
基因片段Myo5a原核表达、纯化及多克隆抗体制备①
2023-11-13 09:29游荷花唐锐敏山西医科大学汾阳学院免疫学与免疫学检验教研室汾阳032200
中国免疫学杂志 2023年10期
游荷花 唐锐敏 (山西医科大学汾阳学院免疫学与免疫学检验教研室,汾阳 032200)
Myosin 是肌球蛋白,又称肌凝蛋白,是真核细胞里的一类ATP依赖型分子马达[1]。Myosin于1864年由KUEHNE 在对肌肉的运动进行探讨时发现的,以肌动蛋白丝作为运转的轨道,经过水解ATP 造成了构型的改变而在肌动蛋白丝上移行。最初研究以为其是存在于平滑肌和横纹肌中的ATP 酶,之后陆续发现许多真核细胞也存在同源蛋白基因,证实这种蛋白的用处不局限于肌肉细胞。Myosin 可以进行的细胞活动包括:肌肉运动、趋化作用、细胞分裂、胞饮作用、靶向小泡运输和信号转导等,是生物体内重要的蛋白质成员。据发现时间的先后命名肌球蛋白,目前已发现30 多种,分为传统的和非传统的[2]。
Myo5 也称为Myosin5 或MyoⅤ,在细胞中物质运输中有重要的作用。MyosinⅤa(Myo5a)是MyosinⅤ家族中的一分子,是一种“摩托蛋白”,主要负责转运细胞内物质[3]。近十几年不断有研究发现Myo5a与某些肿瘤的发生发展关系密切。为进一步探究Myo5a 可能与某些肿瘤发展的关系,本实验拟制备Myo5a 片段的原核表达质粒PGEX-4T-3/Myo5a,使其表达并纯化该蛋白质,制备其抗血清,为后续探讨Myo5a与肿瘤的关系作铺垫。
1 材料与方法
1.1 材料 Myo5a PCR 模板、原核载体、ELISA 反应板、实验动物、佐剂以及E.coliDH5α和BL21均由实验室提供;DNA 限制性核酸内切酶BamHⅠ和XhoⅠ、T4DNA连接酶、DNA聚合酶等均购于TaKaRa公司;镍亲和层析柱购自Bio-Rad 公司;低分子量蛋白标准Marker 和DNA Marker 等购自北京全式金生物技术有限公司。
1.2 方法
1.2.1 Myo5a 目的基因的获取 按照人Myo5a 基因序列,设计一对引物来扩增Myo5a 羧基末端的近100 bp 大小的基因小片段,命名为Myo5a-D1[BamHⅠF(5'~3'):CG/GGATCC/ATGAATAGTGGGGCTAAAG;XhoⅠR(5'~3'):CCC/CTCGAG/AGAACAAATGGCTTCTGCATC,长度111 bp]。BamHⅠ是正向引物的酶切位点,XhoⅠ是反向引物的酶切位点。扩增条件如下:95 ℃ 预变性2 min;95 ℃变性15 s、54 ℃退火30 s、72 ℃延伸45 s,共30 个循环;最后4 ℃保温。扩增完毕,进行琼脂糖电泳鉴定。采用RCR纯化试剂盒纯化基因小片段。
1.2.2 PGEX-4T-3/Myo5a 重组质粒的构建及鉴定 原核表达载体PGEX-4T-3 的基因片段经BamHⅠ/XhoⅠ双酶切反应(37 ℃、CO2孵箱过夜),用DNA回收试剂盒回收所需DNA。将原核表达载体PGEX-4T-3 与Myo5a-D1 连接, EP 管中加入1.5 µl PGEX-4T-3 质粒、0.3 µl D1、0.4 µl T4DNA 连接酶、1 µl缓冲液后补加去离子水(DI H2O)使连接体系为10 µl,4 ℃冰箱过夜。连接产物转化到大肠杆菌的DH5α 感受态细胞,37 ℃培养,筛选重组子。经双酶切鉴定和测序,获得重组质粒PGEX-4T-3/Myo5a-D1。
1.2.3 重组PGEX-4T-3/Myo5a 的诱导表达 4 µl PGEX-4T-3(对 照)、8 µl 重 组 质 粒PGEX-4T-3/Myo5a-D1转化入大肠杆菌BL21。置于冰中30 min,42 ℃热击40 s,再冰镇1~2 min。经复苏后,取适量涂布于含有Amp 的LB 盘中,37 ℃孵箱放置12~16 h后,可见菌落长出。挑取阳性克隆菌落,接于2 ml含有Amp 的LB 培养基中(两个培养皿依次挑取)。37 ℃摇床振荡3~4 h,至OD 值为0.6。各取0.5 ml菌液作为诱导前的对照。剩余菌液均加15 µl IPTG,37 ℃摇床中培养至终浓度为1 mmol/L(2~3 h)。培养结束后,1.5 ml 菌液均分成3 份。其中0.5 ml 菌液保种储存;0.5 ml 菌液为诱导后的总菌液,离心,收集沉淀加50 µl PBS重悬菌液;最终0.5 ml菌液,离心,收集沉淀后加250 µl的PBS重悬菌液,超声裂解细菌细胞,离心后分别得到细菌沉淀和裂解上清,去离子水溶解沉淀。最终得到质粒包含诱导前总菌液、诱导后总菌液、裂解上清、细菌沉淀共4 个样品。利用聚丙烯酰胺凝胶电泳检测蛋白的表达。
1.2.4 大量表达重组PGEX-4T-3/Myo5a 蛋白 LB培养基(含Amp)中接种1 个BL21 转化成功的阳性克隆,37 ℃ 过夜培养,第2天再加入300 ml LB,继续摇菌至OD 值约为0.6。按1∶100 的比例加入IPTG培养至终浓度为1 mol/L。4 ℃离心2 次,弃上清。10 ml的去离子水重悬菌液,分装10管,-20 ℃储存。
1.2.5 采用镍亲和层析柱提纯蛋白,Western blot检测纯化蛋白 取1 ml 菌体,加29 ml 去离子水,50 µl PMSF 和1% Triton。超声裂解细胞,4 ℃离心,得到裂解上清,滤纸过滤。层析柱中加0.5 ml 50%谷胱甘肽琼脂糖,样品过柱(前后均用5×柱体积的PBS 洗层析柱)。采用3×柱体积的洗脱液洗脱蛋白,得到提纯的融合蛋白。收集蛋白0.5 ml/管,-80 ℃储存。其中20 µl 样品SDS-PAGE 电泳鉴定。PGEX-4T-3/Myo5a-D1 得到了大量提纯的蛋白,采用Western blot 检测提纯的蛋白,并用SDS-PAGE 电泳纯化后的融合蛋白和阴性对照,在80 V 恒压状态下转膜2 h。4 ℃过夜后在NC 膜条中加入一抗,4 ℃过夜,TBST 洗涤5 次。加二抗孵育,洗膜。最后显色记录结果。
1.2.6 制备兔抗人抗血清,并对其进行滴度测定查阅文献[4],采用PGEX-4T-3/Myo5a-D1 融合蛋白为免疫原,免疫动物选适龄,健康的家兔。初次注射免疫原剂量为200 µg,佐剂用完全弗氏佐剂,佐剂与免疫原按1∶1 体积比混匀,于家兔皮下注射。注射3 周后,再次注射进行加强免疫, 家兔皮下均用100 µg PGEX-4T-3/Myo5a-D1 融合蛋白,与等体积的不完全弗氏佐剂混匀后注射。之后每隔2~3周进行加强免疫1 次,总共注射4 次。在最后一次注射后第6 天进行静脉采血,ELISA 法测血清抗体滴度。酶标板先用PGEX-4T-3/Myo5a-D1 融合蛋白(10 µg/ml,100 µl/孔)4 ℃过夜包被,后用10%牛血清白蛋白37 ℃进行封闭2 h,之后洗板5次。加入梯度稀释的家兔抗血清(100 µl/孔),恒温孵箱37 ℃放置1 h,阴性对照用家兔未免疫时的血清。后加入HRP-羊抗兔IgG 后,恒温孵箱37 ℃放置1 h。加入显色底物TMB 恒温孵箱37 ℃放置0.5 h 后加中止液。酶标仪检测波长450 nm 处OD 值,判定抗血清效价(Cut off 值为阴性孔OD450值×2.1,大于此值的抗血清最大稀释度为其效价)。
1.2.7 抗血清特异性鉴定 Western blot 检测:将纯化的PGEX-4T-3/Myo5a-D1 蛋白经SDS-PAGE 电泳,后转移至NC 滤膜上。制备的兔抗血清与膜4 ℃孵育过夜,加二抗,孵育,洗涤,显色并记录结果。免疫荧光法检测:脑神经细胞(因富含Myo5a 蛋白)作为基质细胞切片,加制备的兔抗血清于4 ℃,过夜,阴性对照还是用兔未免疫时的血清,加二抗,37 ℃孵育0.5 h后洗涤5次,最后荧光显微镜下观察发射荧光情况。
2 结果
2.1 扩增了基因小片段My05a-D1 以人类Myo5a基因为模板,利用引物,成功扩增了基因小片段Myo5a-D1,PCR结果见图1。

图1 Myo5a-D1小片段的琼脂糖凝胶电泳图Fig.1 Agarosegel electrophoresis of small fragment of Myo5a-D1
2.2 PGEX-4T-3/Myo5a 重组质粒构建及鉴定 将在抗性平板上生长的菌落随机挑取进行摇菌培养,提取重组质粒进行PCR 鉴定,均含目的条带。质粒DNA 双酶切鉴定结果见图2。将鉴定正确的重组质粒进行测序,测序结果符合预期。

图2 PGEX-4T-3/Myo5a-D1重组质粒的酶切鉴定Fig.2 Enzymatic digestion of PGEX-4T-3/Myo5a-D1 recombinant plasmid
2.3 融合蛋白诱导表达 选重组质粒PGEX-4T-3/Myo5a-D1 与 空 载 体PGEX-4T-3 进 行IPTG 诱 导 表达,聚丙烯酰胺凝胶电泳检测蛋白表达(图3)。PGEX-4T-3/Myo5a-D1 经诱导后在裂解上清、沉淀、诱导后的总菌液中均产生分子质量约30 kD 的大量蛋白。与对照质粒PGEX-4T-3 表达的GST 蛋白(分子质量为29 kD)略高。表明融合蛋白诱导表达成功。
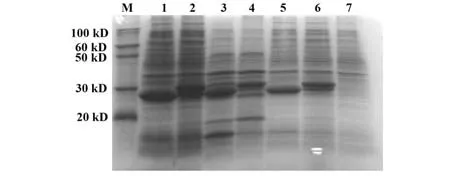
图3 PGEX-4T-3/Myo5a-D1的原核表达产物的电泳鉴定Fig.3 Electrophoretic identification of prokaryotic expression product of pGEX-4T-3/Myo5a-D1
2.4 融和蛋白的纯化鉴定 将裂解后上清采用镍亲和层析柱纯化,结果见图4,PGEX-4T-3/Myo5a-D1得到大量纯化的蛋白。对PGEX-4T-3/Myo5a-D1 进行Western blot 鉴定,见图5。显色条带大小约为30 kD,符合预期,表明目的蛋白表达成功。

图4 蛋白纯化的电泳鉴定Fig.4 Electrophoretic identification of protein purification

图5 纯化蛋白的免疫印迹鉴定Fig.5 Western blot identification of purified protein
2.5 多克隆抗体鉴定 包括滴度和特异性的测定。PGEX-4T-3/Myo5a-D1 融合蛋白包被酶标板,ELISA 间接法检测制备的兔抗人PGEX-4T-3/Myo5a-D1抗血清滴度为1∶128 000。将兔抗人PGEX-4T-3/Myo5a-D1抗血清(1∶100 00)作为一抗,与PGEX-4T-3/Myo5a-D1融合蛋白反应,以Western blot 实验检测其抗原特异性,见图6。结果表明,在分子量30 kD左右显现特异条带,说明兔抗人PGEX-4T-3/Myo5a-D1 多克隆抗体具有良好的抗原特异性。脑组织细胞因富含Myo5a 在荧光显微镜下出现绿色荧光,阴性对照无荧光(图7)。证明制备的抗血清与天然的Myo5a蛋白发生特异性反应。

图6 Western blot检测兔抗血清Fig.6 Western blot detection of rabbit antiserum

图7 免疫荧光染色Fig.7 Immunofluorescence staining
3 讨论
目前发现Myosin 中的很多类型都和各型肿瘤发生及发展关系密切。可能原因是传统的Myosin的主要功能是参与肌肉收缩,而非传统的Myosin 是细胞骨架的组成,为细胞质流动、物质运输、有丝分裂、胞质裂解等提供所需的能源[5]。癌症主要生物学特征是癌细胞转移,癌细胞转移涉及过程极其复杂,但有研究认为癌细胞转移运动需要细胞骨架提供支撑与动力[6]。目前研究比较多的是非传统的Myosin2、Myosin9 和Myosin10 等。研究显示非传统的Myosin2 与消化道肿瘤、乳腺癌、膀胱癌、肺癌等肿瘤有紧密的关系,也有研究表明适当的T 细胞反应和T细胞特异性抗原受体相互作用的强度与细胞骨架重组有关,丝状肌动蛋白和非肌肉肌球蛋白Ⅱ(NMMⅡ)起主要作用[7-8]。Myosin9是近年新发现的癌细胞转移相关蛋白,研究显示,Myosin9 可促使胃癌细胞向腹膜转移[9]。同时Myosin10可导致多种疾病,包括恶性肿瘤和病原微生物感染。与正常结直肠组织相比,结直肠癌组织中Myosin10 表达会明显升高,推测Myosin10 可能与结直肠癌的发生发展关系密切[10]。除结直肠癌外,也有研究报道Myosin10在乳腺癌和非小细胞肺癌等几种侵袭性恶性肿瘤中高表达[11]。此外,乳腺癌患者的预后不良也与高水平的Myosin10 有关。而关于Myosin5 近十几年也陆续发现其与某些肿瘤的发生发展关系密切,因此本研究选择Myosin5作为切入点。
Myosin5 是迄今为止在细胞功能、酶性质和调控方面最具特色的非传统肌球蛋白。脊椎动物有3 个Myosin5 基 因,分 别 编 码Myo5a、Myo5b 和Myo5c[12]。Myosin5 参与了许多膜质载体:如分泌囊泡、液泡、黑素体、色素颗粒、循环核内体等和mRNA的运输[13]。因此脊椎动物Myo5a的调控也已经受到了广泛的关注。文献显示Myo5a高表达与肿瘤浸润深度显著相关。有文献报道食管鳞状细胞癌患者中Myo5a表达水平可用于预测肿瘤的侵袭性及预测患者的临床生存期,可用于指导临床诊疗方案和评估疗效,此外,Myo5a在其他许多恶性肿瘤中高表达并与癌细胞侵袭相关,例如胰岛细胞瘤、乳腺癌、结肠癌、肺癌、前列腺癌和恶性黑素瘤等[14-15]。因此,近年来对Myo5a 的运动和循环都进行了较深入的探讨。
本文采用PCR扩增了Myo5a末端特异性片段的蛋白编码序列,用BamHⅠ和XhoⅠ双酶切,用原核表达载体pGEX-4T-3 将片段插入,构建了重组质粒PGEX-4T-3/Myo5a-D1。原核重组质粒PGEX-4T-3/Myo5a-D1 经测序等证实构建成功。而后将重组质粒依次转化至大肠杆菌BL21 感受态细胞,经IPTG诱导重组蛋白表达,后制备了一个小片段的多克隆抗体。因国外有文献报道,许多外源蛋白在BL21中表达时,以一种不溶性的沉淀即包涵体形式存在,这样不利于外源蛋白施展生物学活性。所以本研究分别对菌体沉淀和裂解液中的蛋白质进行SDSPAGE 电泳,最后选取在裂解细菌后的上清中能找到目的蛋白的质粒,这样的质粒才具有可溶的生物学活性。同时电泳显示蛋白相对分子量约为30 kD,与预期相符。Western blot 也同时证实该蛋白具有免疫反应特异性。因此最终经过亲和层析柱对蛋白进行了纯化,得到了PGEX-4T-3/Myo5a-D1融合蛋白的纯品制备多克隆抗体,为后续进一步研究Myo5a 与其他肿瘤的作用提供了铺垫,从而能够继续探讨Myo5a在癌细胞转移中的作用。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
疯狂英语·新悦读(2020年7期)2020-07-30
癌变·畸变·突变(2020年1期)2020-02-12
装备制造技术(2019年12期)2019-12-25
上海建材(2017年4期)2017-04-06
现代检验医学杂志(2016年1期)2016-11-12
上海蔬菜(2015年2期)2015-12-26
安徽医科大学学报(2015年9期)2015-12-16
动物医学进展(2015年10期)2015-12-07
动物医学进展(2015年10期)2015-12-07
