
小麦芒长近等基因系的表型及遗传分析
2023-11-07 06:26:14陈真真周国勤陈金平石守设谢旭东申冠宇
湖北农业科学 2023年10期
陈真真,周国勤,陈金平,石守设,谢旭东,申冠宇
(信阳市农业科学院,河南信阳 464000)
在结构和功能上芒与叶片有许多相似之处,芒是叶的变态,是小花外稃的延伸。芒是小麦重要的形态标记和穗部重要的光合器官。小麦(Triticum aestivumL.)芒中的绿色细胞富含叶绿体与线粒体,麦芒中的叶绿体具有光合作用的结构和光化学活性[1]。由此可知,芒在光合作用和碳同化的过程中也具有非常重要的作用。大麦栽培品种Morex 的内外稃、种子和芒的基因表达表明,与光合作用、叶绿素生物合成和活性氧清除有关的基因在麦芒中优先表达;在外稃中防御相关的基因优先表达。小穗器官对于子粒灌浆期的贡献还没有完全阐明,而在大麦中芒的光合作用占穗部光合作用的90%,内外稃是大麦穗部第二重要的光合器官[2]。芒与干旱有关的基因多于内外稃[3],与旗叶相比由于芒表面有硅质和角质层从而可以保持较高的电子运输速率和相对含水量[4]。对郑引1 号、丰产3 号和矮丰3 号3 个小麦品种进行剪芒处理结果发现,3 个品种千粒重分别比对照减少了8.7%、2.2%和5.3%,穗粒数分别减少了0.8%、0.5%和2.2%。杨兆生等[5]、王瑞清等[6]、冯朝章等[7]的试验结果与之类似,且粒重与芒的有无、功能期的长短有密切的关系,不同品种间存在显著差异。
小麦芒的性状分为芒长、芒抑制、钩芒等几种情况。芒的发育由A、B1、B2、B3和Hd共5 个主效基因和一些微效基因共同决定。其中,A基因具有促进作用;B1、B2、B3基因具有抑制作用,B1具有较强的抑制作用,B2的抑制作用居中,B3具有最弱的抑制作用;Hd基因调控钩芒的产生。早期的研究表明,芒性状是由一对等位基因控制的,无芒表现为显性,有芒表现为隐性。黄瑾等[8]采用常规杂交方法,王彦梅等[9]应用单体分析法,郑建敏等[10]利用常规杂交法对芒性状进行遗传分析,结果表明性状分离符合孟德尔规律,无芒对有芒为显性,且受一对显性基因控制。Shin 等[11]用杂交的98 个F2代植株对短芒基因lks2进行分子定位,Takahisa 等[12]完成了大麦芒基因lks2的精细定位,然而目前对于麦芒的遗传与发育还缺乏系统的研究。
作者在前期的杂交育种工作中从CSAM 的F8代单株分离后代中获得一对芒长度存在差异、遗传背景一致的近等基因系CSAM1(长芒)与CSAM2(短芒)。本研究通过对其性状的调查以及对穗部光合作用和千粒重的比较,进一步证明了芒在小麦生育过程的重要作用。同时利用CSAM1(长芒)与CSAM2(短芒)杂交获得分离群体,并对控制二者芒长度差异的基因进行初步遗传分析,以期为芒性状相关基因的后续研究和小麦的遗传改良研究奠定理论基础。
1 材料与方法
1.1 材料
CSAM1 和CSAM2 是信阳市农业科学院利用美国软红和中国春杂交的后代,为F8代中选育出的2个株系,连续多年种植性状表现稳定。利用CSAM1(长芒)与CSAM2(短芒)杂交获得F1,自交获得F2分离群体和F2∶3家系。
1.2 田间种植和性状调查
2个材料均种植于信阳市农业科学院试验基地,每种材料种植2 行,行长为5.00 m,行距为0.33 m,3 次重复。F2群体按单粒播种的方式种植,行长为2.00 m,行间距为0.33 m,株距为0.15 m;F2∶3家系群体按单粒播种种植成株行,行长为2.00 m,行间距为0.33 m,株距为0.15 m,田间管理按大田常规标准进行。用LI-6400 型光合仪进行旗叶叶片光合速率的测定。测定时间为2020 年5 月6 日,每个处理重复测定3 次。 灌浆速率测定于材料花后10 d(2020 年4 月26 日)开始,每5 d 取1 次样直至小麦收获。每个材料随机取穗,脱粒烘干称量干重。
1.3 田间芒长表型评估
将小麦穗部平均分成上、中、下3 个部分,蜡熟期后取10 株植株,每株在主茎穗部上、中、下部分别选取3 根芒用直尺测量,取平均值,记录每一部分的数据用作表型及遗传分析。
1.4 遗传分析
利用CSAM1(长芒)与CSAM2(短芒)杂交获得F1,观察F1代芒性状的表现,F1代自交获得F2分离群体和F2∶3家系,统计F2代出现的芒类型,经过卡方检验看其是否遵循1∶2∶1 或3∶1 的孟德尔遗传定律分离比。
2 结果与分析
2.1 CSAM1 和CSAM2 植株性状表现
对CSAM1(长芒)和CSAM2(短芒)2 个材料的性状进行调查,考察的农艺性状包括株高、穗长、壳色、粒色、每株穗数、穗粒数、千粒重、芒类型以及抽穗期和开花期。结果(表1、图1)表明,2 个材料芒长差异达极显著水平(P<0.01),而生育期、株高、穗长、壳色、粒色、每株穗数、小穗数和穗粒数等性状差异不明显,基本一致。这说明CSAM1(长芒)和CSAM2(短芒)可作为一对良好的近等基因系用于芒长性状的研究。

表1 CSAM1(长芒)和CSAM2(短芒)的性状特点

图1 CSAM1(长芒)和CSAM2(短芒)的穗部
2.2 CSAM1(长芒)和CSAM2(短芒)旗叶光合速率及灌浆速率差异
花后20 d 时,分别测定了CSAM1(长芒)和CSAM2(短芒)旗叶的光合速率,结果见表2。其中,长芒材料CSAM1 旗叶的净光合速率明显高于短芒材料CSAM2 旗叶的净光合速率,且部分时间点差异达显著水平(P<0.05),说明芒的存在显著增加了旗叶的光合能力。从花后10 d 开始每隔5 d 取材,分别测定比较2 个材料的灌浆速率,可以看出,开花后15 d 左右子粒千粒重增长迅速,2 个材料的千粒重差异不明显,但从花后25 d 开始,长芒材料CSAM1 千粒重高于短芒材料CSAM2,至收获CSAM1 的平均千粒重为34.3 g,CSAM2 的平均千粒重为33.0 g(表2)。这证明芒在子粒灌浆后期同化产物的积累中起到了重要作用,从而造成二者千粒重的差异。
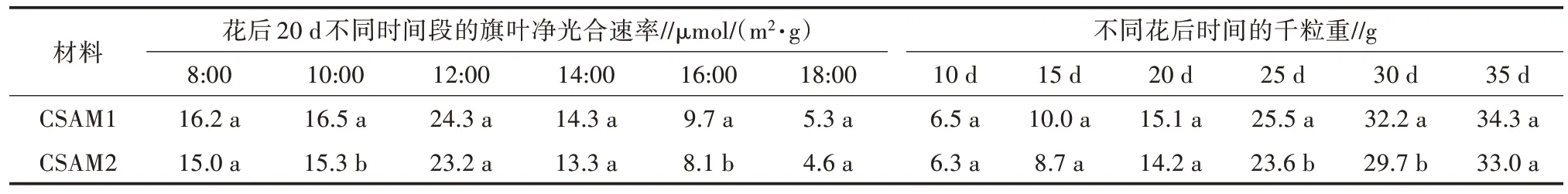
表2 CSAM1(长芒)和CSAM2(短芒)旗叶的光合速率和灌浆速率
2.3 群体芒长表型评估鉴定
2 个亲本中,CSAM2(短芒)表现为短芒,CSAM1(长芒)表现为有芒(穗部平均芒长大于4.2 cm)。由于芒的长度不同以及在穗部着生的位置不同,于是对亲本和F1群体穗部各个部位芒长进行评估鉴定,结果发现,无芒亲本芒长不超过0.5 cm,中部芒长不超过0.25 cm,下部没有芒;有芒亲本上部和中部芒长都大于4.2 cm,下部平均芒长超过2.8 cm。将无芒亲本与有芒亲本杂交,杂交F1代个体全部表现为有芒,但都为短芒,且各部位芒长介于双亲之间。同时发现,2 个亲本和杂交后代F1群体穗部下部芒长最短,中部芒长最长,上部芒长居于二者之间。但通过性状调查发现,虽然亲本中国春是无芒小麦品种,但其穗上部存在少量很短的芒,因此用上部芒长小于0.6 cm 作为划分小麦群体无芒的标准。
2.4 群体芒长遗传分析
CSAM1(长芒)与CSAM2(短芒)的杂种F1代全部表现为短芒,其F2代共调查983 株,其中长芒246株,短芒737 株。对F3代调查发现,737 个短芒F2植株的后代中,有529 个短芒F2植株的后代发生了芒长性状的分离,208 个短芒F2植株的后代未发生分离。246 株长芒F2的后代植株全部表现为长芒。χ2检验结果表明,F2代短芒植株和长芒植株的分离比例符合3∶1 的分离比例(表3)。以上结果说明,本试验所研究材料CSAM1 与CSAM2 的芒长性状受单基因控制,其中短芒对长芒为显性。

表3 F2群体的遗传分析以及卡方检验
3 讨论
芒作为小麦穗部器官的组成之一,具有促进小麦光合作用、推动种子传播、防御食草动物、干旱条件下增加产量等作用[13]。在植物长期进化过程中,任何组织器官的存在都是适应自然界和自然选择的结果。芒作为小麦穗器官的组成,是小麦长期驯化和适应环境的结果[14]。芒作为穗部外稃和护颖的针状延伸结构,着生有硅质倒刺,因此芒对防止鸟虫害和种子传播是有利的[1,4]。从芒的超显微结构来看,小麦的芒中富含叶绿体和线粒体,具有同叶片中相同的结构和光化学特性,能极大地增加穗部光合面积[1]。小麦的芒对光合作用及蒸腾作用有积极影响,并对干旱具有适应性属性[1,8]。芒具有提高光合面积、光合效率和提高小麦产量的作用[10]。因此,芒的有无也对产量有直接影响。小麦的芒是叶的变态,是由小花外稃的末端延伸形成的针状物,在内部形态结构和功能上与叶片有许多共同之处[15]。在对小麦光合效率和产量研究方面,除叶片以外,芒也起重要作用[8]。从结构上讲,芒具有对小麦生长和繁殖有利的结构特征。还有研究表明,在逆境条件下,无芒小麦比有芒小麦产量降低的幅度要大[13]。因此,探究小麦芒的形成发育及其背后的遗传分子机理,不仅有助于小麦抗逆性和产量的提升,而且为小麦的遗传改良提供了新的思路。
Duwayri[16]研究了在干旱条件下旗叶和芒的去除对小麦子粒产量的影响,结果表明,只去除旗叶、只去除芒和同时去除旗叶+芒的组合产量分别降低了10.7%、15.9% 和21.2%,单株粒数分别减少了11.1%、11.3%和11.2%。研究还表明,适应干旱地区条件品种的芒对产量的的贡献要大于旗叶对产量的贡献[17]。Khaliq 等[18]的研究则表明,去除旗叶和去除芒相比,去除芒对产量相关性状的影响较小,去除旗叶对产量相关性状的影响较大,而与单独去除相比,二者共同去除均具有更显著的作用。这表明在选择具有良好的光合活性和高产小麦品种时,旗叶+芒可以用作形态标记。
目前人们多是通过采用剪芒的方法研究芒对产量形成的影响,芒对子粒灌浆的影响通过剪芒和未剪芒材料的千粒重和产量差异来说明[16,19]。但是这种研究方法无法避免剪芒造成的机械伤害的影响,同时也难以进行较大面积的试验。而利用近等基因系研究芒在子粒形成过程中的贡献则不会存在以上这种影响,同时可以解决利用不同小麦品种进行研究所存在的遗传背景差异问题。 也有一些获得不同芒长性状近等基因系的报道,Weyhrich 等[20]利用3 个有芒硬粒小麦TAM107、Mustang、Century 与无芒软红冬小麦品系McNair1003 杂交,杂交后代获得了以这3 个硬粒小麦品种为背景的无芒、有芒近等基因系,并在光合作用和水分利用效率的试验中比较了近等基因系之间的差异,同时试验也证明在供水充足的条件下,有芒穗部的光合效率明显高于无芒小麦的穗部光合能力。Tsunewaki 等[21]获得了小麦品种S-615 遗传背景的芒B1基因的近等基因系,发现2 个近等基因系不仅芒长存在差异,其第一节间长度、花药长度、收获指数、穗轴密度、小穗密度等性状也存在明显差异。这可能是由于回交代数比较少,所获近等基因系除了B1位点存在差异外,控制其他性状的基因位点也存在较大差异。
研究者对小麦芒长基因进行了初步定位。宫希等[22]利用长芒的普通小麦郑麦9023 与无芒的西藏半野生小麦Q1028 构建了一个重组自交系群体(186个株系),采用SSR 和DArT 分子标记构建了覆盖小麦全基因组的遗传图谱(2 597 cM),基于重组自交系群体2 年芒长表型数据,采用ICIM 作图法对小麦芒长性状进行QTL 定位分析,将麦芒抑制基因Hooded定位在4A 染色体短臂的标记Xgpw4448 和wPt-1038 之间,遗传距离为9 cM。金迪等[23]利用短芒材料六柱头与长芒材料石矮1 号构建的F2群体(SL-F2)研究了芒的遗传与发育,将B2精细定位到了6B 染色体dCAPS18 和dCAPS3 标记之间,遗传距离为0.29 cM,对应着中国春参考基因组(IWGSC.Ref. V 1.0)4.84 Mb 的物理区间。在B2定位区间共有61 个基因,其中5 个在中国春穗部特异表达,TraesCS6B02G264400 在中国春和Azhurnaya 幼穗表达差异显著。
本研究表明,CSAM1(长芒)和CSAM2(短芒)材料的农艺性状如穗型、株高、壳色、粒色、穗长、小穗数和穗粒数均一致,仅芒长和千粒重存在差异。目前,小麦芒的发育及其遗传调控机制尚不明确,充分了解芒的遗传基础,发挥其光合潜能,对小麦的遗传改良和分子辅助育种具有十分重要的理论和实践意义。这套优良的试验材料为研究芒的作用、小麦芒抑制基因的克隆和相关功能研究提供了重要的参考价值。
猜你喜欢
区域治理(2022年40期)2022-11-27 04:01:54
作物学报(2022年9期)2022-07-18 00:58:42
干旱地区农业研究(2021年6期)2021-12-08 07:52:22
今日农业(2021年17期)2021-11-26 23:38:44
浙江农业科学(2019年11期)2019-11-09 01:37:48
动漫界·幼教365(小班)(2019年10期)2019-10-28 02:04:20
动漫界·幼教365(大班)(2019年10期)2019-10-28 01:54:09
动漫界·幼教365(中班)(2019年10期)2019-10-28 01:53:17
中央民族大学学报(自然科学版)(2016年3期)2016-06-27 07:55:28
广西林业科学(2016年2期)2016-03-20 05:53:36
