
围产期饲粮中添加异位酸对奶牛生产性能、营养物质表观消化率、瘤胃发酵参数和奶牛、新生犊牛血清指标的影响
2023-11-10 06:09马小雪聂存喜罗瑞卿吴妍妍张文举
动物营养学报 2023年10期
龙 锐 马小雪 聂存喜 罗瑞卿 吴妍妍* 张文举*
(1.石河子大学动物科技学院,石河子 832000;2.新疆天山军垦牧业有限责任公司,石河子 832000)
奶牛围产期又叫“过渡期”,一般是从奶牛产前21 d(干奶的晚期)到产后21 d(泌乳的初期),是奶牛整个泌乳期中最重要的一个阶段[1]。围产期奶牛物质代谢发生剧烈改变,免疫力降低、抗病能力下降、食欲减退,机体处于应激状态[2]。围产期奶牛对营养物质需求急剧增加,但由于胎儿生长迅速,体积增大,导致奶牛瘤胃空间受挤压而变小,加上基础饲粮组成的改变,致使围产期奶牛干物质(DM)采食量、生理代谢功能下降严重[3-4],当奶牛免疫机能紊乱时,代谢疾病和炎症疾病发病率显著提高[5]。由于围产期和泌乳前期的奶牛极易出现能量负平衡(NEB)状态,机体只能通过体脂动员来满足能量需要,导致脂质代谢增强[6];自由基在体内大量积累,易产生氧化应激,进而降低生产性能,威胁奶牛健康[7]。
异位酸属于短链挥发性脂肪酸,主要包括异戊酸、异丁酸和2-甲基丁酸[8]。瘤胃中的异位酸是由蛋白质降解后的支链氨基酸(缬氨酸、亮氨酸和异亮氨酸)经过氧化脱氨基得到,也是纤维分解菌生长繁殖所需要的营养因子[9]。异位酸是反刍动物瘤胃中蛋白质生产必不可少的重要中间体,具有促进瘤胃纤维降解菌生长繁殖[10]、调节瘤胃发酵、促进微生物蛋白(MCP)合成、提高氨氮利用率[11-12]、抗热应激、改善NEB等作用[13]。异位酸可增加瘤胃内乙酸的合成,而乙酸对于动物机体体脂的合成至关重要[14]。饲粮中添加异丁酸或异戊酸能提高瘤胃中乙酸产量、纤维分解菌数量、纤维分解酶活性[15-16]和饲粮养分消化率[17];还能提高瘤胃中MCP合成量和饲粮蛋白质利用率[18]。王加启等[13]发现,异位酸型添加剂可使奶牛产奶量增加15.4%。Liu等[19]在泌乳牛饲粮中添加异丁酸,机体血清葡萄糖(GLU)含量上升。刘晏榕等[20]研究表明,犊牛饲粮中添加异位酸,能显著提高血清中GLU和白蛋白(ALB)含量。张振威等[21]发现,异丁酸能增加西门塔尔牛的饲料转化率和日增重。冯蕾[22]研究表明,热应激会对荷斯坦奶牛的抗氧化能力产生不利影响。但有研究表明,添加混合异位酸对热应激有缓解作用[23]。此外,封丽梅[24]研究表明,饲粮中添加戊酸可增强奶牛的免疫能力。因此,在围产期饲粮中添加异位酸可提高动物机体蛋白质含量、生产性能、产奶量、血清GLU含量、抗氧化及免疫能力。但目前关于异位酸对围产期奶牛及其通过母体效应对新生犊牛的相关影响研究较少。因此,本试验旨在研究饲粮中添加异位酸对围产期奶牛生产性能、营养物质表观消化率、瘤胃发酵参数和奶牛、新生犊牛血清指标的影响,确定其适宜添加量,为在围产期奶牛生产中应用异位酸添加剂提供科学依据。
1 材料与方法
1.1 试验设计
本试验于2022年7—10月在新疆石河子市西锦牧业有限责任公司奶牛场进行。选择18头健康、第3胎次、预产期相近的经产荷斯坦围产期奶牛,按照完全随机区组设计分为3组,每组6个重复,每个重复1头奶牛。对照组饲喂基础饲粮,Ⅰ组饲喂基础饲粮+30 mL/(d·头)的异位酸,Ⅱ组饲喂基础饲粮+60 mL/(d·头)的异位酸。预试期10 d,正试期42 d。
1.2 试验饲粮及饲养管理
试验各组围产前期(产前21 d~分娩)和围产后期(分娩~产后21 d)饲喂同样的基础饲粮,其组成及营养水平见表1。每天等量饲喂3次(09:00、15:00和21:00),牛只全天自由饮水,试验牛每次晨饲前将异位酸喷洒在部分精料中混匀,每头奶牛单独采食,确保试验牛全部采食后,再进行正常饲喂。试验所用异位酸由异丁酸和异戊酸按1∶1混合而成,由郑州某化工公司提供,纯度均>99%。
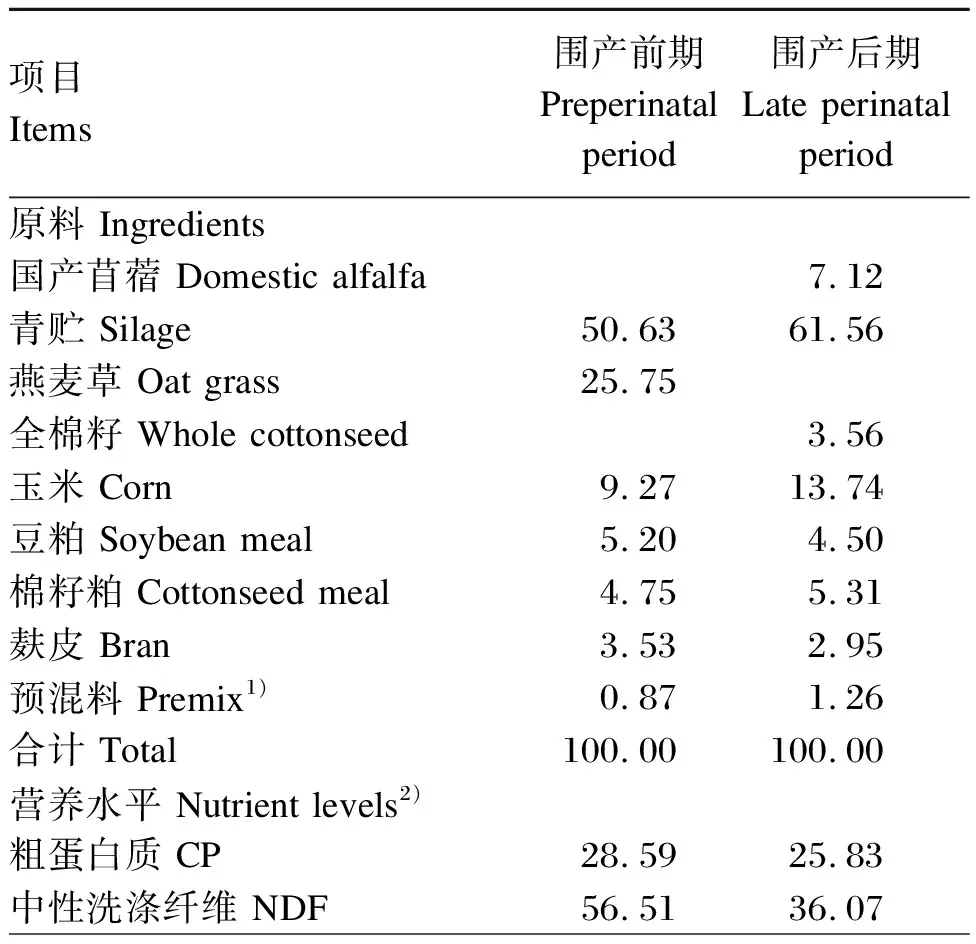
表1 基础饲粮组成及营养水平(干物质基础)
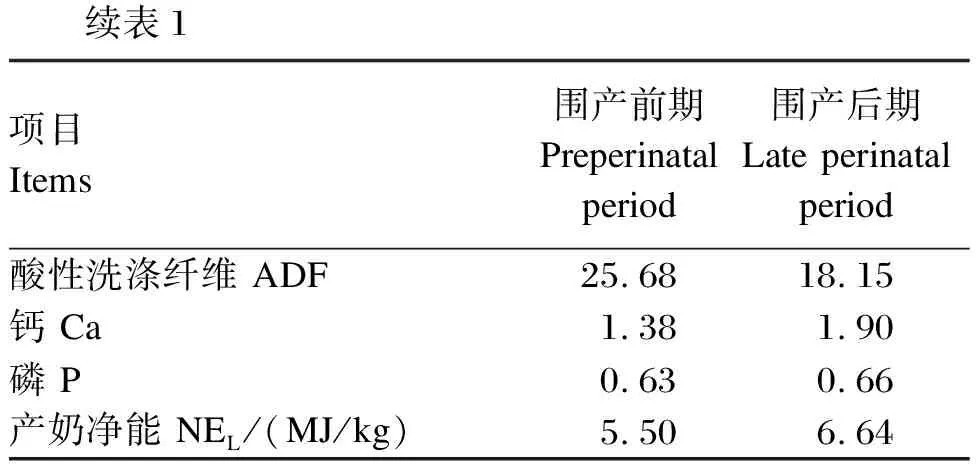
续表1项目 Items围产前期 Preperinatal period围产后期 Late perinatal period酸性洗涤纤维 ADF25.6818.15钙 Ca1.381.90磷 P0.630.66产奶净能 NEL/(MJ/kg)5.506.64
1.3 样品采集与测定
1.3.1 产奶量
在奶牛产后9~10 d和产后20~21 d连续2 d记录早、中、晚3次产奶量,取2 d产奶量的平均值记为1次产奶量。
1.3.2 初生重和体尺
犊牛出生后喂初乳之前用电子磅秤测定初生重;待犊牛站立较稳时用卷尺和测杖测定相关体尺指标(体高、体斜长、胸围、管围)。
1.3.3 营养物质表观消化率
试验期最后3 d,每天早、中、晚通过直肠收集粪便,随机取样法收集饲粮样品。饲粮、剩料及粪便样品在65 ℃下用鼓风干燥48 h后将其碾碎制成风干样。参考张丽英[25]所描述的方法对样品中的DM、粗蛋白质(CP)、中性洗涤纤维(NDF)和酸性洗涤纤维(ADF)含量进行测定。以粪便和饲粮中的酸不溶灰分(AIA)作为内源性指示剂[26],计算营养物质表观消化率,计算公式如下:
某营养物质表观消化率(%)=[1-(饲粮中
AIA含量×粪便中该营养物质含量)/
(粪便中AIA含量×饲粮中该
营养物质含量)]×100。
1.3.4 瘤胃发酵参数
在奶牛产后21 d晨饲前,对试验组所有奶牛采用瘤胃收集液导管装置采集约100 mL瘤胃液,立即使用便携式pH计(力辰科技笔式酸度计)测定瘤胃液pH。将瘤胃液用4层无菌棉纱布过滤后收集滤液,将其分装于15 mL无菌离心管中,做好标记,置于-20 ℃冰柜保存,用于测定氨态氮(NH3-N)、MCP和挥发性脂肪酸(VFA)含量。
NH3-N含量采用冯宗慈[27]比色法测定;MCP含量采用试剂盒(TransGen Biotech Co,中国),并配合全波长酶标仪(Multiskan GO,美国)测定,测定方法按照试剂盒说明书进行;乙酸、丙酸和丁酸含量采用顶空-气相色谱仪(Agilent GC7890B,美国)测定[28]。
1.3.5 血清相关指标
在奶牛产前10 d、分娩当天、产后10 d和产后21 d晨饲前,每头奶牛经尾静脉采集血样15 mL;在试验各组新生犊牛饲喂其母系初乳12 h后,经颈静脉采集血样10 mL。4 000 r/min离心10 min,分离血清,用2 mL无菌尖头吸管吸取血清,分装于1.5 mL的尖底无菌离心管中,分别做好标记,于-20 ℃储存。
1.3.5.1 奶牛血清生化指标
送石河子大学第一附属医院使用全自动生化分析仪测定奶牛血清中ALB、球蛋白(GLB)、尿素氮(UN)、甘油三酯(TG)、总胆固醇(TC)、GLU、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)含量及谷丙转氨酶(ALT)和谷草转氨酶(AST)活性。
1.3.5.2 奶牛和新生犊牛血清抗氧化指标
分别测定奶牛和新生犊牛血清中谷胱甘肽过氧化物酶(GSH-Px)、超氧化物歧化酶(SOD)活性,丙二醛(MDA)含量和总抗氧化能力(T-AOC)。GSH-Px、MDA和T-AOC试剂盒购自南京建成生物工程研究所,SOD试剂盒购于北京盒子生工科技有限公司,样品处理和测定按照相关试剂盒说明书进行操作。所用仪器为全波长酶标仪(Thermo,美国)。
1.3.5.3 奶牛和新生犊牛血清免疫指标
分别测定奶牛和新生犊牛血清中免疫球蛋白(Ig)A、IgG和IgM含量,由上海酶联生物科技有限公司进行测定。
1.4 数据统计分析
采用SPSS 20.0软件对试验数据进行单因素方差分析(one-way ANOVA),组间差异性比较采用最小显著性差异法(LSD),以P<0.05表示差异显著,P>0.05表示差异不显著。试验结果以平均值±标准差表示。
2 结果与分析
2.1 围产期饲粮中添加异位酸对奶牛生产性能的影响
2.1.1 围产期饲粮中添加异位酸对奶牛产奶量的影响
由表2可知,在产后10和21 d,Ⅰ组和Ⅱ组奶牛产奶量均显著高于对照组(P<0.05)。添加异位酸可促进产后0~21 d奶牛产奶量的增长。

表2 围产期饲粮中添加异位酸对奶牛产奶量的影响
2.1.2 围产期饲粮中添加异位酸对新生犊牛初生重和体尺的影响
由表3可知,Ⅰ组和Ⅱ组新生犊牛的初生重、体高、体斜长、胸围和管围均高于对照组,但差异不显著(P>0.05)。犊牛初始体重Ⅰ组为41.50 kg,Ⅱ组为42.67 kg,分别较对照组提高6.88%和9.98%。
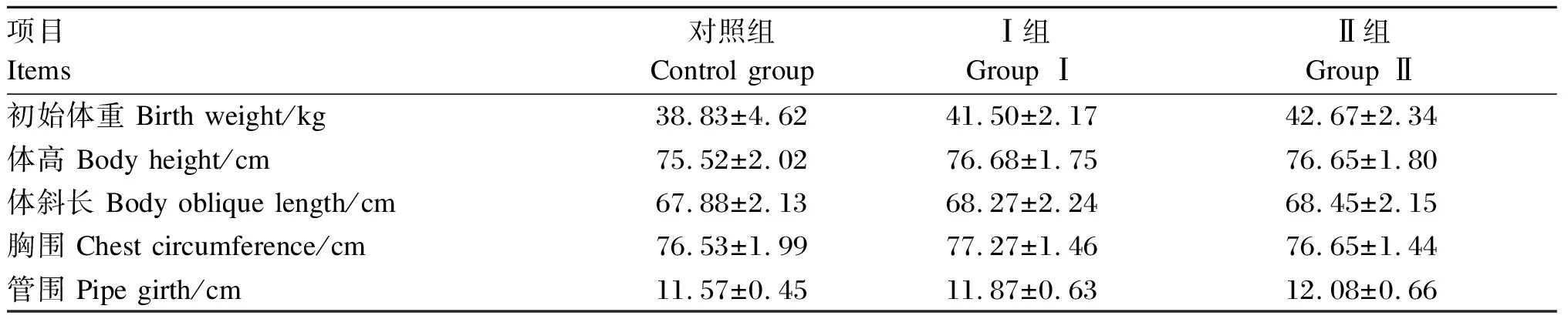
表3 围产期饲粮中添加异位酸对新生犊牛初生重和体尺的影响
2.2 围产期饲粮中添加异位酸对奶牛营养物质表观消化率和瘤胃发酵参数的影响
2.2.1 围产期饲粮中添加异位酸对奶牛营养物质表观消化率的影响
由表4可知,Ⅰ组和Ⅱ组奶牛DM、CP、NDF和ADF表观消化率均显著高于对照组(P<0.05)。
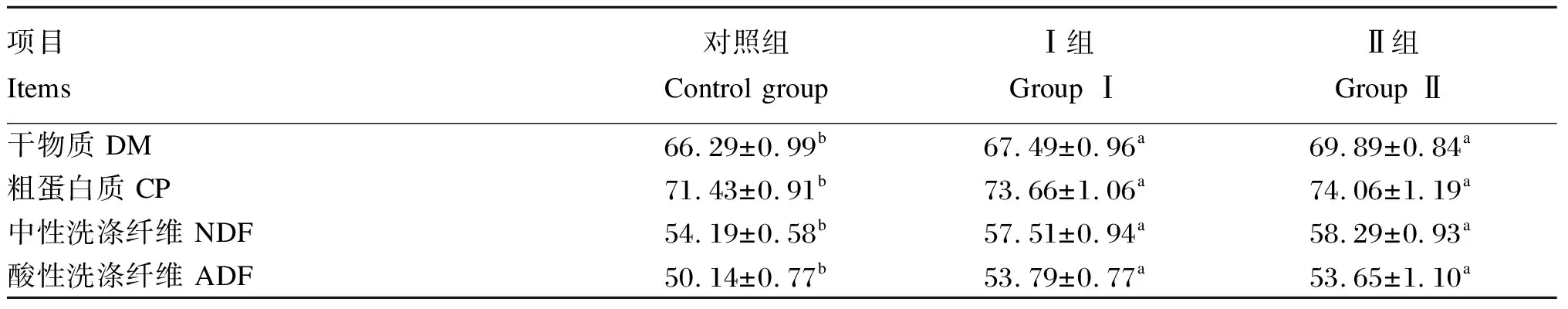
表4 围产期饲粮中添加异位酸对奶牛营养物质表观消化率的影响
2.2.2 围产期饲粮中添加异位酸对奶牛瘤胃发酵参数的影响
由表5可知,Ⅰ组和Ⅱ组瘤胃NH3-N含量均显著低于对照组(P<0.05);Ⅰ组和Ⅱ组瘤胃MCP含量均显著高于对照组(P<0.05);Ⅱ组瘤胃乙酸和总挥发性脂肪酸(TVFA)含量均显著高于对照组(P<0.05);各组间瘤胃pH、丙酸、丁酸含量均无显著差异(P>0.05)。
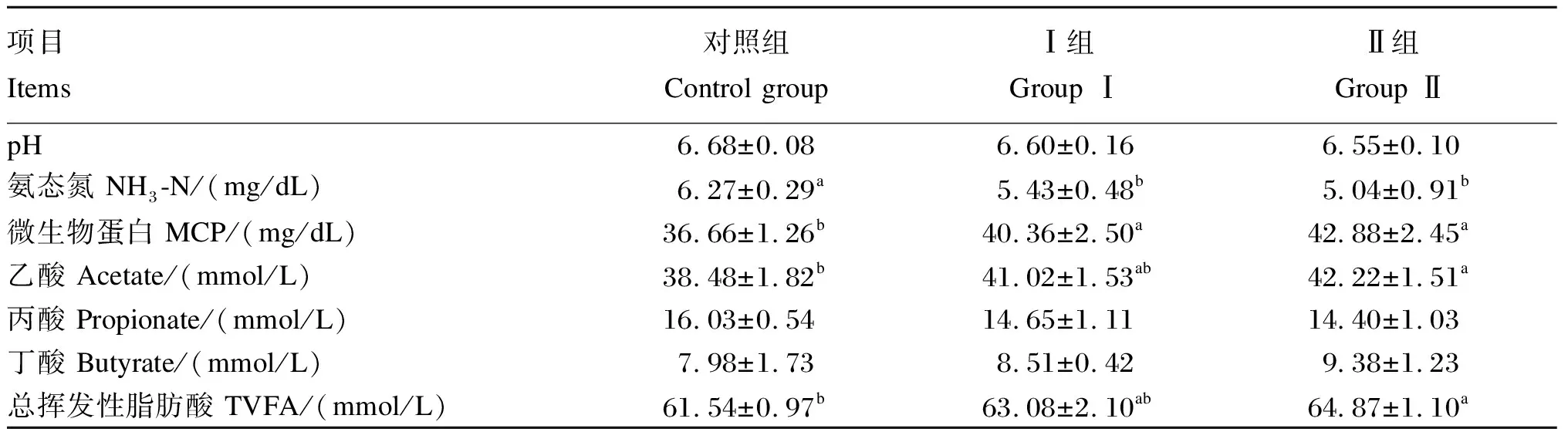
表5 围产期饲粮中添加异位酸对奶牛瘤胃发酵参数的影响
2.3 围产期饲粮中添加异位酸对奶牛血清生化指标的影响
由表6可知,在产前10 d和分娩当天,Ⅱ组血清中ALB和GLB含量均显著高于对照组(P<0.05);在产后10 d,Ⅰ组血清中GLB含量显著高于对照组(P<0.05)。在产前10 d,Ⅰ组和Ⅱ组血清中UN含量均显著低于对照组(P<0.05),分别降低8.94%和10.15%;在产后21 d,Ⅱ组血清UN含量显著低于对照组(P<0.05),降低28.75%。在分娩当天,Ⅱ组血清TG含量显著高于Ⅰ组(P<0.05),但与对照组无显著差异(P>0.05)。在产后21 d,Ⅰ组和Ⅱ组血清中GLU含量均显著高于对照组(P<0.05),分别提高11.35%和15.25%。在产前10 d,Ⅰ组血清中ALT活性显著高于对照组(P<0.05)。不同试验组间血清中TC、HDL-C、LDL-C含量和AST活性均无显著差异(P>0.05)。

表6 围产期饲粮中添加异位酸对奶牛血清生化指标的影响
2.4 围产期饲粮中添加异位酸对奶牛血清抗氧化能力的影响
由表7可知,在产前10 d,Ⅰ组血清中GSH-Px活性显著高于对照组(P<0.05);在产后10和21 d,Ⅱ组血清中GSH-Px活性显著高于对照组(P<0.05)。在分娩当天,Ⅰ组和Ⅱ组血清中MDA含量均显著低于对照组(P<0.05),分别降低6.07%和7.5%;在产后10 d,Ⅱ组血清中MDA含量显著低于对照组(P<0.05),降低15%。在分娩当天,Ⅰ组血清中T-AOC显著高于对照组(P<0.05),提高8.92%;在产后10 d,Ⅱ组血清中T-AOC显著高于对照组(P<0.05),提高13.7%。Ⅰ组和Ⅱ组血清中SOD活性均较对照组有上升趋势,但无显著差异(P>0.05)。

表7 围产期饲粮中添加异位酸对奶牛血清抗氧化能力的影响
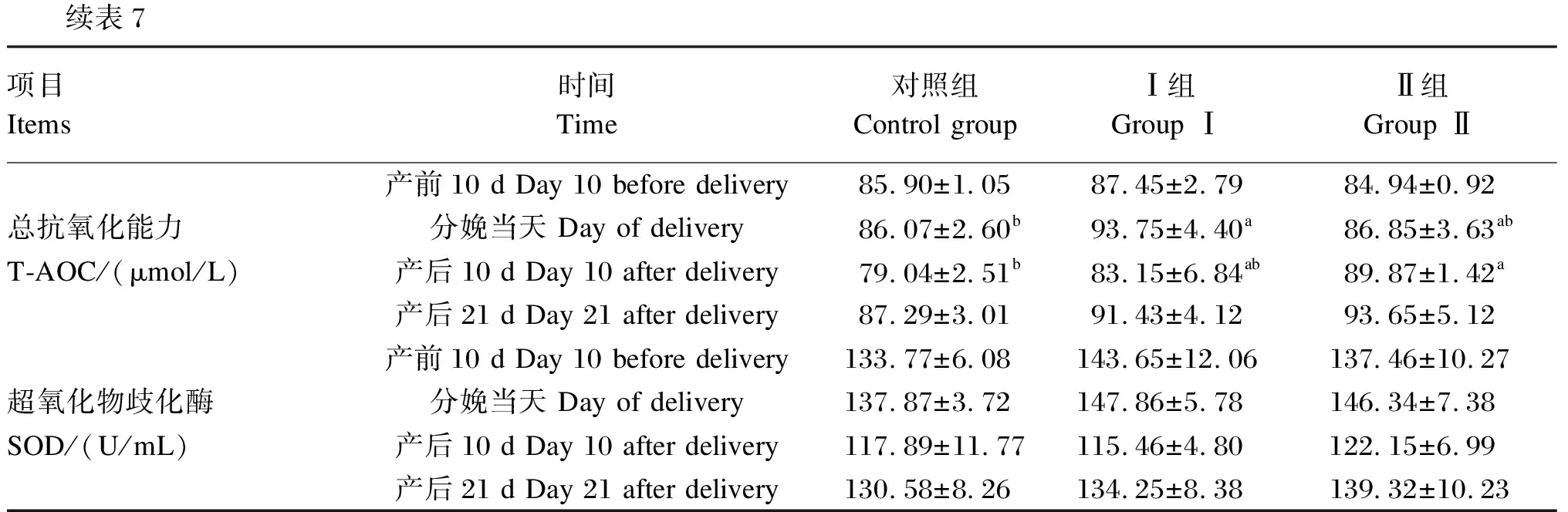
续表7项目Items时间Time对照组Control groupⅠ组Group ⅠⅡ组Group Ⅱ产前10 d Day 10 before delivery85.90±1.0587.45±2.7984.94±0.92总抗氧化能力分娩当天 Day of delivery86.07±2.60b93.75±4.40a86.85±3.63abT-AOC/(μmol/L)产后10 d Day 10 after delivery79.04±2.51b83.15±6.84ab89.87±1.42a产后21 d Day 21 after delivery87.29±3.0191.43±4.1293.65±5.12产前10 d Day 10 before delivery133.77±6.08143.65±12.06137.46±10.27超氧化物歧化酶分娩当天 Day of delivery137.87±3.72147.86±5.78146.34±7.38SOD/(U/mL)产后10 d Day 10 after delivery117.89±11.77115.46±4.80122.15±6.99产后21 d Day 21 after delivery130.58±8.26134.25±8.38139.32±10.23
2.5 围产期饲粮中添加异位酸对奶牛血清免疫能力的影响
由表8可知,在产前10 d,Ⅰ组和Ⅱ组血清中IgG含量均显著高于对照组(P<0.05);分娩当天,Ⅱ组血清中IgG和IgM含量均显著高于对照组(P<0.05);在产后10 d,Ⅱ组血清IgA含量和Ⅰ组血清IgM含量均显著高于对照组(P<0.05)。

表8 围产期饲粮中添加异位酸对奶牛血清免疫能力的影响
2.6 围产期饲粮中添加异位酸对新生犊牛血清抗氧化及免疫指标的影响
由表9可知,Ⅰ组和Ⅱ组新生犊牛血清中GSH-Px活性均显著高于对照组(P<0.05);Ⅰ组和Ⅱ组血清中T-AOC、SOD活性均高于对照组,MDA含量均低于对照组,但无显著差异(P>0.05)。Ⅰ组新生犊牛血清中IgA和IgG含量均显著高于对照组(P<0.05)。Ⅰ组和Ⅱ组新生犊牛血清中IgM含量均高于对照组,但无显著差异(P>0.05)。
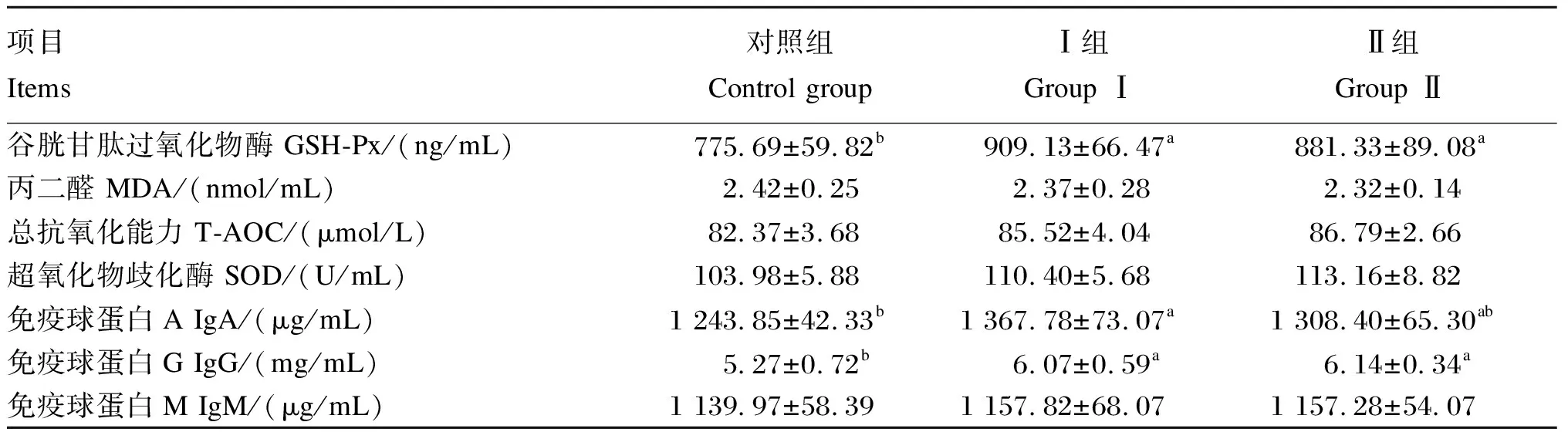
表9 围产期饲粮中添加异位酸对新生犊牛血清抗氧化及免疫指标的影响
3 讨 论
3.1 围产期饲粮中添加异位酸对奶牛生产性能的影响
3.1.1 围产期饲粮中添加异位酸对奶牛产奶量的影响
异位酸能显著提高奶牛的产奶量和饲料转化利用率。异位酸在瘤胃中被分解吸收,可增强反刍动物瘤胃内纤维降解菌的生长繁殖,促进采食量[29],提高饲粮营养物质表观消化率[30],增加VFA含量[31-33],促进MCP合成,增加氮的沉积,为机体提供更多的能量来满足泌乳。张平等[34]研究表明,添加异位酸显著提高产奶量和乳脂率,改善奶牛的产奶性能。王振权等[35]研究表明,在饲粮中添加异位酸可使每头奶牛每天产奶量增加1.80~2.27 kg。在本试验中,Ⅰ组和Ⅱ组奶牛的产奶量在产后10和21 d均显著高于对照组,产奶量持续增长。这表明在围产期饲粮中添加异位酸可以增加产奶量,增加经济效益。
3.1.2 围产期饲粮中添加异位酸对新生犊牛初生重和体尺的影响
新生犊牛的初生体重对其以后生长发育速度和生产性能直接相关,但如果体重过高会导致奶牛难产率显著提高。一般情况下,动物体生长发育情况的评判标准是体重和体尺。体重能大致反映出犊牛机体生长发育情况,而且能够评价消化系统能力、各个器官的整体发育水平以及免疫能力[36]。处于NEB或氧化应激较高的奶牛所分娩产下的犊牛普遍出现体重较轻的情况,孕后期热应激奶牛所产后代的平均初生重较低[37]。本试验中,Ⅰ组和Ⅱ组新生犊牛初生体重和体尺与对照组相比无显著差异,但有上升趋势。这表明围产期饲粮中添加异位酸对新生犊牛初生体重和体尺的影响作用不大。
3.2 围产期饲粮中添加异位酸对奶牛营养物质表观消化率和瘤胃发酵参数的影响
3.2.1 围产期饲粮中添加异位酸对奶牛营养物质表观消化率的影响
动物机体的发育离不开采食量的增加和对饲粮中营养物质的吸收利用。高爱琴[38]研究发现,异丁酸和异戊酸对饲料DM、ADF和NDF表观消化率均有一定的促进作用。李红玉等[39]研究表明,饲粮中添加异丁酸能显著提高玉米秸秆DM、有机物、ADF和NDF的降解率。在本试验中,异位酸处理组均能显著提高饲粮中DM、ADF和NDF表观消化率,但本研究中CP表观消化率也显著升高,可能原因是异位酸组的瘤胃中NH3-N含量显著降低,促进了机体对氨氮的利用率,瘤胃内环境中MCP生成量增加,从而提高了机体蛋白质利用率。
3.2.2 围产期饲粮中添加异位酸对奶牛瘤胃发酵参数的影响
瘤胃发酵参数的基本指标主要包括pH及MCP、NH3-N和VFA含量,通过这些指标可以综合反映瘤胃发酵情况。瘤胃液正常的pH一般维持在5.5~7.5。在本试验中,Ⅰ、Ⅱ组与对照组的pH无显著差异,与刘永嘉等[15]研究结果一致,说明瘤胃发酵环境不会因为添加异位酸而改变,对微生物生长繁殖的环境无显著影响。NH3-N含量可以反映出机体利用蛋白质的能力,其含量降低,氨氮转化率变高,适宜的含量能确保瘤胃中微生物的正常生长繁殖,利于MCP的合成[40]。在本试验中,Ⅰ、Ⅱ组瘤胃中NH3-N和MCP含量与对照组相比均有显著性差异,这与Allison[41]、姜菲等[42]研究结果一致,即添加异位酸可降低NH3-N含量和提高MCP含量。这说明饲粮中添加异位酸能进一步促进奶牛机体对氮的利用。VFA是反刍动物能量的主要来源,是反映饲粮发酵水平的重要指标,能够供给生命所需70%~80%的能量[43]。照日格图等[44]通过体外批量培养发现,异戊酸可以提高培养基中乙酸、丙酸、丁酸和TVFA的含量,以及乙酸/丙酸。王仁杰[45]通过体外瘤胃发酵试验发现,饲粮中添加异位酸有利于乙酸、丙酸和TVFA的合成,精料比例越高,添加异位酸的效果越好。Felix等[46]认为,异位酸可以提高泌乳奶牛瘤胃液中的乙酸含量,这与本研究结果一致,异位酸对瘤胃发酵有促进作用,能提高乙酸、TVFA含量,但各试验组间丙酸和丁酸含量相比无显著差异。瘤胃中乙酸含量的显著提高会导致瘤胃发酵类型趋于乙酸型发酵,这种发酵类型有利于乳脂的合成,从而提高奶牛的产奶量和乳中乳脂率。
3.3 围产期饲粮中添加异位酸对奶牛血清生化指标的影响
动物机体血清蛋白含量的高低能反映出其对蛋白质的吸收、消化和分解等代谢情况,其含量越高说明机体对饲料的利用率越高,更加促进机体的生长发育。支链氨基酸能调控机体蛋白质的合成,抑制其他组织器官的蛋白质分解[47]。李明等[48]研究表明,在中、高非蛋白氮(NPN)饲粮中添加异位酸可显著提高血液总蛋白、GLB和GLU含量。Felix等[49]的研究结果表明,异位酸能促进MCP的生长速率,提高机体的氮循环和氮留存,进而提高血清蛋白含量。在本试验中,围产期饲粮中添加异位酸,Ⅰ组和Ⅱ组奶牛血清中GLB和ALB含量在各个时期均有所提升。
血清中UN含量与饲料利用率有关。血液中UN含量减少,机体对蛋白质利用率升高,从而为其他部位的蛋白质沉积提供足够的氨基酸和其他原料[50]。GLU是机体主要的能源物质,饲粮中的能量物质越高,机体GLU含量也会越高,机体糖代谢就越旺盛。张斌等[51]研究表明,饲粮中添加20 g/(d·头)异位酸可降低血清UN含量,提高GLU含量。王仁杰[45]研究表明,南江黄羊饲粮中添加7.1 g/(d·头)的异位酸可降低血清UN含量。血清中GLU含量的增加会导致奶牛产奶量的上升[38],这与本试验结果一致。支链脂肪酸具有调控肝脏脂质和糖代谢的功能[52]。在本试验中,整个围产期Ⅰ组和Ⅱ组奶牛血清中UN含量与对照组相比均下降,血清GLU含量与对照组相比均出现一定程度的上升。原因是饲粮中添加异位酸能增加机体对氨氮的利用率,促进瘤胃微生物的生长繁殖,提高血糖水平,为机体提供更多的能量来源;乙酸是合成体脂的前体物,饲粮中添加异位酸可促进瘤胃内乙酸的合成,进而促进动物机体脂肪的沉积,有利于动物生长发育[53-55]。
TG和TC是脂肪代谢的主要生成物,其含量与脂肪代谢程度成反比。Ⅰ组血清中TG含量在分娩当天显著低于Ⅱ组,可能原因是Ⅰ组奶牛在分娩当天机体能量供应不足或能量摄入量过低,导致TG含量下降;各试验组间血清TC含量均无显著差异。
AST和ALT是反映肝脏功能的重要相关指标[56],在奶牛产前,若肝脏处于代偿状态、功能受损时,会导致血液中AST和ALT活性急剧上升。本试验中,Ⅰ组血清中ALT活性在产前10 d显著高于对照组,可能原因是Ⅰ组奶牛肝脏功能受到抑制,肝细胞受损导致线粒体损伤从而引起ALT活性显著升高。
3.4 围产期饲粮中添加异位酸对奶牛血清抗氧化能力的影响
动物机体的抗氧化能力与机体健康密不可分,通过合理途径补充适量的绿色抗氧化剂可以提高动物机体抗氧化能力。MDA是脂质过氧化的终产物之一,其含量可以直接反映机体受自由基攻击的程度,间接说明细胞的损伤程度。血液中GSH-Px活性越高表明动物的抗氧化能力越强,它作为动物机体重要的抗氧化剂和自由基清除剂,可以提高机体免疫力和抗病能力[57]。T-AOC是反映动物抗氧化能力的综合指数,其水平高低能反映动物机体的自由代谢和抗氧化系统补偿外部刺激的能力。SOD是体内清除超氧阴离子自由基的重要因子并参与转换氧自由基的过氧化物酶,平衡氧化与抗氧化机制,保护细胞免受伤害[58]。奶牛处于围产时期时,机体免疫功能急剧下降,围产期奶牛氧化应激的发生可能是由于围产期奶牛活性氧(ROS)产量急剧增加或抗氧化水平降低导致的生理失衡。体内氧化与抗氧化失衡,导致机体处于氧化应激状态,持续的氧化应激会导致机体的慢性炎症[59]。李爱玲等[60]研究发现,支链氨基酸能显著降低大鼠体内MDA含量,从而防止体内GSH-Px和SOD的活性降低。在本试验中,添加异位酸能提高血清GSH-Px活性和T-AOC,降低血清MDA含量,对SOD活性无显著影响。这说明在围产期奶牛饲粮中添加异位酸可以提高奶牛机体的相关抗氧化因子活性,从而提高抗氧化能力。目前,关于异位酸以及饲粮中添加异位酸对反刍动物抗氧化能力影响相关的研究报道较少,还有待进一步研究。
3.5 围产期饲粮中添加异位酸对奶牛血清免疫能力的影响
围产期是奶牛一个重要而又特殊的时期,在此阶段奶牛的免疫系统变得错综复杂。奶牛处于分娩应激状态下会导致机体血清免疫球蛋白整体水平降低,影响免疫功能。免疫球蛋白是指受抗原刺激后所产生的蛋白质,与抗原发生免疫反应,通过抗体介导的免疫途径从动物体内清除外来病原体和抗原物质,从而阻断病原体对动物机体的危害[61]。免疫球蛋白含量在一定程度上可以反映反刍动物的免疫能力,主要包括IgG、IgA和IgM,其中IgG含量最高[62]。本试验结果表明,饲粮中添加异位酸可提高围产期奶牛血清中IgA、IgG和IgM含量。原因可能是异丁酸能刺激泌乳奶牛机体的免疫功能,使血液中免疫球蛋白的含量增加[51]。在本试验中,围产期饲粮中添加异位酸能提高动物机体的免疫能力,和预期试验结果一致。
3.6 围产期奶牛饲粮中添加异位酸对新生犊牛血清抗氧化及免疫指标的影响
动物机体在正常情况下,氧化与抗氧化机能处于相对平衡状态,当动物受到体内或外界因素导致应激时,机体的这种动态平衡就会破坏,产生大量的自由基。犊牛通过摄入初乳来启动被动免疫以抵御疾病和感染。在本试验中,Ⅰ组和Ⅱ组新生犊牛血清GSH-Px活性显著高于对照组,MDA含量、T-AOC和SOD活性无显著差异。Ⅰ组和Ⅱ组新生犊牛各抗氧化指标均比对照组高的原因可能是Ⅰ组和Ⅱ组奶牛通过胎盘输送给犊牛的营养物质、氨基酸及蛋白质的含量均高于对照组。这表明在奶牛围产前期饲粮中添加异位酸能一定程度提高新生犊牛的抗氧化能力。
免疫球蛋白不能从母体传递给胎儿的原因是母牛胎盘的特殊结构[63],母牛在妊娠期间母体免疫球蛋白类物质不能穿过胎盘,导致新生犊牛出生时缺乏免疫保护能力,若未能及时吃到初乳,犊牛血液中将不能产生抗体,机体免疫力缺乏,进而导致后期发育受阻[64]。初乳中免疫球蛋白的含量最高。新生犊牛饲喂初乳后,血清中IgG含量迅速升高,主要原因是犊牛胃肠道可将初乳中的IgG非特异性转运至血液,初乳中的胰蛋白酶抑制肽可抑制肠胰蛋白酶活性,保护IgG免受胃肠道分解破坏[65]。本试验结果表明,在饲喂初乳后,Ⅰ组和Ⅱ组新生犊牛血清IgA和IgG含量均显著高于对照组,IgM含量无显著影响。原因是在围产前期奶牛饲粮中添加异位酸提高了奶牛的初乳质量和免疫球蛋白含量,从而提高新生犊牛的免疫能力。
4 结 论
① 在围产期饲粮中添加异位酸可提高奶牛的生产性能和产后泌乳初期产奶量。
② 在围产期饲粮中添加异位酸可提高奶牛DM、CP、NDF和ADF表观消化率;提高瘤胃中MCP、乙酸和TVFA含量,降低NH3-N含量。
③ 在围产期饲粮中添加异位酸可降低奶牛血清中UN含量,提高ALB和GLB含量;增强机体抗氧化及免疫能力。在奶牛围产前期饲粮中添加异位酸可一定程度上增强新生犊牛的抗氧化和免疫能力,改善犊牛健康状况。
④ 在本试验条件下,异位酸在奶牛围产期饲粮中适宜的添加量为60 mL/(d·头)。
猜你喜欢
动物营养学报(2022年8期)2022-09-01
中华养生保健(2020年2期)2020-11-16
中成药(2018年3期)2018-05-07
婚育与健康(2017年7期)2017-09-12
动物营养学报(2017年8期)2017-08-16
现代检验医学杂志(2016年4期)2016-11-15
兽医导刊(2016年12期)2016-05-17
兽医导刊(2016年12期)2016-05-17
动物营养学报(2015年9期)2016-01-07
当代畜禽养殖业(2014年5期)2014-08-31
