carbon Metabolism in Wheat Flag Leaves and Spikes. MS Thesis of Northwest A&F University, Yangling, Shaanxi, China, 2019 (in Chinese with English abstract).
[15] 马春媚. 一氧化氮对桃果实三羧酸循环相关酶活性及蛋白质结构的影响. 山东农业大学硕士学位论文, 山东泰安, 2013. Ma C M. Effects of Nitric Oxide on Activities and Structure of Enzymes in Tricarboxylic Acid Cycle. MS Thesis of Shandong Agricultural University, Tai’an, Shandong, China, 2013 (in Chinese with English abstract).
[16] 李晓炜. 种子萌发过程中呼吸关键酶调控物质能量代谢规律研究. 兰州大学硕士学位论文,甘肃兰州, 2016. Li X W. The Law of Material and Energy Metabolism Regulated by the Key Enzyme of Respiration in the Process of Seed Germination. MS Thesis of Lanzhou University, Lanzhou, Gansu, China, 2016 (in Chinese with English abstract).
[17] Smiri M, Chaoui A, Ferjani E E. Respiratory metabolism in the embryonic axis of germinating pea seed exposed to cadmium., 2009, 166: 259–269.
[18] Peng Y, Niklas K J, Reich P B, Sun S. Ontogenetic shift in the scaling of dark respiration with whole-plant mass in seven shrub species., 2010, 24: 502–512.
[19] Zhang X, Wei X, Ali M, Rizwan H, Li B, Li H, Jia K, Yang X, Ma S, Li S. Changes in the content of organic acids and expression analysis of citric acid accumulation-related genes during fruit development of yellow (f.) and purple (f.) passion fruits., 2021, 22: 5765.
[20] Bailey-Serres J, Tom J, Freeling M. Expression and distribution of cytosolic 6-phosphogluconate dehydrogenase isozymes in maize., 1992, 30: 233–246.
[21] Hauschild R, Von Schaewen A. Differential regulation of glucose-6-phosphate dehydrogenase isoenzyme activities in potato., 2003, 133: 47–62.
[22] Huang J, Zhang H, Wang J, Yang J. Molecular cloning and characterization of rice 6-phosphogluconate dehydrogenase gene that is up-regulated by salt stressa., 2003, 30: 223–227.
[23] 王玮.基因调控甜菜块根发育的功能分析. 内蒙古农业大学博士学位论文, 内蒙古呼和浩特, 2021. Wang W. Functional Analysis ofGene in Regulating Taproot Development in Sugar Beet (L.). PhD Dissertation of Inner Mongolia Agricultural University, Hohhot, Inner Mongolia, China, 2021 (in Chinese with English abstract).
[24] 邵金旺, 蔡葆, 张家骅. 甜菜生理学. 北京: 农业出版社, 1991. pp 1–4. Shao J W, Cai B, Zhang J H. Sugar Beet Physiology. Beijing: Agriculture Press, 1991. pp 1–4 (in Chinese).
[25] 郇町, 胡华兵, 贺碧微, 孙琳琳, 刘建雄, 刘珣, 袁团团, 丁兆斐. 不同引进甜菜品种经济性状关系分析. 中国农学通报, 2023, 39(3): 11–19. Xun D, Hu H B, He B W, Sun L L, Liu J X, Liu X, Yuan T T, Ding Z F. Economic characters of introduced sugar beet varieties: relationship analysis., 2023, 39(3): 11–19 (in Chinese with English abstract).
[26] 吴珏, 谢文华, 徐淑婷, 曹锦萍, 王岳, 孙崇德. 高压静电场处理对椪柑采后贮藏性的影响. 浙江大学学报(农业与生命科学版), 2020, 46: 64–73. Wu J, Xie W H, Xu S T, Cao J P, Wang Y, Sun C D. Effect of high voltage electrostatic field treatment on the storability of postharvest ponkan fruit.(Agric Life Sci), 2020, 46: 64–73 (in Chinese with English abstract).
[27] 李志霞, 秦嗣军, 吕德国, 聂继云. 植物根系呼吸代谢及影响根系呼吸的环境因子研究进展. 植物生理学报, 2011, 47: 957–966. Li Z X, Qin S J, Lyu D G, Nie J Y. Research progress in root respiratory metabolism of plant and the environmental influencing factors., 2011, 47: 957–966 (in Chinese with English abstract).
[28] 郭文双. 内蒙古高寒地区甜菜品种适应性筛选与评价. 吉林农业大学硕士学位论文, 吉林长春, 2020. Guo W S. Adaptability Screening and Evaluation of Sugarbeet Varieties in Inner Mongolia. MS Thesis of Jilin Agricultural University, Changchun, Jilin, China, 2020 (in Chinese with English abstract).
[29] 朱芳慧. 甜菜表型特征及其与产质量关系的研究. 内蒙古农业大学硕士学位论文, 内蒙古呼和浩特, 2020. Zhu F H. Study on the Phenotypic Characteristics of Sugar Beet and Its Relationship with Yield and Quality. MS Thesis of Inner Mongolia Agricultural University, Hohhot, Inner Mongolia, China, 2020 (in Chinese with English abstract).
[30] 陈晓晶, 徐忠山, 赵宝平, 米俊珍, 严威凯, 刘景辉. 盐胁迫对燕麦根系呼吸代谢、抗氧化酶活性及产量的影响. 生态学杂志, 2021, 40: 2773–2782. Chen X J, Xu Z S, Zhao B P, Mi J Z, Yan W K, Liu J H. Effects of salt stress on root respiratory metabolism, antioxidant enzyme activity and yield of oat., 2021, 40: 2773–2782 (in Chinese with English abstract).
[31] 王天波, 赫文学, 张峻铭, 吕伟增, 梁雨欢, 卢洋, 王雨露, 谷丰序, 宋词, 陈军营. 人工老化玉米种胚ROS产生及ATP合成酶亚基mRNA完整性研究. 作物学报, 2022, 48: 1996–2006. Wang T B, He W X, Zhang J M, Lyu W Z, Liang Y H, Lu Y, Wang Y L, Gu F X, Song C, Chen J Y. ROS production and ATP synthase subunit mRNAs integrity in artificially aged maize embryos., 2022, 48: 1996–2006 (in Chinese with English abstract).
[32] Henry M, Besnard A, Asante W A, Eshun J, Adu-Bredu S, Valentini R, Bernoux M, Saint-Andrén L. Wood density, phytomass variations within and among trees, and allometric equations in a tropical rainforest of Africa., 2010, 260: 1375–1388.
[33] Moore S, Adu-Bredu S, Duah-Gyamfi A, Addo-Danso S D, Ibrahim F, Mbou A T, Grandcourt A, Valentini R, Nicolini G, Djagbletey G, Owusu-Afriyie K, Gvozdevaite A, Oliveras I, Ruiz-Jaen M C, Malhi Y. Forest biomass, productivity and carbon cycling along a rainfall gradient in West Africa., 2017, 24: 496–510.
[34] 李春萍, 王世伟, 丁俊杰,潘存德, 马彬, 努尔夏提·克里木江, 米热丁·艾海提, 祖力皮卡尔·吐松. 施氮水平对核桃细根呼吸速率及相关酶活性的影响. 林业科学研究, 2019, 32(6): 56–62. Li C P, Wang S W, Ding J J, Pan C D, Ma B, Nurxat K, Mihirdin A, Zulpikar T. Effects of nitrogen application level on respiration rate and related enzyme activities of walnut roots., 2019, 32(6): 56–62 (in Chinese with English abstract).
[35] 白书农, 肖翊华. 杂交水稻根系生长与呼吸强度的研究. 作物学报, 1988, 14: 53–59. Bai S N, Xiao Y H. Studies on the root growth and respiration of hybrid rice., 1988, 14: 53–59 (in Chinese with English abstract).
[36] 王雪峰. 基于iTRAQ技术对甜菜块根膨大蛋白质组学的研究. 内蒙古农业大学硕士学位论文, 内蒙古呼和浩特, 2016. Wang X F. The Proteomics Research of Tap Root Inflation in Sugar Beet (L.) by iTRAQ. MS Thesis of Inner Mongolia Agricultural University, Hohhot, Inner Mongolia, China, 2016 (in Chinese with English abstract).
[37] 时鹏涛. 转基因抗虫棉花不同发育期叶片蛋白质组的比较分析. 中国计量学院硕士学位论文, 浙江杭州, 2013. Shi P T. Comparative Proteomic Analysis of Transgenic Cotton leaves at Different Developmental Stages. MS Thesis of China Jiliang University, Hangzhou, Zhejiang,China, 2013 (in Chinese with English abstract).
[38] Scheible W R È, González F A, Morcuende R, Lauerer M, Geiger M, Glaab J, Gojon A, Schulze E D, Stitt M. Tobacco mutants with a decreased number of functionalgenes compensate by modifying the diurnal regulation of transcription, post-translational modification and turnover of nitrate reductase., 1997, 203: 304–319.
[39] Stitt M, Krapp A. The interaction between elevated carbon dioxide and nitrogen nutrition: the physiological and molecular background., 1999, 22: 583–621.
[40] Palomo J, Gallardo F, Suárez M F, Cánovas F M. Purification and characterization of NADP+-linked isocitrate dehydrogenase from scots pine: evidence for different physiological roles of the enzyme in primary development., 1998, 118: 617–626.
[41] Stitt M. Nitrate regulation of metabolism and growth., 1999, 2: 178–186.
[42] 陈喜文, 陈德富, 陈海浪.亚基1基因在油菜花粉与种子发育过程中的表达规律. 作物学报, 2005, 31: 838–843. Chen X W, Chen D F, Chen H L. The gene expression pattern ofsubunit 1 during the development of pollen and seed in., 2005, 31: 838–843 (in Chinese with English abstract).
[43] Kurai T, Wakayama M, Abiko T, Yanagisawa S, Aoki N, Ohsugi R. Introduction of thegene into rice enhances carbon and nitrogen assimilation under low-nitrogen conditions., 2011, 9: 826–837.
[44] 卢永恩. 水稻谷氨酸合酶基因和胞质异柠檬酸脱氢酶基因的功能研究以及氨基酸转运蛋白基因家族分析. 华中农业大学博士学位论文,湖北武汉, 2014. Lu Y E. Functional Analysis of Glutamic Synthase Gene and Cytosolic Isocitrate Dehydrogenase Gene and Genome-wide Analysis of Amino Acid Transporter Gene Family in Rice. PhD Dissertation of Huazhong Agricultural University dissertation, Wuhan, Hubei, China, 2014 (in Chinese with English abstract).
Characteristics of respiratory metabolism in growth and development of sugar beet taproot
YU Chao, LI Guo-Long, SUN Ya-Qing, LI Ning-Ning, and ZHANG Shao-Ying*
Agricultural College, Inner Mongolia Agricultural University, Hohhot 010019, Inner Mongolia, China
Respiration is an important metabolic basis for plant growth and development. The objective of this study is to compare the differences of taproot development characteristics, respiratory rate, respiratory key enzyme activity, gene expression, and energy change in different growth stages, and analyze the relationship between various respiratory physiological and sugar accumulation of sugar beet roots. In this study, four different genotypes of sugarbeet varieties were used as the experimental materials. The key enzyme activity and gene expression of respiratory metabolic pathway were determined, and respiratory pathway inhibitors were used at different growth stages. These results indicated the EMP-TCA was the main respiratory metabolic pathway during the growth process in beet taproot. No matter which respiratory pathway was inhibited, the weight and diameter of beet taproot can be reduced. Different respiratory metabolic pathways (EMP, TCA, and PPP) all played the important roles in promoting taproot expansion. In conclusions, this study laid a theoretical foundation for further research on the physiological mechanism of sugarbeet taproot, and provided physiological indicators for the breeding and identifying high-yield sugarbeet.
sugar beet; respiratory rate; respiratory enzyme activity; gene expression; ATP
2023-05-24;
2023-05-29.
10.3724/SP.J.1006.2023.34026
通信作者(Corresponding author): 张少英, E-mail: syzh36@aliyun.com
E-mail: yuchao19870618@sina.com
2023-02-10;
本研究由财政部和农业农村部国家现代农业产业技术体系建设专项(糖料, CARS-170201)和国家自然科学基金项目(31260347)资助。
This study was supported by the China Agriculture Research System of MOF and MARA (Sugar, CARS-170201) and the National Natural Science Foundation of China (31260347).
URL: https://kns.cnki.net/kcms2/detail/11.1809.S.20230526.1609.004.html
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜欢
中老年保健(2022年2期)2022-11-25 23:46:31
中老年保健(2021年7期)2021-12-02 16:50:22
黑龙江大学自然科学学报(2021年4期)2021-11-19 07:05:04
中老年保健(2021年4期)2021-08-22 07:09:10
小哥白尼(趣味科学)(2021年12期)2021-03-16 05:40:44
祝您健康(2020年1期)2020-04-02 17:17:19
中成药(2018年2期)2018-05-09 07:19:50
现代园艺(2017年19期)2018-01-19 02:50:01
中国糖料(2016年1期)2016-12-01 06:49:04
农业工程学报(2016年6期)2016-05-17 09:36:39
