
NAMPT可能通过烟酸代谢途径参与银屑病的发病
2023-11-13 05:37刘芳华刘守刚庄哲张泽乔苏杭陈永锋
皮肤性病诊疗学杂志 2023年5期
刘芳华, 刘守刚, 庄哲, 张泽乔, 苏杭, 陈永锋
1.广东医科大学,广东 湛江 524000;2.南方医科大学皮肤病医院,广东 广州 510091
银屑病是一种常见的慢性自身免疫性皮肤病,不仅影响皮肤,还累及身体的多个系统,甚至影响心理健康[1]。角质形成细胞及免疫细胞的异常反应在银屑病的发病机制中起关键作用[2]。自噬和烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide, NAD+)代谢是维持细胞稳态和能量代谢的重要调节因子,提高细胞内自噬活性或NAD水平可以恢复代谢活动[3-4]。线粒体自噬是一种特异性清除功能失调或多余的线粒体的自噬形式,在调节细胞因子的产生和免疫细胞发育、激活、分化以及稳态中起关键作用[5]。AMPK通过PINK1/Parkin促进了银屑病小鼠线粒体自噬的调节,抑制AMPK信号加重银屑病的皮肤炎症[6]。研究表明,NAD似乎是银屑病的潜在生物标记物,血清NAD水平的降低与促炎状态有关[7]。Dunnione增强细胞内NAD水平可以改善咪喹莫特诱导的小鼠银屑病皮炎[8]。在银屑病皮损部位局部外用NAD凡士林软膏后可减轻红斑、浸润、鳞屑[9]。自噬和NAD之间存在双向反馈调节机制,调节NAD水平可能对开发改善自噬受损相关疾病的新治疗策略具有重要意义[10]。目前,关于自噬和NAD+代谢在银屑病发病机制的相关性尚不确定。近年来,微阵列技术和综合生物信息学分析方法已广泛运用于鉴定和探索与疾病有关的关键基因和发病机制。本研究旨在探索银屑病中线粒体自噬的特征基因,并探讨与NAD+代谢途径及其相关基因的关系,以有助于更好地理解银屑病自噬的发病机制和潜在治疗策略的发展。
1 材料与方法
1.1 银屑病数据集、线粒体自噬基因和NAD+代谢途径相关基因的获取
从GEO(https://www.ncbi.nlm.nih.gov/geo)数据库中获取GSE30999(包含85例银屑病皮损和85例非银屑病皮损)数据集作为训练集,GSE79704(12例银屑病皮损和20例健康对照皮肤组织)数据集和GSE102725(10例银屑病皮损和10例非银屑病皮损)数据集作为验证集。从人类自噬基因数据库(http://www.autophagy.lu/)获取线粒体自噬基因222个。从KEGG数据库(https://www.genome.jp/kegg/,hsa00760)获取烟酸和烟酰胺代谢通路,从Reactome数据库(https://reactome.org/,R-HSA-196807)获取烟酸代谢通路,删除重复基因,将二者取合集,共获得烟酸代谢途径相关基因51个。
1.2 差异表达基因的获取
利用R语言软件(3.6.3版本)的“limma”包进行差异分析,获取GSE30999数据集中的差异表达基因,筛选条件为|log2FC|>1和P<0.05。绘制火山图和热图展示差异表达基因的分布。采用韦恩图用于获取具有差异表达的线粒体自噬基因、获取Lasso(least absolute shrinkage and selection operator)回归模型和Cox回归模型最佳基因的交集、获取具有差异表达的NAD+代谢途径的相关基因。柱状图用于展示基因的差异表达情况。R语言软件(3.6.3版本)的“ggplot”包用于数据可视化。
1.3 差异表达的线粒体自噬基因的ROC曲线
利用R语言软件(3.6.3版本)的“pROC”包获取差异表达的线粒体自噬基因的ROC曲线。R语言软件(3.6.3版本)的“ggplot”包用于数据可视化。ROC曲线的曲线下面积(area under the curve, AUC)>0.5为有意义。其中0.5
1.4 GO和KEGG分析
GO(gene ontology)是一个国际标准化的基因功能分类体系,依据标准词汇表全面地描述基因和基因产物在生物体中的属性。GO有3个本体,分别描述基因的分子功能(molecular function,MF)、细胞组分(cellular component,CC)、参与的生物过程(biological process, BP)。KEGG(kyoto encyclopedia of genes and genomes)可以系统地分析基因功能、链接基因组和功能信息的数据库,包括基因和基因组数据库、分层分类数据库和代谢通路数据库等。利用R语言软件(3.6.3版本)的“clusterProfiler”包进行富集分析。
1.5 相关性分析
使用R语言软件(3.6.3版本)的“performanceAnalytics”“Hmisc”“pROC”和“corrplot”包分析16个线粒体自噬基因之间的相关性、烟酰胺磷酸核糖转移酶(nicotinamide phosphoribosyl transferase, NAMPT)与NAD+代谢途径及其相关基因的相关性。皮尔逊相关系数的标准定义:0.00-0.19:极弱;0.20-0.39:弱;0.40-0.59:中度;0.60-0.79:强;0.80-1.0:极强。
1.6 Lasso回归模型和Cox比例风险模型
利用数据分析Sangerbox 平台 (http://www.sangerbox.com/tool),使用Lasso-cox回归分析工具筛选银屑病线粒体自噬特征基因;使用COX模型计算调整后的风险比值(hazard ratio, HR),通过HR评估线粒体自噬基因和银屑病之间的关系。使用R语言软件(3.6.3版本)的“survival”“survminer”“tidyverse”和“eoffice”包进行Cox比例风险建模。
1.7 GSEA分析
在Sangerbox 工具中使用“单基因GSEA分析”, 探索NAMPT基因的调控通路和分子功能富集情况。
1.8 评估NAD+代谢途径状态
使用R语言软件(3.6.3版本)的“GSVA”包进行了单样本基因集富集分析(single sample gene set enrichment analysis, ssGSEA)以评估NAD+代谢途径在银屑病中的状态。
2 结果
2.1 银屑病中差异表达线粒体自噬基因的鉴定和评估
对GEO银屑病数据集GSE30999使用“limma”包分析,获得差异表达基因1 407个,其中上调基因871个,下调基因536个(图1A)。在人类自噬基因数据库获得线粒体自噬基因222个,与GSE30999数据集中的差异表达基因取交集后,获得16个差异表达的线粒体自噬基因(图1B)。16个线粒体自噬基因差异表达分析结果显示,与对照组相比表达均增高(图1C、1E)。ROC曲线显示PRKCQ(AUC=0.988)、NAMPT(AUC=0.965)、CASP1(AUC=0.980)、APOL1(AUC=0.962)、BIRC5(AUC=0.948)、SERPINA1(AUC=0.945)、EIF4EBP1(AUC=0.956)、SESN2(AUC=0.963)、 IKBKE(AUC=0.948)、CTSD(AUC=0.941)、RAB24(AUC=0.929)、ATG2A(AUC=0.907)、 CCL2(AUC=0.887)、FKBP1B(AUC=0.893)、IL24(AUC=0.930)和RGS19(AUC=0.755)。表明16个线粒体自噬基因在银屑病中均具有较好的诊断价值(图1D、1F)。

图1 筛选和验证银屑病中差异表达的线粒体自噬基因 1A:火山图显示GSE30999数据集中的差异表达基因;1B:韦恩图显示GSE30999数据集中的差异表达的线粒体自噬基因;1C、1E:16个线粒体自噬基因在GSE30999数据集的差异表达情况;1D、1F:16个线粒体自噬基因在GSE309999数据集中的ROC曲线Figure 1 Screening and validation of differentially expressed mitophagy genes in psoriasis 1A:Volcano plot showing differentially expressed genes in GSE30999 dataset; 1B: Venny plot showing the differentially expressed mitophagy genes in the GSE30999 dataset; 1C,1E: Differential expression of 16 mitophagy genes in GSE30999 dataset; 1D,1F: ROC curves for 16 mitophagy genes in GSE30999 dataset.
2.2 线粒体自噬基因的富集分析和相关性分析
进一步分析银屑病中差异表达的16个线粒体自噬基因的表达和分布情况。火山图和热图(图2A、2D)显示16个线粒体自噬基因在银屑病皮损中均呈不同程度的高表达。相关性分析(图2C)显示16个自噬基因之间相关性密切。其中,NAMPT与CASP1相关性为0.81,CASP1与APOL1相关性为0.80, EIF4EBP1与APOL1和BIRC5相关性均为0.81。富集分析结果显示,16个线粒体自噬基因的GO分析主要富集在一氧化氮合酶生物合成过程的正向调控、一氧化氮合成酶生物合成工艺、一氧化氮合酶生物合成过程的调控、内肽酶活性的调节、肽酶活性的调节、半胱氨酸型内肽酶调节活性参与凋亡过程、肽酶调节活性、细胞因子活性、半胱氨酸型内肽酶活性、受体配体活性。KEGG富集分析主要为NOD样受体信号通路、自噬-动物、甲型流感、化学致癌-受体激活和脂质与动脉粥样硬化通路上(图2B、表 1)。

图2 银屑病差异表达的线粒体自噬基因的相关性分析和富集分析 2A、2D:火山图和热图显示16个线粒体自噬基因的表达和分布;2B:气泡图展示了16个线粒体自噬基因的功能和通路富集分析情况;2C:矩阵图展示了16个线粒体自噬基因之间的相关性Figure 2 Correlation and enrichment analyses of differentially expressed mitophagy genes in psoriasis. 2A,2D: Volcano plot and a heat map showing expression and distribution of 16 mitophagy genes;2B: Bubble plots demonstrate functional and signaling pathway enrichment analysis of 16 mitophagy genes;2C: Matrix plots present the association among the 16 mitophagy genes.
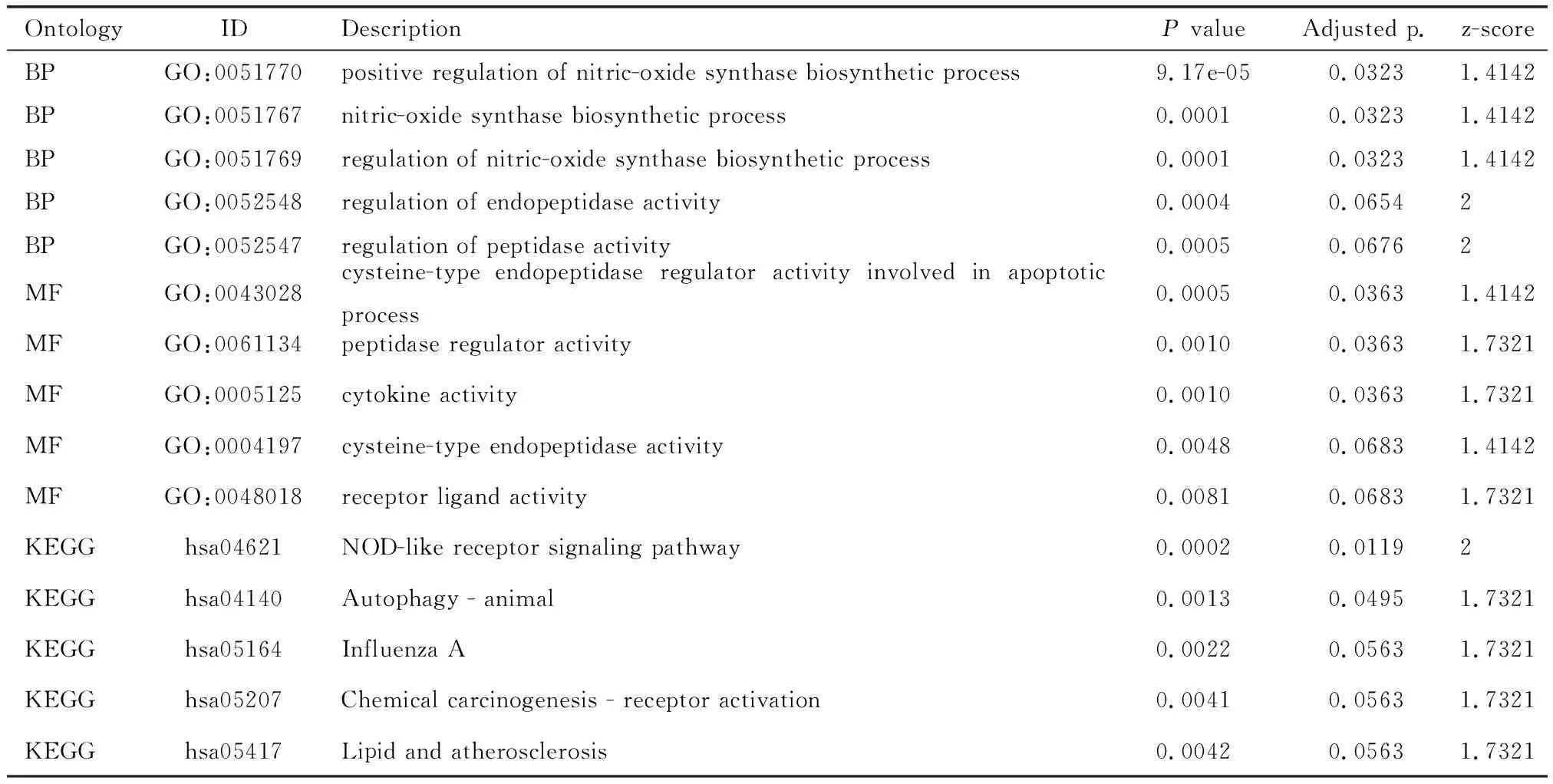
表1 16个线粒体自噬基因的富集分析Table 1 Enrichment analysis of 16 mitochondrial autophagy genes
2.3 线粒体自噬基因NAMPT的筛选及其在GSEA中的富集分析
使用两种不同的算法来筛选特征基因。通过Lasso回归算法缩小差异表达的线粒体自噬基因的范围,共筛选了11个基因(图3A)。基于LASSO的λ值,由11个具有LASSO系数的基因构建了基于基因的生存风险评分模型RiskScore。筛选公式(LASSO系数乘以基因表达量之和)如下:RiskScore=-0.546231539729713*PRKCQ-0.258825070629174*NAMPT-0.139354953296213*CASP1-0.0249556420497596*APOL1-0.0219626226300222*SERPINA1-0.203740011478795*SESN2-0.0690735086804116*IKBKE-0.10810713861763*RAB24-0.125345037556547*ATG2A-0.015930181451998*FKBP1B+0.0700684193781504*RGS19。

图3 银屑病线粒体自噬特征基因 3A、3B:Lasso和Cox回归模型筛选最佳基因;3C:韦恩图显示Lasso和Cox回归模型的交集Figure 3 Mitophagy signature genes in psoriasis. 3A,3B: Lasso and Cox regression models to screen the most robust genes; 3C: Venny plot showing the intersection of the Lasso and Cox regression models.
通过COX回归模型,最终选择了这两种算法的1个重叠基因NAMPT(HR:1.22,95%CI:0.91~1.8,P=0.013)(图3B、3C)。在GSE79074数据集和GSE102725数据集中验证NAMPT mRNA的表达,结果显示NAMPT mRNA 均高表达(图4A、4B)。GSEA富集分析显示NAMPT主要富集在蛋白酶体、嘧啶代谢、泛素介导的蛋白水解、DNA复制、抗原处理和呈递、氨酰基 TRNA生物合成、细胞质DNA感知途径、卵母细胞减数分裂、同源重组、TOLL样受体信号通路(图4C、4D)。

图4 NAMPT的差异表达情况和GSEA分析 4A、4B:GSE79704数据集和GSE102725数据集验证NAMPT的差异表达;4C、4D:GSEA分析显示NAMPT的富集情况Figure 4 The differential expression and GSEA analysis of NAMPT. 4A,4B: Using GSE79704 and GSE102725 datasets to validate differential expression of NAMPT; 4C,4D: GSEA analysis reveals the enrichment of NAMPT.
2.4 NAD+代谢途径在银屑病中处于激活状态,NAMPT可能与NAD+代谢途径的激活有关
对KEGG和Reactome数据库的烟酸和烟酰胺代谢通路基因取交集,获得NAD+代谢途径相关基因共51个。通过ssGSEA分析,发现NAD+代谢途径在银屑病中处于激活状态(图5A),并与NAMPT呈正相关(图5B)。将NAD+代谢途径相关基因与差异表达基因取交集(图5C),共获得4个基因,分别为NAMPT、BST1、PARP9和PNP,与对照组相比,BST1呈低表达(图5D)、PARP9(图5E)和PNP(图5F)呈高表达。相关性分析显示,NAMPT与BST1呈负相关(图5G)、与PARP9(图5H)和PNP(图5I)呈正相关。说明在银屑病皮损中,NAD+代谢途径处于激活状态,NAMPT与NAD+代谢途径的激活正相关,且NAMPT与NAD+代谢途径中BST1、PARP9 和PNP密切相关。

图5 NAMPT与NAD+代谢途径的关系 5A:NAD+代谢途径在银屑病中处于激活状态;5B:NAD+代谢途径与NAMPT之间的相关性;5C:韦恩图显示了GSE30999数据集中差异表达的基因和NAD+代谢途径相关基因的交集;5D~5F:BST1、PARP9和PNP在GSE30999数据集中的差异表达; 5G~5I:NAMPT与BST1、PARP9和PNP的相关性Figure 5 Relationship between NAMPT and NAD+ metabolic pathway. 5A: The NAD+ metabolic pathway is activated in psoriasis; 5B: Correlation between NAD+ metabolic pathway and NAMPT; 5C: Venn diagram shows the intersection of DEGs and NAD+ metabolic pathway-related genes in the GSE30999 dataset; 5D~5F: Differential expression of BST1, PARP9 and PNP in psoriasis; 5G~5I: Correlation between NAMPT and BST1, PARP9 and PNP.
3 讨论
银屑病是一种慢性复发性炎症性皮肤病,由表皮角质形成细胞、真皮血管细胞和免疫细胞之间的相互作用介导[11]。有研究发现自噬异常导致角质形成细胞产生炎症细胞因子和细胞增殖,突出了自噬在银屑病发病中的重要作用;同时揭示了TLR2/6或TLR4刺激触发自噬途径并增加原代人角质形成细胞中p62表达的证据;此外,抑制自噬增强了原代人角质形成细胞中炎症细胞因子的产生和p62的表达[12]。另有研究表明,一种涉及ATG5和高尔基体重组堆叠蛋白2 (GORASP2)的自噬依赖非常规分泌途径(自分泌)会增加银屑病角质形成细胞炎症[13]。自噬缺陷可能会加重炎症和角质形成细胞增殖,抑制自噬会放大银屑病中的皮肤炎症,使银屑病加重[14-16]。这表明自噬在银屑病中受损,也提示自噬在疾病发病机制中的作用。
本研究在GSE30999数据集中筛选了16个差异表达的线粒体自噬基因,并在银屑病皮损组织中均高表达。富集分析显示KEGG主要为NOD样受体信号通路、自噬-动物、甲型流感、化学致癌-受体激活和脂质与动脉粥样硬化通路上。在经过差异表达基因筛选、LASSO回归模型和Cox回归模型的筛选后,只有NAMPT为最佳基因。NAMPT在银屑病中高表达,且NAMPT对银屑病的诊表现出极高的敏感性和特异性(AUC=0.965)。因此推测NAMPT可能在银屑病的发病机制中起着重要作用。
NAMPT是细胞内NAD+代谢途径的调节因子[17]。NAD是参与细胞氧化还原反应的重要辅酶,是NAD依赖酶的底物[18]。NAD动态平衡的失调与炎症和疾病有关,在各种代谢紊乱和衰老过程中,NAD水平下降[19]。NAMPT通过其NAD生物合成活性,影响NAD依赖酶的活性,从而调节细胞代谢[17]。除了酶促功能外,胞外NAMPT(extracellular, eNAMPT)还具有细胞因子样活性[20]。异常的eNAMPT水平与各种代谢紊乱、炎症和自身免疫性疾病有关,如肥胖、胰岛素抵抗和类风湿关节炎有关[20-21]。NAMPT能够通过影响氧化应激反应、细胞凋亡、脂质和葡萄糖代谢、炎症和胰岛素抵抗来调节肥胖和相关疾病(如非酒精性脂肪性肝病和2型糖尿病)的发病[22-24]。最近的一项研究表明,银屑病患者皮损中NAD含量增加,并与高NAMPT转录水平相关[25]。Th1/ Th17细胞因子强烈诱导细胞内NAMPT表达,并诱导角质形成细胞过度增殖。NAMPT介导的NAD+代谢促进了银屑病皮肤中常驻皮肤细胞的免疫反应。本研究探讨NAMPT与NAD+代谢途径及其相关基因的关系,结果发现NAMPT mRNA在银屑病中高表达,且NAD+代谢途径处于激活状态,NAMPT与NAD+代谢途径的激活呈正相关。在NAD+代谢途径中,发现3个基因(BST1、PARP9和PNP)在银屑病中呈现差异表达,BST1呈低表达、PARP9和PNP呈高表达。NAMPT与BST1呈负相关、与PARP9和PNP呈正相关。BST1可以将烟酰胺核糖水解为烟酰胺,有助于NAD的产生[26]。反过来,抑制PNP的活性增强了烟酰胺核糖的NAD合成[27]。NAMPT是NAD途径中的限速酶。在胰腺癌中,IFN上调PARP9的表达诱导NAD的消耗增加了对NAMPT的依赖,以防止NAD的严重消耗[28]。说明NAMPT可能在银屑病中激活了NAD+代谢途径,并可能通过BST1、PARP9或PNP来发挥作用。
综上所述,NAMPT作为线粒体自噬基因在银屑病皮损中高表达,对于银屑病具有极高的诊断价值。同时,NAD+代谢途径处于激活状态,NAMPT与NAD+代谢途径的激活密切相关,并与BST1、PARP9或PNP密切相关。NAMPT与NAD+代谢之间的调节作用可能是银屑病治疗的潜在治疗策略。但本研究中GSE79704和GSE102757验证组的病例数较少,从3个数据集获得的特征基因及其与NAD+代谢途径的关系需要进一步通过体内和体外实验验证其可重复性,自噬和NAD+代谢相关的具体机制亦需进一步探讨。
猜你喜欢
海洋通报(2021年1期)2021-07-23
中国民间疗法(2021年1期)2021-04-20
生物学通报(2021年4期)2021-03-16
中国医学科学院学报(2015年5期)2015-03-01
中华皮肤科杂志(2014年4期)2014-12-19
中华皮肤科杂志(2014年4期)2014-12-19
中华皮肤科杂志(2014年3期)2014-12-19
浙江中西医结合杂志(2014年10期)2014-07-07
癌变·畸变·突变(2014年1期)2014-03-01
现代检验医学杂志(2014年3期)2014-02-02
