
荔枝果皮细胞壁组成与裂果发生关系的研究
2023-11-13 07:16葛涵涛林诗雅王惠聪
广东农业科学 2023年9期
葛涵涛,林诗雅,王惠聪
(华南农业大学园艺学院,广东 广州 510642)
【研究意义】荔枝(LitchichinensisSonn.)的种植面积在我国果树面积中排第六,属于大宗果树,产业地位重要。然而,一些优质荔枝品种裂果高发是制约荔枝产业发展的重要因素,亟需对裂果发生的内在机制深入研究,以研发减轻荔枝裂果的技术策略,为抗裂育种提供重要参考,促进荔枝产业的高质量发展。【前人研究进展】裂果是一种生理性病害,是果实对内部生长与外界环境不协调而作出的应激反应[1]。果实从膨大期到采收期都可能发生裂果现象,多集中在果实膨大期,但部分果实也可能在采后出现开裂现象[2-4]。裂果的发生主要是由果皮的生理失调导致[5],其破坏了果实的外观,使果实内部水分流失、病原菌入侵,影响果实的采收,从而降低果实的商品性[6]。果实开裂的现象常见于柑橘、荔枝、枣和葡萄等果树中[7-10]。与裂果相关因素的研究大多集中在生理生化方面,关于水分、赤霉素、脱落酸、钙等因素对裂果的影响已有大量研究报道[5]。荔枝果实的发育始于果皮和种皮的发育,荔枝果实生长上著名的“球皮对球胆效应”[11]认为,荔枝果皮在为假种皮提供生长空间的同时又限制了假种皮的生长。当果皮生长接近最大值时,假种皮开始快速生长[12]。大量研究表明,环境因素、矿质营养、植物激素、细胞壁修饰酶、果皮组织结构和遗传因素都可能与荔枝裂果的发生有关[13-15]。【本研究切入点】在裂果高发期,假种皮的突发性生长是裂果的直接原因,但对裂果起关键作用的果皮生长情况已在果实发育的早中期决定了。使用2,4-D 对荔枝处理后可以促使荔枝提前成熟[16],而关于2,4-二氯苯氧乙酸(2,4-D)对荔枝裂果的作用则报道较少。还有研究表明,荔枝裂果与果皮细胞壁的降解代谢关系密切,但有关细胞壁合成代谢方面研究也较少。【拟解决的关键问题】本研究比较裂果易感性不同的荔枝品种细胞壁组分的差异,通过外源浸泡2,4-D,揭示细胞壁组分与裂果发生的关系,初探荔枝裂果发生机制,为荔枝防裂果栽培技术的研发提供重要的参考。
1 材料与方法
1.1 试验材料
于2021 年4 月— 6 月在华南农业大学长岗山荔枝种质资源圃,选取树龄约20 年、树体健壮、长势良好、花期相近的‘怀枝’‘岭丰糯’‘桂味’‘糯米糍’各3 株作为比较试验树,其中‘桂味’和‘糯米糍’属于易裂品种,在果实成熟期每株随机采取状态相似的10 穗果实,带回实验室后,立即将果皮分离并用清水清洗,用纱布吸干后放入液氮,后转入-80 ℃冰箱保存备用。
2021 年在深圳南山西丽果场第二分场和华南农业大学荔枝种质资源圃各选择树体健壮、长势良好且相似的3 株‘糯米糍’作为试验树,进行生长素类似物2,4-D 浸泡试验,并跟踪调查果实落果和开裂情况。
1.2 外源2,4-D 浸泡处理
从每株‘糯米糍’试验树上随机选取长势一致的果穗80 穗进行挂牌,在花后28 d 分别用10、20、40 mg/L 的2,4-D 浸泡,以清水为对照,每个浓度处理20 穗,处理后每周对落果数和裂果数进行调查统计。在果实转色的裂果高发期(花后75 d)将果实采下,将果皮分离并用清水清洗,用纱布吸干后放入液氮,后转入-80 ℃冰箱保存备用。
1.3 荔枝果皮组织结构观察
1.3.1 透射电镜样品制备与观察 在田间用干净的双面刀片将果皮切成长×宽约为0.3 cm×0.2 cm的小块,放入装有4%戊二醛溶液的2 mL 离心管中,并用针管抽气至样品完全沉在管底,密封后置于4 ℃,固定过夜,用0.1 mol/L 磷酸缓冲液(pH=7.0)漂洗4 次,每次20 min。将样品转至1% 锇酸溶液中固定2~4 h,ddH2O 漂洗4 次,每次20 min。用1% 醋酸双氧铀块染色,4 ℃ 过夜,ddH2O 漂洗4 次,每次20 min。依次用30、50、70、85、95%的乙醇进行脱水,每次15 min,再用100%的乙醇脱水2 次,每次20 min。将样品转至纯丙酮中浸泡2 次,每次20 min。依次用3∶1、1∶1、1∶3 的丙酮:树脂渗透,每次2~8 h,再用纯树脂渗透2 次,每次12 h。将渗透后的样品转至包埋板上用树脂包埋,置于常温干燥器,3 h后放入烘箱,70 ℃ 聚合24~48 h。
使用Leica UC7 冷冻超薄切片机(德国徕卡公司)对包埋好的样品进行切片,厚度为70~90 nm,用醋酸双氧铀溶液和柠檬酸铅溶液对切片分别染色30 min。在Talos L120C 型透射电镜(美国FEI公司)中观察切片中果皮的组织结构,并测量不同部位细胞壁的厚度。
1.3.2 果皮组织木质素染色与分布观察 参照Yi等[17]的方法进行。
1.4 碳水化合物的提取和含量测定
参照Yi 等[17]的方法测定荔枝果皮中非结构性碳水化合物(白坚木皮醇、果糖、葡萄糖、半乳糖、蔗糖、淀粉)、结构性碳水化合物(果胶、纤维素和半纤维素)和木质素的含量,所测定的值均为鲜重含量。
2 结果与分析
2.1 不同裂果易感性荔枝品种果皮碳水化合物含量比较
2.1.1 果皮非结构性碳水化合物含量 花后75 d是各品种果实开始转色的时期,也是裂果高发期。比较裂果高发期4 个品种果皮中非结构碳水化合物的含量(图1)发现,易裂荔枝品种‘桂味’和‘糯米糍’的白坚木皮醇含量分别为9.8、8.5 mg/g,显著高于‘怀枝’的7.5 mg/g;而‘桂味’和‘糯米糍’的果糖含量(20.4、22.6 mg/g)则显著低于‘怀枝’(32.3 mg/g)。‘桂味’中葡萄糖与半乳糖含量显著低于‘怀枝’中的葡萄糖与半乳糖含量。‘糯米糍’中蔗糖和淀粉含量也显著低于‘怀枝’的蔗糖和淀粉含量。
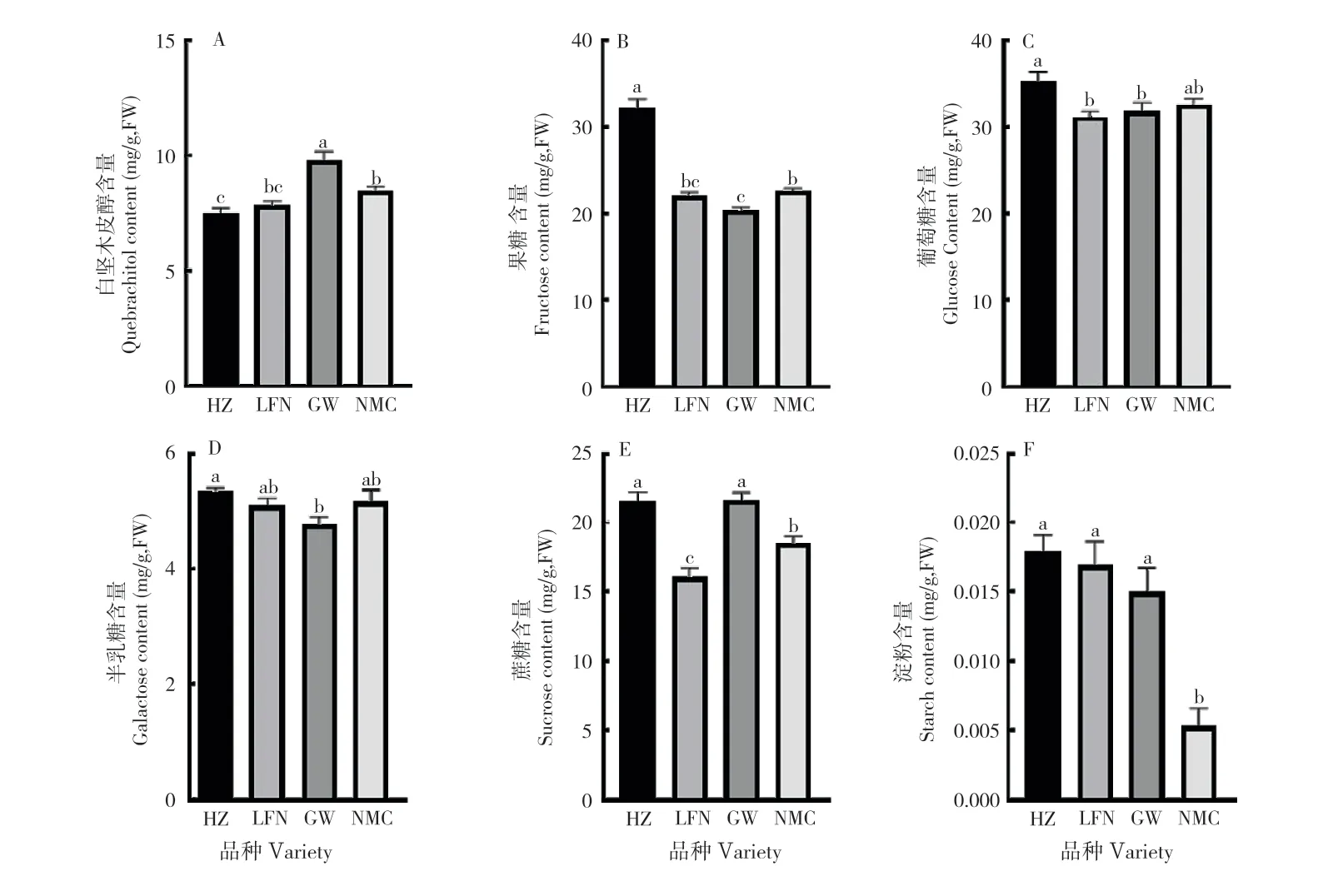
图1 不同荔枝品种果皮非结构性碳水化合物含量比较Fig.1 Comparison of non-structural carbohydrate contents in the pericarp of different litchi varieties
2.1.2 果皮结构性碳水化合物和木质素含量 比较裂果高发期4 个品种果皮中结构性碳水化合物和木质素含量(图2A),发现‘怀枝’果皮中的果胶含量(7.8 mg/g)显著高于‘岭丰糯’(1.5 mg/g)和‘糯米糍’(1.1 mg/g),‘桂味’果皮中果胶含量(4.3 mg/g)也显著高于‘岭丰糯’和‘糯米糍’,其中‘糯米糍’果皮中果胶含量最低。‘怀枝’‘岭丰糯’‘桂味’‘糯米糍’的半纤维素和纤维素含量分别为12.7、11.8、17.8、10.3 mg/g,其中‘桂味’果皮中的半纤维素和纤维素含量显著高于其他3 个品种,‘糯米糍’果皮中的半纤维素和纤维素含量最低,显著低于‘怀枝’。‘桂味’果皮中的木质素含量(9.7 mg/g)也显著高于‘怀枝’(6.6 mg/g)和‘糯米糍’(6.7 mg/g)。
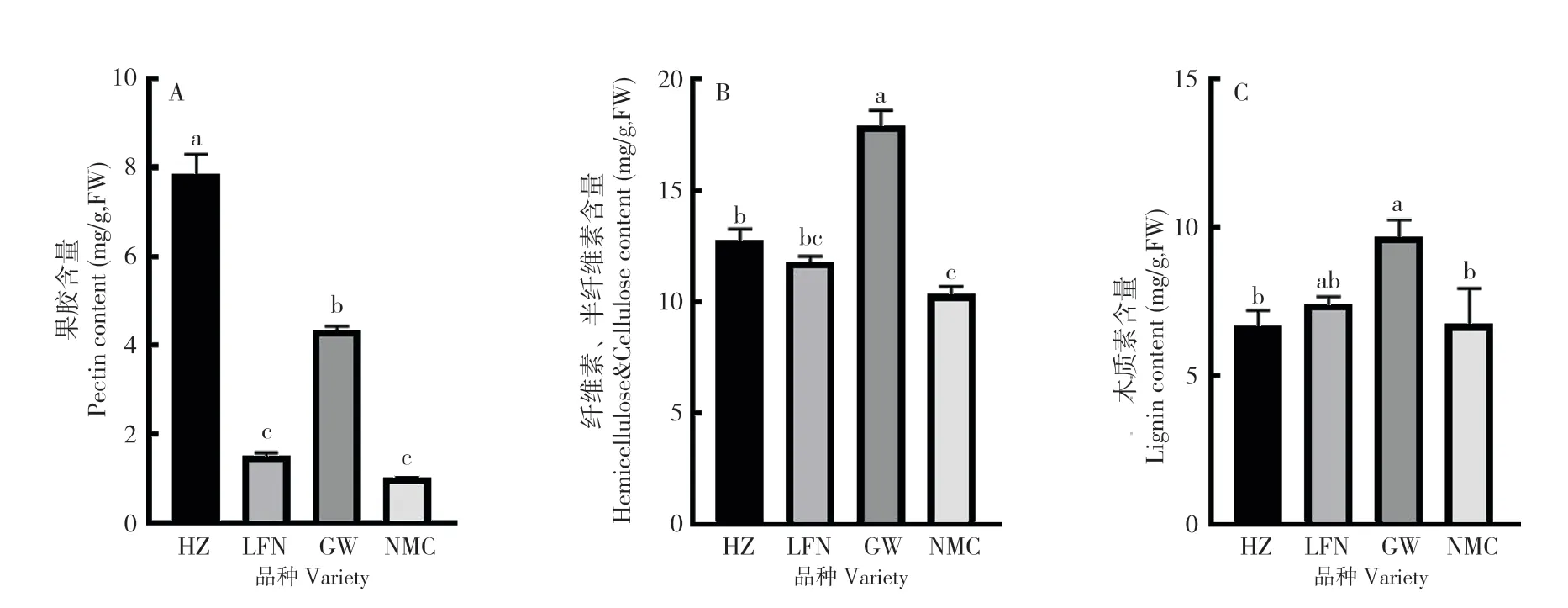
图2 不同的荔枝品种果皮结构性碳水化合物和木质素含量比较Fig.2 Comparison of structural carbohydrate and lignin contents in the pericarp of different litchi varieties
2.1.3 果皮组织结构和木质素分布 为更加直观地观察不同品种荔枝果皮组织结构和木质素的分布,用间苯三酚-盐酸对4 个品种花后60~85 d的果皮进行染色观察。发现(图3)随着果实的发育,花后70~75 d,4 个品种的果皮都显著变薄,这与裂果高发期的时间相符,花后80 d 后,‘岭丰糯’和‘糯米糍’的果皮继续变薄,而‘怀枝’和‘桂味’则变化不大。另外,花后75 d 后,‘怀枝’的果皮整体被染得较红,而‘糯米糍’的果皮中则出现略微变褐的情况,表明该时期‘怀枝’果皮中的木质素含量高于‘糯米糍’。

图3 不同荔枝品种果皮组织结构和木质素分布Fig.3 Tissue structure and lignin distribution in the pericarp of different litchi varieties
2.2 2,4-D 处理对‘糯米糍’荔枝落果率和裂果率的影响
2.2.1 落果率 在花后28 d 用10、20、40 mg/L的2,4-D处理‘糯米糍’的果穗,在西丽果场(图4),处理后7 d,3 个处理的落果率分别为26.5%、32.2%、13.9%,处理后14 d,3 个处理的落果率分别为52.8%、57.3%、33.9%,均极显著低于对照的52.8%和64.5%;处理后25 d,20 mg/L 和40 mg/L 处理落果率分别为64.5%、70.6%、仍显著低于对照(78.2%),但10 mg/L 处理的落果率(76.3%)已与对照无显著差别;处理后42 d,20 mg/L 处理累积落果率(87.4%)甚至显著高于对照(80.6%);处理后52 d,处理与对照间的落果率无显著差异。在华农果场的处理中(图4),处理后7 d,3 个处理的落果率分别为86.3%、85.9%、86.4%,均极显著低于对照(87.9%);处理后14 d,20 mg/L 和40 mg/L 处理落果率为93.1%和93.2%、仍极显著低于对照(96.4%),但10 mg/L的处理与对照无显著差异;处理后21 d,处理与对照间的落果率无明显差异。这些结果表明,2,4-D 在果实发育前期可保果,且浓度较大保果效果较好,但2,4-D 只是延迟果实的脱落,对最终的座果率无显著的影响。
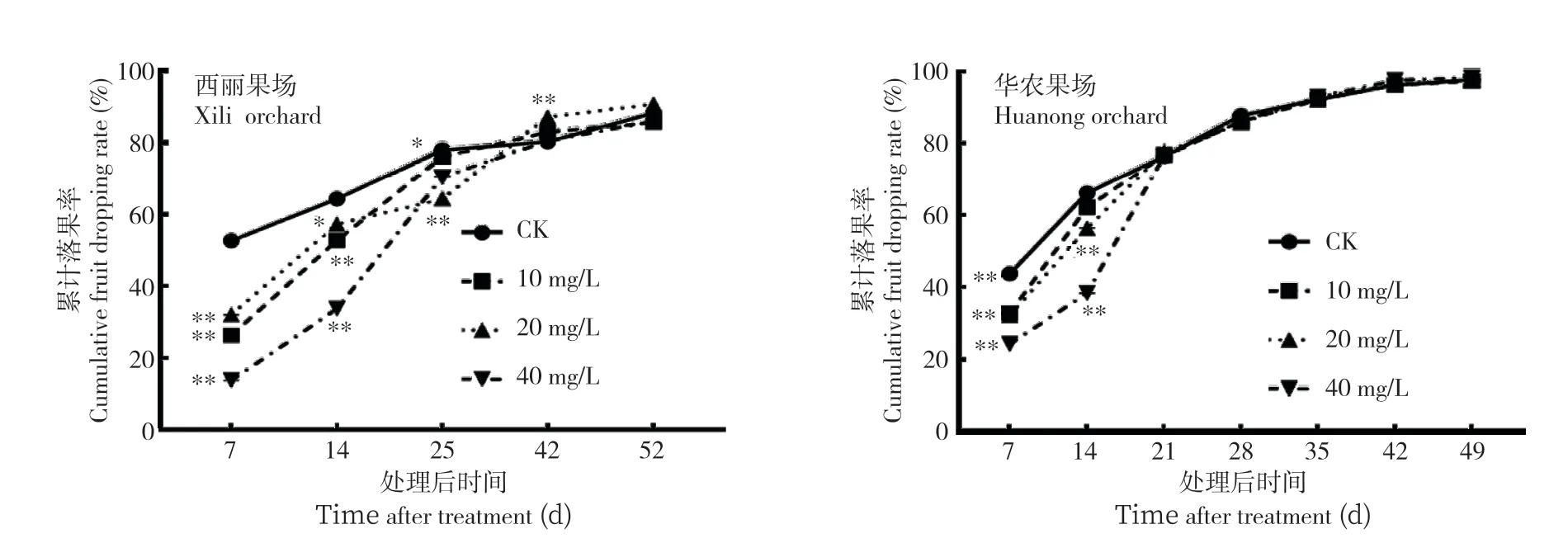
图4 2,4-D 处理对‘糯米糍’荔枝落果率的影响Fig.4 Effect of 2,4-D treatment on fruit dropping rate of ‘Nuomici’ litchi
2.2.2 裂果率 对裂果率进行调查时发现(图5),在西丽果场的处理中,10 mg/L 处理后的果实没有出现开裂情况,20、40 mg/L 处理后的果实裂果率分别为2.78%和3.05%,均高于对照(1.08%),但由于果穗之间的差异较大,处理与对照之间裂果率差异均未达到显著水平。在华农果场的处理中,3 个不同浓度的2,4-D 处理均增加了裂果率,且随着2,4-D 浓度增加裂果率升高,40 mg/L 2,4-D处理后的累积裂果率达10.75%,显著高于对照的1.9%。上述结果表明,高浓度的2,4-D 处理显著增加裂果的发生。
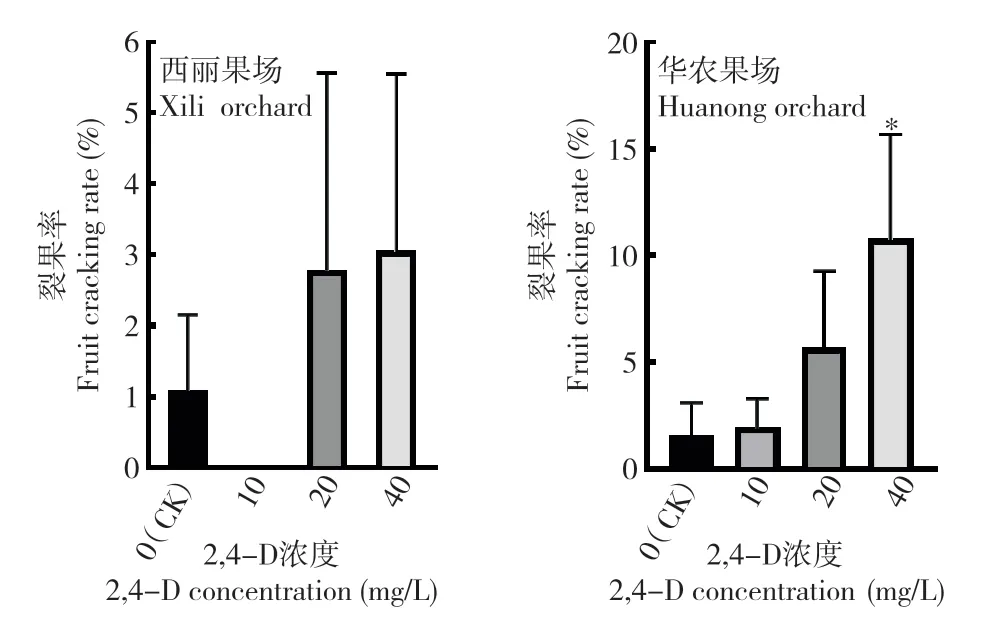
图5 2,4-D 处理对‘糯米糍’荔枝裂果率的影响Fig.5 Effect of 2,4-D treatment on fruit cracking rate of ‘Nuomici’ litchi
2.2.3 2,4-D 处理对果皮组织结构的影响 2,4-D处理后果实开裂时间大大提早,华南农业大学荔枝园40 mg/L 处理的果实,在处理后42 d 就开始出现裂果。本研究取处理后42 d 的样品进行透射电镜观察发现(图6),2,4-D 处理后,果皮细胞层数增多,内果皮细胞轮廓变模糊,内中果皮细胞较大且大小不均。果皮组织结构变化程度与处理浓度呈正相关,说明果皮细胞层数、内果皮细胞轮廓清晰度和内中果皮细胞大小与果皮应力相关,进而影响了裂果。另外,外中果皮的薄壁细胞明显变小、数量增多,40 mg/L 2,4-D 处理的外中果皮的细胞壁明显增厚。

图6 2,4-D 处理对‘糯米糍’荔枝果皮组织结构的影响Fig.6 Effect of 2,4-D treatment on the tissue structure in pericarp of ‘ Nuomici’ litchi
2.2.4 2,4-D 处理对果皮细胞壁组分的影响进一步测定2,4-D 处理后的每克鲜质量果皮中细胞壁组分的含量,结果如图7 所示,随着果实发育,处理组中果皮果胶含量逐渐下降,对照组中的果胶含量则逐渐上升。在处理后7 d 3 个浓度2,4-D处理的果胶含量分别为6.0、8.9、12.1 mg/g,对照组中的果胶含量仅为2.7 mg/g,表明2,4-D 处理后显著增加果皮果胶含量,且随着处理浓度增加,增加幅度加大。在处理后42 d 除40 mg/L 的果胶含量(10.3 mg/g)仍显著高于对照(5.9 mg/g)外,两个低浓度处理果皮中的果胶(4.8、6.7 mg/g)均与对照无显著差异。到花后52 d,随着对照样品果胶的增加和处理样品果胶的减少,两个低浓度处理果胶含量(2.2、4.6 mg/g)均显著低于对照(7.7 mg/g),而裂果最严重的40 mg/L处理中果皮的果胶(8.9 mg/g)与对照无显著差异。不同浓度2,4-D 处理对荔枝果皮半纤维素和纤维素的含量(图7B)影响较小,高浓度处理在不同的阶段似乎会降低半纤维素和纤维素的水平,但未达到显著。处理对木质素含量(图7C)的影响也不明显,在处理后7 d,20、40 mg/L 处理的木质素含量(8.6、9.4 mg/g)均明显低于对照和10 mg/L(10.7、11.3 mg/g)的处理果皮,在其余发育阶段,对照和处理之间木质素的含量无显著差异。
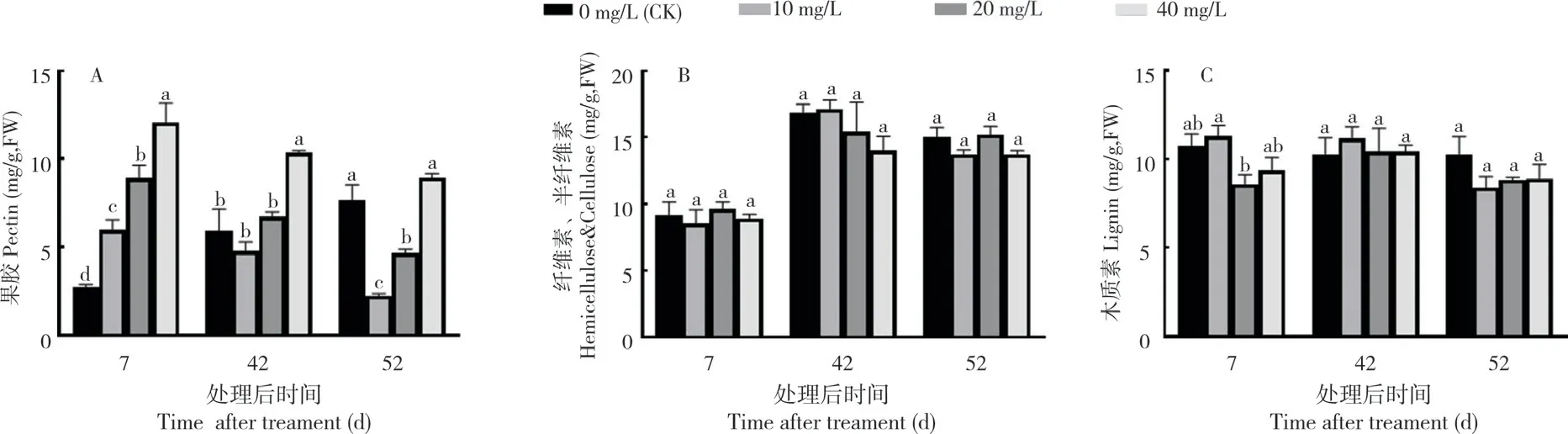
图7 2,4-D 处理对果皮细胞壁组分的影响Fig.7 Effect of 2,4-D treatment on the components of pericarp cell wall
3 讨论
3.1 不同品种荔枝果皮碳水化合物与细胞壁代谢和裂果易感性的关系
碳水化合物是植物光合作用的主要产物,也是植物生长发育过程的重要营养因子。叶片是光合作用的主要场所,光合作用的同化产物从叶片(源器官)运输到果实(库器官)后被分解成己糖,磷酸戊糖途径和糖酵解途径进一步将己糖代谢,为植物提供生长发育所需的能量及代谢中间产物,同时也为植物提供细胞碳架、调控相关基因的表达与酶活性,从而影响植物的生长发育[18-20]。己糖包含果糖、葡萄糖和半乳糖等,其中葡萄糖是果胶与半纤维素和纤维素的重要合成前体,而半乳糖是组成果胶的重要单体。有研究发现,在果胶降解的过程中,果胶的合成前体半乳糖醛酸、葡萄糖和阿拉伯糖等含量也会减少[21-24],说明己糖与细胞壁之间有着密不可分的关系。本研究对裂果易感性不同的荔枝品种果皮中非结构性碳水化合物的变化趋势与含量进行分析发现,裂果率低的‘怀枝’3 种己糖的含量明显高于易裂果的‘糯米糍’和‘桂味’。相对于己糖,白坚木皮醇和蔗糖的含量与裂果没有明显规律性的联系,表明果皮中的己糖水平与裂果的发生有重要联系,可能在果皮的细胞壁代谢和发育中发挥重要作用[17],影响果皮细胞壁的强度与完整性,进而影响裂果的易感性。
细胞壁结构性多糖组成的变化可能通过影响细胞壁的完整性与机械性来影响果皮的结构与状态,进而影响裂果的发生[25]。原果胶在细胞壁代谢过程中被降解成可溶性果胶,导致细胞壁胞间层与初生壁分解,细胞间黏合力下降,破坏细胞壁结构[26]。果皮中原果胶与纤维素的减少,会引起细胞壁总体结构的破坏,降低果皮的硬度与应力,增加果实的开裂[27-29]。本研究中,‘怀枝’的果胶含量明显高于其他3 个品种,易裂果的‘糯米糍’果皮中的果胶基本上均低于其余3 个品种。半纤维素和纤维构成了细胞壁的基本骨架,两者连接形成网状结构保持细胞壁的稳定性[24],同时纤维素微纤丝的排列方式也决定了细胞壁的致密性[30],果皮纤维素含量较高或纤维素酶活性较低的果实一般开裂率较低[12]。本研究中,易裂果的‘糯米糍’果皮半纤维素和纤维素含量均明显低于其他品种。次生壁中木质素的沉积造成木质化,提高了细胞壁的强度[31]。观察果皮组织结构和木质素的分布可发现,‘糯米糍’果皮纤维中的木质素一直比‘怀枝’少,然而‘桂味’中却比‘怀枝’中多,表明裂果高发期果皮中果胶含量可能更显著影响裂果的发生。此外,在易裂品种‘糯米糍’与‘桂味’中的细胞壁组成成分含量与代谢可能存在差异(特别是半纤维素与纤维素和木质素),不同品种诱发果实开裂的具体原因可能存在差异。
3.2 2,4-D 处理对‘糯米糍’荔枝细胞壁代谢和裂果率的影响
2,4-D是生产上常用的一种保果生长调节剂,在荔枝上使用的浓度一般为10 mg/L,一些生产者会提高2,4-D 的浓度,以求达到更好的保果效果。本研究设置了10、20、40 mg/L 浓度的2,4-D 对‘糯米糍’进行处理,发现在果实发育前期,2,4-D处理可显著降低落果率,但在果实发育后期,落果率与对照没有显著差异,且随着使用浓度的增加裂果率明显上升。上述结果说明,2,4-D 可以减少荔枝落果是果实发育前期的表象,后期反而会增加裂果的风险。
果皮组织结构的差异与完整性程度是裂果发生的重要影响因素。表皮细胞的厚度、细胞壁的厚度和角质层的厚度都是影响裂果的因素[7,32]。本研究中,2,4-D 处理在增加裂果率的同时,也加厚了果皮的厚壁组织、外中果皮和内果皮,说明2,4-D 可能诱导了细胞壁厚度的增加,而果皮细胞壁的增厚可能限制细胞的膨大,降低了果皮的延展性从而增加了开裂的发生。果皮的机械性是保证果实完整性的重要性质,果皮组织结构的差异与裂果息息相关[33-35]。易裂的荔枝果实外果皮细胞较细长,而内果皮的细胞层数较多且细胞的轮廓不清晰,果实开裂处果皮组织的排列连接变得模糊不清[36-38]。本研究对果皮进行透射电镜观察,发现2,4-D 处理后果皮细胞层数增多,内果皮的细胞轮廓变模糊,内中果皮细胞较大且大小不均,同时外中果皮的薄壁细胞明显变小、数目增多,果皮组织结构的变化和差异与前人研究结果一致,说明果皮的细胞层数和中果皮的细胞大小及均匀程度会影响果实的开裂。比较细胞壁组分含量与变化,发现在裂果高发期时,在2,4-D 诱导下的裂果果皮中半纤维素和纤维素含量比对照低但差异不显著;处理的果皮果胶含量显著高于对照,但发育过程中的果胶含量变化动态有明显区别,对照明显增加,而处理明显下降。处理显著增加果皮果胶的含量可能与细胞壁明显增厚有关,植物细胞的生长涉及细胞壁组成的降解和重新合成,在生长发育过程中细胞壁代谢是一个动态的过程,仅测定细胞壁的组分可能很难判定2,4-D 如何影响细胞壁代谢,进一步的研究需要探讨处理后细胞壁代谢相关基因表达和酶活性的变化。
4 结论
裂果易感性不同的荔枝品种果皮中非结构性碳水化合物与结构性碳水化合物含量存在明显差异,其中果皮中己糖和果胶的含量与裂果的发生密切相关。易裂果的‘糯米糍’果皮中的果糖(22.6 mg/g)和蔗糖(18.6 mg/g)含量明显低于不易裂果的‘怀枝’(32.3、21.5 mg/g);果胶与裂果的关系较为复杂,本研究的4个品种中,易裂的‘糯米糍’果皮果胶含量(1.1 mg/g)低于不易裂果的‘怀枝’(7.8 mg/g)和‘岭丰糯’(1.5 mg/g),而同是易裂果的‘桂味’果皮果胶含量(4.3 mg/g)则高于‘怀枝’和‘岭丰糯’,说明果胶含量过低或过高都可能导致裂果高发。2,4-D 处理后果皮果胶增多,裂果率增加,进一步表明高含量果胶会增加裂果风险。
猜你喜欢
今日农业(2022年15期)2022-09-20
今日农业(2022年2期)2022-06-01
河北果树(2022年1期)2022-02-16
今日农业(2021年11期)2021-11-27
河北果树(2020年4期)2020-11-26
中国化肥信息(2018年11期)2018-12-06
浙江工业大学学报(2017年5期)2018-01-22
广东农业科学(2017年5期)2017-08-29
时代农机(2016年6期)2016-12-01
塔里木大学学报(2014年3期)2014-03-11
