
冲绳海槽南部热液区附近柱状沉积物中微生物的垂直分布多样性及其对环境因子的响应
2023-11-03 02:47邓兴渝曹文瑞姜明玉曾志刚常凤鸣宋召军
海洋地质与第四纪地质 2023年5期
邓兴渝,曹文瑞,姜明玉,曾志刚,常凤鸣,宋召军
1. 山东科技大学地球科学与工程学院,青岛 266590
2. 中国科学院海洋研究所海洋地质与环境重点实验室,青岛 266071
深海沉积环境不仅具有低温、高压等特点,而且还含有复杂和独特的化学成分,不同深海生境(如热液喷口、冷泉、多金属结核区和海山区等)的微生物群落存在显著差异[1],它们不仅参与元素循环,同时其参与的氧化还原过程也非常复杂[2]。而深海热液区则代表了地球上最具物理和化学多样性的生境[3-4],其化学梯度和热梯度为微生物群落提供了广泛的生态位[5-6]。
为了研究与深海热液环境相关的微生物多样性,研究人员利用纯培养和免培养的方法,发现了包括深海热液喷口在内的极端环境中的微生物群落[7]。尽管纯培养的方法对于利用模式微生物来解释验证热液发生过程非常关键,而且对于发现新物种、新的天然活性产物等以及其他有工业应用潜力的天然产品也有重要意义[8-9],但是由于热液环境的微生物分离培养较为困难,所以这方面的研究目前还较少。Wang 等[10]从西北印度洋卡尔斯伯格洋中脊热液区成功分离与纯化到了代表性的菌种,首次证实了该类群微生物具有硫还原功能,并在氢气的氧化过程中获得能量,揭示了其单质硫还原的关键基因和代谢途径。杜瑞等[11]曾对冲绳海槽热液区的硫氧化细菌进行了分离培养,并对其代谢类型和活性进行了初步探究,为深海热液系统的硫循环机制奠定了基础。与纯培养的技术相比,免培养的方法具有检测整个微生物系统多样性的优势,包括尚未培养的微生物的多样性[12-14]。迄今为止,利用免培养方法对冲绳海槽热液区微生物群落的研究相继报道[15-21],但是这些研究对象大多是热液区的表层沉积物。
冲绳海槽是西北太平洋的新近纪弧后裂谷盆地,存在着许多活动热液区,在气候生态、海洋矿产资源[22-23]和营养循环中发挥着重要作用[24]。冲绳海槽南部,平均水深在2 000 m 以上[25-26],除具有一般深海环境的高压、低温和黑暗等特点外,该区域还含有丰富的金属资源,其包含锰、铁、镍、钴和铜等几十种元素[27]。本研究样品位于第四与那国(Yonaguni Knoll Ⅳ)热液区以及唐印(Tangyin)热液区附近。其中第四与那国热液区位于长约1 000 m、宽约500 m 的狭长的深谷区,活动的热液区呈南-北向排列[28-29]。唐印热液区(25°04′N、122°34′E)则是2014 年HOBAB3 航次利用“科学号”科学考察船新发现的,它位于雨花山海丘北坡,水深1 206 m,在本区发现了旺盛的海底热液活动和密集的化能自养生物群落[28,30]。目前,关于冲绳海槽南部HOBAB4-S2 沉积物柱状样的研究,主要是对其进行了年代学分析、粒度分析、矿物学分析(硫化物矿物和黏土矿物)、沉积物地球化学以及单矿物的元素和硫同位素组成等方面的研究[28]。然而应用微生物学和分子生物学等技术手段对该样品的微生物群落组成进行深入调查,并结合环境因子进行分析的研究尚未开展。
因此,本研究以冲绳海槽南部热液区附近HOBAB4-S2 沉积物柱中不同层位的沉积物样品为研究对象,基于纯培养方法和16S rRNA 基因的V3-V4 区域Illumina 平台的高通量测序技术,对该沉积物柱的微生物垂直分布多样性开展调查,并结合样品的地球化学特征分析微生物群落组成对关键环境因子的响应。
1 材料与方法
1.1 样本来源
本研究样品是2016 年5 月由中国科学院海洋研究所“科学”号考察船在HOBAB4 航次期间通过重力沉积物柱取样器从冲绳海槽南部的S2 站采集的,该HOBAB4-S2 沉积物岩芯全长4.77 m,其位于唐印热液区和第四与那国热液区之间的裂谷盆地(24°52'49.91″N、122°37'19.70″E),水深为1 505 m[28]。
1.2 元素地球化学特征
分层处理后的沉积物样品被冷冻干燥,选取所需层位的沉积物样品,每个层位取>10.0 g 沉积物样品于60℃烘箱中烘干,将烘干样品用玛瑙研钵研磨并过筛(100 目)。利用S8 Tiger X 射线荧光光谱仪(X-ray Fluorescence,XRF)(Bruker,德国)测定每个样品中的主、微量元素组成情况,检出限为0.8~220 μg/g;利用PE 2400 II CHNS/O 分析仪(Perkin-Elmer,美国)测定每个样品的总有机碳(TOC)、总碳(TC)和总氮(TN)含量, TOC、TC 和TN 的检测下限分别为0.07%、0.07%和0.02%。上述分析测试均在中国科学院海洋研究所分析测试中心完成,分析结果用于了解沉积物样品的地球化学特征和初级生产力。
1.3 主要试剂和培养基
在无菌条件下,将1 g 沉积物样品加至9 mL 无菌海水中,充分震荡,静置10 min 后取上清液1 mL继续用无菌海水进行梯度稀释,最终获得10-1、10-2、10-3、10-4、10-5共计5 个浓度梯度的稀释液。每个梯度分别取100 μL 涂布在分离培养基平板上,随后置于25℃培养5—7 天。本研究所用的分离培养基有2216E[31]、SPG 和ASC 培养基[11,32]。
在25℃培养箱中培养后,观察平板中菌落形态,对菌落大小、颜色、边缘、透明度等特征进行描述和记录。根据形态特征,挑取平板中不同形态的单菌落,在对应的培养基平板上进行划线纯化。将纯化后的菌株接种对应的液体培养基培养,离心后将菌体细胞重悬于20%的甘油中,并转入冻存管,放在-80℃下保藏。
1.4 菌株DNA 提取、16S rRNA 基因扩增和分类鉴定及系统发育分析
采用煮沸法提取菌株的基因组DNA,选择27F/1492R 通用引物(27F:5'-AGAGTTTGATCMTGGCTCAG-3';1492R:5'-TACGGYTACCTTGTTACGAC-3')对菌株的16S rRNA 基因进行PCR 扩增[33,34]。PCR 扩增使用以下程序:在94℃下初始预变性3 min;94℃变性45 s,55℃退火60 s,72℃延伸90 s,30 个循环;72℃继续延伸10 min,于4℃保存[35]。PCR 产物在1%琼脂糖凝胶上通过电泳进行检测,将合格的PCR 产物送至北京六合华大基因科技有限公司进行测序。
将所得的16S rRNA 基因序列在NCBI 数据库中进行BLAST 比对( https://blast.ncbi.nlm.nih.gov/Blast.cgi),同时在EzTaxon-e 服务器(http://www.ezbiocloud.net)中计算所得序列与亲缘模式菌株的16S rRNA 基因序列的相似性。使用CLUSTAL X 对所有获得的16S rRNA 基因序列和所对应的亲缘模式菌株的16S rRNA 基因序列进行多重比对。随后使用MEGA(version 11)[36]进行系统发育分析,并构建基于最大似然法( Maximum likelihood approach)[37]的系统发育树。构建过程使用complete deletion 选项进行分析,并根据双参数模型(Kimura's two-parameter model)[38]计算距离,进行1 000 次重复验证。
1.5 环境样品DNA 提取扩增、高通量测序和数据分析
对于选取的每个沉积物样品,分别采集三份重复样,利用试剂盒Fast DNA Soil-Direct Kit(MP BIO,美国)提取沉积物样品的总DNA。DNA 浓度和完整性分别通过NanoDrop 2000 分光光度计(Therm,USA)和琼脂糖凝胶电泳进行评估。以提取的样品DNA 为模板,用引物343F(5'-TACGGRAGGCAGCAG-3')和798R(5'- AGGGTATCTAATCCT-3'),对16S rRNA基因的V3-V4 区进行扩增[39]。PCR 产物使用电泳检测,检测后使用磁珠纯化,纯化后作为二轮PCR模板,进行二轮PCR 扩增,并再次使用电泳检测,检测后使用磁珠纯化,纯化后对PCR 产物进行Qubit定量。根据PCR 产物浓度进行等量混样,并上机测序。
获得的高通量原始序列数据用cutadapt 软件剪切掉引物序列,并使用DADA2 将上一步合格的双端原始数据(raw data)使用Qiime2 默认参数进行质量过滤、降噪、拼接及去嵌合体等质控分析之后,对序列进行去重;得到代表序列及ASV(Amplicon Sequence Variant,扩增子序列变异)丰度表格。使用QIIME 2 软件包挑选出各个ASV 的代表序列后,并将所有代表序列与Silva(version138)数据库(https://ftp.arb-silva.de)进行比对注释。物种比对注释使用q2-feature-classifier 软件默认参数进行分析。菌群丰富度指数(Chao)、多样性指数(Shannon和Simpson)以及发育树谱系多样性指数(PD whole tree)等α多样性指数使用QIME 计算。由于每个样品的3 个重复呈现的微生物群落组成是相近的,在进一步分析中使用了3 个重复的平均值。利用主坐标分析(Principal coordinates analysis,PCoA)评估不同样品之间的相似性及差异性,同时利用冗余分析(Redundancy analysis,RDA)讨论环境因子、样品、微生物类群三者之间的关系。上述16S rRNA基因扩增子测序和数据分析在青岛欧易生物科技有限公司和欧易生物云平台(https://cloud.oebiotech.com/)完成。
1.6 登录序列号
本研究可培养菌株16S rRNA 基因序列已提交GenBank,登录号见表1。本研究16S rRNA V3-V4 区高通量测序序列已提交GenBank,登录号为:SRR24489263-SRR24489263319。

表1 本研究使用样品的地球化学组成Table 1 Geochemical compositions of the samples used in this study
2 结果
2.1 主量元素和微量元素组成分析
冲绳海槽南部沉积岩芯HOBAB4-S2 的主量元素分析结果表明,其SiO2的含量较高,其次是Al2O3含量,MgO、Na2O 含量则较低。结合前人测得的部分数据[40],其中SiO2含量的变化范围为55.35%~69.31%,平均值59.56%(n=61);Al2O3含量的变化范围为10.01%~16.86%,平均值15.54%(n=61);MgO 含量的变化范围为1.91%~2.71%,平均值2.42%(n=61);Na2O 含量的变化范围为1.78%~2.46%,平均值2.09%(n=61)。Cu 含量为18.9×10-6~65.0×10-6,平均值38.35×10-6(n=61),Zn 和Pb 含量分别为83.4×10-6~356×10-6和27.6×10-6~160×10-6,平均值分别为129.82×10-6(n=61)和64.49×10-6(n=61)。在沉积岩芯HOBAB4-S2 孔中发现了两个富含Cu-Zn-Pb 的层(图1),分别在166 cmbsf 和444 cmbsf处。我们研究的冲绳海槽南部沉积岩芯HOBAB4-S2 微量元素含量普遍较高,其Ba、Zr、Ti 的含量平均值分别为507.74×10-6、193.32×10-6、4 693.74×10-6(n=19)。此外,该沉积物柱的总碳、总氮和总有机碳含量测试结果表明,其总碳、总氮和总有机碳平均含量分别为1.42%、0.13%、0.88%,且总碳在30 cmbsf 处的沉积物中含量最低,而总氮在171 cmbsf 处的沉积物中含量则最低,分别为1.10%和0.07%(表1)。

图1 不同层位沉积物中Cu、Zn、Pb、TC、TN 和TOC 含量变化部分数据来自Yang et al[40]。Fig.1 Variations in Cu, Zn, Pb, TC, TN, and TOC concentrations in different layersParts of data are from Yang et al[40].
2.2 可培养细菌的分离鉴定及其多样性
共从冲绳海槽沉积物中分离获得64 株菌种,随后通过近全长16S rRNA 基因序列分析鉴定。结果表明,这些菌株属于4 个门、5 个纲、7 个目、14 个科、25 个属;有38 株、1 株、21 株、4 株分别属于Proteobacteria(变形菌门)、Bacteroidota(拟杆菌门)、Bacillota(芽孢杆菌门)、Actinomycetota(放线菌门)。其中,Proteobacteria 是最大的一个类群,包含两类,即19 株Alphaproteobacteria(α-变形菌纲)和19 株Gammaproteobacteria(γ-变形菌纲)。在目水平上,21 株Bacillales(芽孢杆菌目)是优势类群,其次是19 株Oceanospirillales( 海洋螺菌目) 、 12 株Rhizobiales(根瘤菌目)、4 株Sphingomonadales(鞘脂单胞菌目)和4 株Pseudonocardiales(假诺卡式菌目)。对于其他3 个目,每个目的分离菌株都少于4 株。在科水平上,最丰富的科是15 株Alcanivoracaceae(食碱菌科)、13 株Bacillaceae(芽孢杆菌科)和7 株Phyllobacteriaceae(叶杆菌科)。其他9 个科中的每科都少于7 株。从属上看,其中占据优势地位的是Alcanivorax的菌株,占比达到了分离菌株总数的1/4,其次为Paenisporosarcina,占比约11%。从不同沉积物层位分离得到菌株的详细信息见表2。
将所有菌株及其已知亲缘模式菌株一起构建基于16S rRNA 基因序列的系统发育树(图2)。结果表明,作为占据优势数量的Proteobacteria 中的Alcanivorax xenomutans,从表层到深层的沉积物中均有分离出,而其他分离到的属于Proteobacteria 的物种,如Citromicrobium bathyomarinum,Martelella mediterranea,Nitratireductor aquibiodomus,Pelagerythrobacter marinus,Qipengyuania vulgaris则大多来自于更深层的环境中。分离得到隶属于Bacillota 的菌株则明显分为两大类群,其中属于Bacillaceae 的菌株除了181E02、475E01 之外,均来自于较上层的沉积物(≤110 cmbsf);而Planococcaceae(葡萄球菌科)的菌株则全部来自于深层的沉积物(≥248 cmbsf)。此外,我们还在334 cmbsf层位处分离到唯一一株属于Bacteroidota 的菌株334S03,它与亲缘模式菌株Muricauda ruestringensisDSM 13 258T的16S rRNA 基因序列相似性为98.48%,为潜在的新物种。
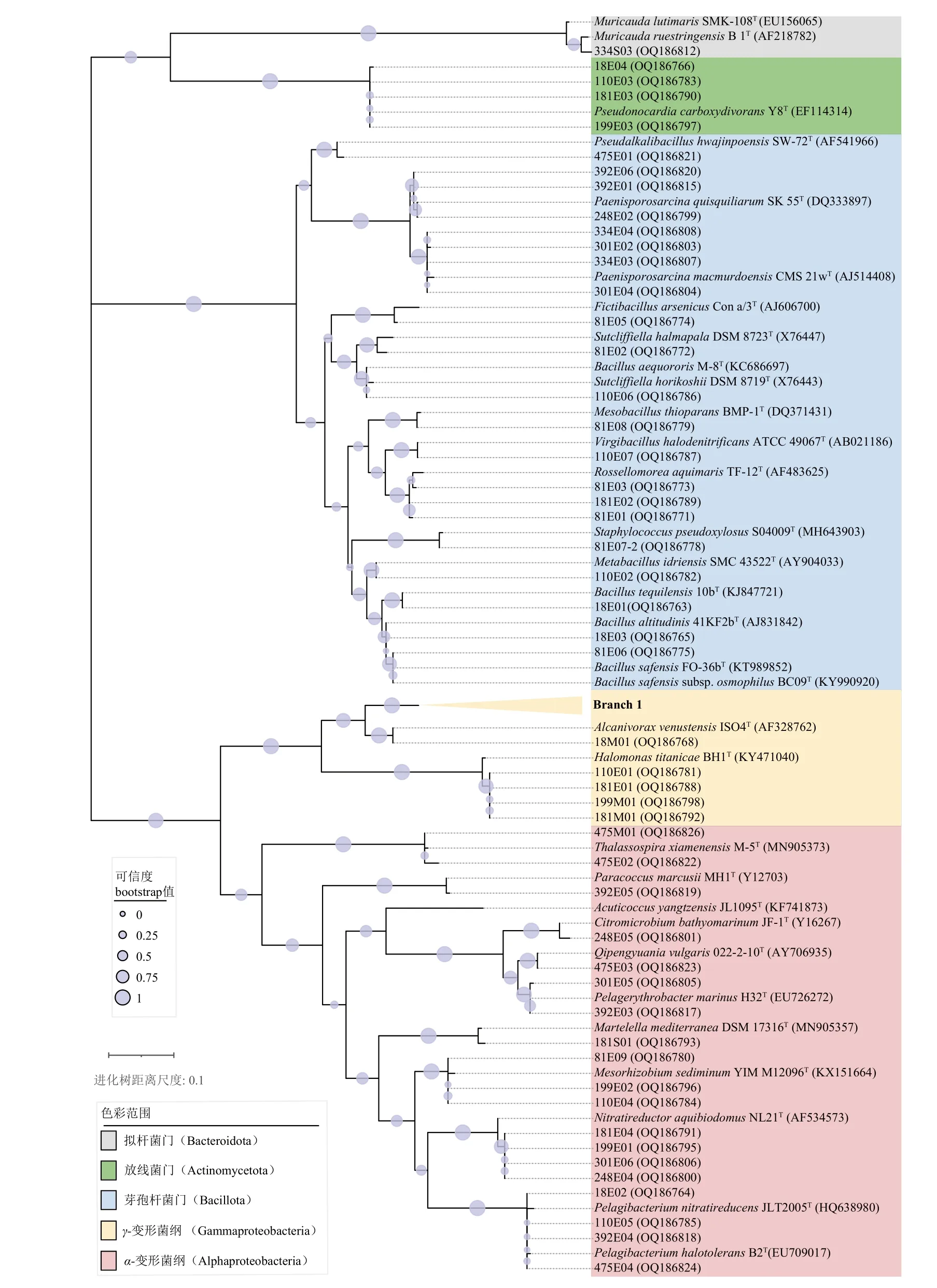
图2 基于16S rRNA 基因序列构建的可培养细菌的系统发育树分支1 中包含的菌株有18A01、18E05、18S01、81A02、81E07-1、199A01、301A01、334E05、334M01、334S02、334S04、392A01、392E02、475E05 和Alcanivorax xenomutans JC109T (HE601937)。Fig.2 Phylogenetic tree of cultivable bacteria isolated from hydrothermal field sediment core in the southern Okinawa Trough based on the 16S rRNA gene sequences using the maximum-likelihood algorithmGenBank accession numbers are shown in parentheses. Bar, 0.1 substitutions per nucleotide position. Branch 1 represented 18A01, 18E05, 18S01, 81A02,81E07-1, 199A01, 301A01, 334E05, 334M01, 334S02, 334S04, 392A01, 392E02, 475E05, and Alcanivorax xenomutans JC109T (HE601937).
2.3 基于16S rRNA 基因高通量测序分析的微生物多样性
所有样品的测序深度指数(Good’s Coverage)覆盖范围在0.999 1 至0.999 9 之间,表明测得的序列代表了研究样品中的绝大多数微生物类群。所有样品的菌群丰富度指数(Chao)、多样性指数(Shannon和Simpson)以及发育树谱系多样性指数(PD whole tree)见图3。可见,18 cmbsf 的样品表现出最大Chao值,为461.62,而475 cmbsf 的样品,其Chao 值在所有样品中最低,为165.09。样品的Shannon 和Simpson指数范围分别为2.13~5.46 和0.50~0.91,它们的最大值出现在100 cmbsf 的样品中,最小值出现在380 cmbsf 的样品中。结合PD whole tree 指数分析,不难发现浅层的样品体现出更大的微生物多样性,且随着样品深度的增加,沉积物样品中微生物的多样性呈现出减少的趋势。
物种聚类分析结果显示(图4),在门水平上,所有样品中Proteobacteria 均占据了主要的微生物类群,其占比69.0%~95.4%。Proteobacteria 的最大丰度出现在380 cmbsf 的层位,而最小丰度出现在100 cmbsf 的层位样品中。样品中检测到的其他主要门类还有Bacteroidota 和Bacillota,但它们的相对丰度在所有样品中均小于12%。Bacteroidota 的相对丰度呈现出随着沉积物深度增加而不断减小的趋势,尤其在240~248 cmbsf 的层位处表现为断崖式的下降,由240 cmbsf 处的7.3%下降到248 cmbsf 处的2.4%。在纲水平上,Proteobacteria 主要分为两个主要类群,其分别为Alphaproteobacteria 和Gammaproteobacteria,且它们在不同层位的样品中分布不尽相同;在浅层次(18~110 cmbsf)的沉积物中Alphaproteobacteria 比Gammaproteobacteria 更多,而随着沉积物深度的增加,Gammaproteobacteria 相对丰度有明显的增加(从18 cmbsf 处的9.0%增加到>248 cmbsf处的55.7%~73.3%),同时Alphaproteobacteria 明显减少(从18 cmbsf 处的63.5%减少到>248 cmbsf 处的18.3%~32.2%),这个现象在240~248 cmbsf界面也表现得尤为突出,分析数值显示240 cmbsf处Alphaproteobacteria 和Gammaproteobacteria 相对丰度分别为21.0%和45.9%, 而248 cmbsf 处的Alphaproteobacteria 和Gammaproteobacteria 相对丰度分别为71.4%和20.3%。在目水平上,属于Alphaproteobacteria 的未培养类群的SAR11 分支和属于 Gammaproteobacteria 的 Ectothiorhodospirales(外硫红螺旋菌目)分别占据了主导地位,在不同层位的沉积物样品中,它们的丰度变化主导了更高级分类单元上物种相对丰度的变化。在科水平上,可以看出高丰度水平的 Alphaproteobacteria 和Gammaproteobacteria 实际上是由高比例的SAR11-Clade I 和Thioalkalispiraceae(硫碱螺旋菌科)引起的。此外,科水平的细菌组成随着沉积物深度的不同体现出更高的多样性,比如属于Campylobaterota( 弯曲菌门) 的Sulfurvaceae 相对丰度在浅层(18~110 cmbsf)的沉积物中随着深度而增加,而在248 cmbsf 的层位及更深层处又急剧减少;在100 cmbsf 层位处的样品中,SAR11-Clade III 占据了整体群落比例的14.2%,而其他层位中SAR11-Clade III 的比例均小于1%;在475 cmbsf 层位的样品中,除了Thioalkalispiraceae 以外,Pseudomonadaceae(假单胞菌科) 和Alcanivoracaceae 同样成为构成Gammaproteobacteria 的主要分类单元,它们的相对丰度值分别达到了16.6%和7.6%,而在其他所有样品中二者的占比均低于5%。
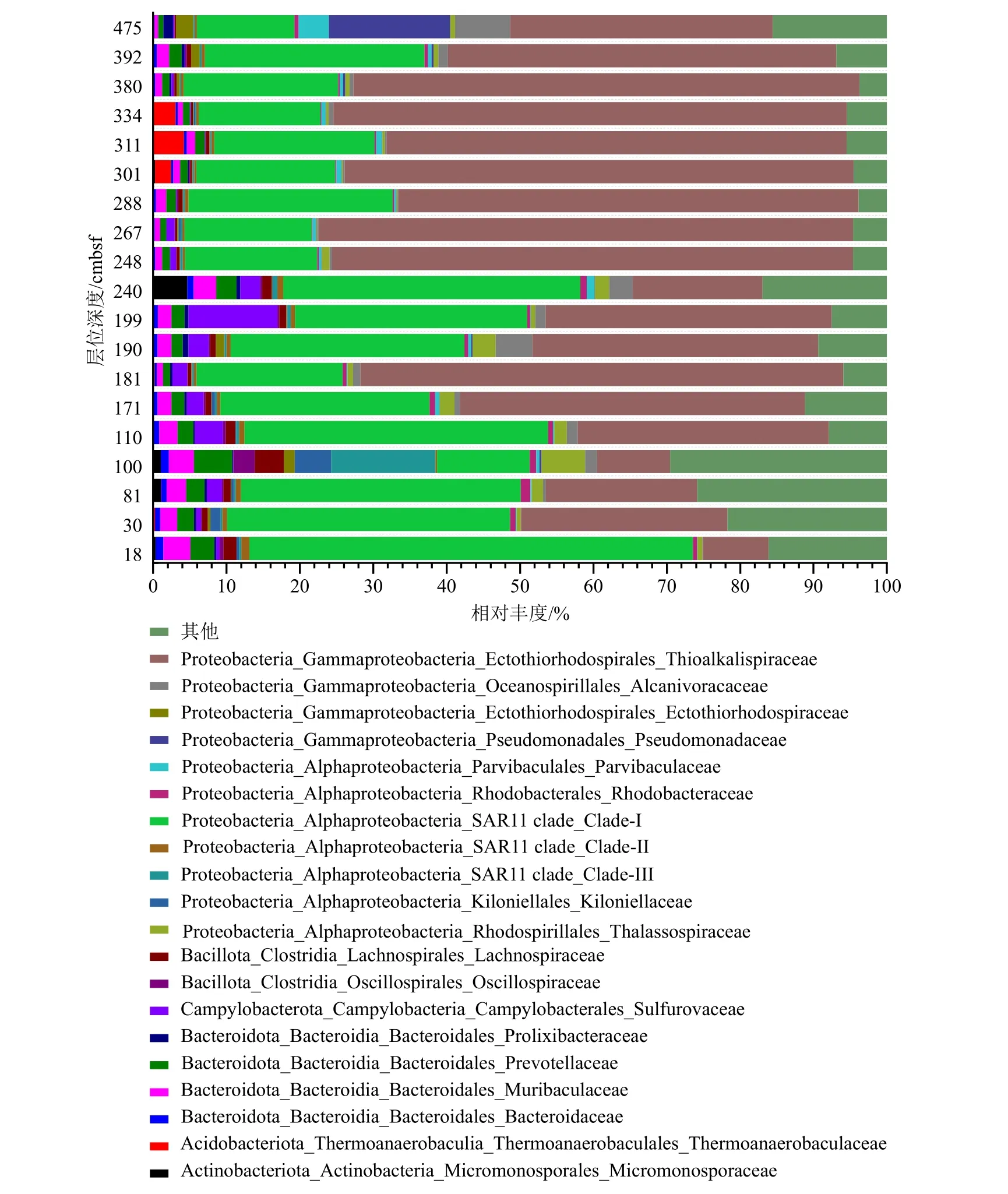
图4 基于科水平的物种聚类显示的样品中不同细菌所占的比例Fig.4 Relative abundance of bacteria in the samples on family level
为了更好地阐明不同微生物类群在不同生境之间的分布规律及差异性,以16S rRNA 基因序列测序获得的相对丰度值,通过Bary Curtis 矩阵法得到了PCoA 图(图5);以科水平ANOVA 差异统计获得的差异物种的相对丰度值绘制了聚类热图(Cluster heatmap)(图6)。结果表明,浅层(18~81 cmbsf)沉积物和深层沉积物(380~475 cmbsf)的样品有着显著的分布差异,而它们自身的微生物组成则是比较接近的。171~311 cmbsf 层位的样品和上述两者都有较明显的差异,通过聚类分析也能与浅层和深层的样品区别开来,自身形成一个较大的分支。在科水平上,深层沉积物尤其是475 cmbsf层位的样品与其他样品相比体现出了巨大的物种差异性。这些都说明,尽管来自于同一个位点,微生物群落在垂直水平上的分布是有明显差异的,同时也反映了冲绳海槽热液区中的沉积物是一个相对复杂的生态系统。

图6 基于科水平差异物种相对丰度的样本聚类热图Fig.6 Cluster heatmap based on relative abundance of differential species at family level
2.4 微生物群落、环境因子和样品间的关系
选取TC、TN、TOC、Cu、Zn、Pb 这6 个典型的环境因子,选取以科水平相对丰度前10 的微生物,通过RDA 分析展示了冲绳海槽南部热液区附近柱状沉积物中微生物群落分布、不同层位样品以及环境因子三者之间的关系。根据图7 发现,在环境因子之间, 柱状沉积物中的Cu、 Zn、 Pb、 TN 与TOC 呈现出正相关。而TOC 对研究样品中微生物群落组成的影响最强,同时其他几个环境因子对微生物的群落分布也具有显著影响且影响力相当。具体来说,未培养的微生物类群SAR11-Clade I 与TOC 含量,Thioalkalispiraceae 与TC 含量之间表现出明显的正相关;Sulfurovaceae 与TOC 等环境因子之间也表现出较强的正相关关系;但是在较浅层样品中相对丰度较高的Prevotellaceae(普雷沃氏菌科)、Muribaculaceae、Lachnospiraceae(毛螺旋菌科)等几个类群的丰富度与参与分析的环境因子之间表现出显著的负相关。从各层位样品在图中的分布还可以看出,浅层沉积物样品和深层沉积物样品分别分布在相对独立的区域,意味着深度接近的沉积物表征出相似的环境因子参数,其对微生物群落分布的影响程度也比较近似。

图7 样品、微生物种群及环境因子RDA 图Fig.7 Redundancy analysis on the relationships between environmental factors and relative abundance at family level
3 讨论
3.1 沉积物柱的元素地球化学特征
在本研究中结合前人的研究数据,166 cmbsf 和444 cmbsf 层位的沉积物样品中Cu、Zn、Pb 元素富集,其含量明显高于正常沉积物中的含量。具体来说,在沉积物166 cmbsf 处,Cu、Zn 和Pb 的含量分别为73.7×10-6、356.0×10-6、160.0×10-6;在444 cmbsf处含量略有下降,分别为38.8×10-6、321.0×10-6、121.0×10-6,均明显高于冲绳海槽中部非热液区沉积物中的元素值(Cu 为23×10-6,Zn 为91×10-6,Pb 为18×10-6)[41]和冲绳海槽北端沉积物中的元素值[42],指示其沉积物中除了有海槽正常沉积物之外,可能还有其他来源含金属物质组分的加入,且杨娅敏等[28]对该柱状沉积物样品的研究也表明,HOBAB4-S2 柱状沉积物中存在热液活动事件的记录。
Wang 等[18]研究冲绳海槽北部柱状沉积物样品的TC 含量,其均显著高于南部表层沉积物样品的TC 含量,而其TN 和TOC 含量则在所有沉积物样品中无显著差异,且与我们测得的数据结果一致。有研究报道,在富含有机质的地区如秘鲁海盆和秘鲁海沟处,其沉积物的总有机碳(TOC)含量为2%~8%,而冲绳海槽的沉积物,其总有机碳的含量为0.66%~1.05%,比富有机质的沉积物中有机碳的含量低很多[43],表明研究区属于贫有机质的区域。同时根据前人的研究可以发现,环境中细菌的群落组成结构在不同TOC 含量的环境中表现出很大差异[18,44],而本研究中,从图7 可以看出冲绳海槽南部热液区附近柱状沉积物中细菌群落结构的最重要变量是TOC,跟之前的研究结果表现一致。同时,本研究的微生物群落结构分析和RDA 分析中可以看出由SAR11-Clade I 主导的Alphaproteobacteria 类群和由Thioalkalispiraceae 主导的Gammaproteobacteria整体上来看与TOC 和TC 含量都有一定的正相关关系。但是由于TOC 对Alphaproteobacteria 的影响程度大于Gammaproteobacteria,而TC 对Gammaproteobacteria 的影响程度大于Alphaproteobacteria,造成了从浅层到深层 Gammaproteobacteria 增多而Alphaproteobacteria 减少的趋势,这与Teske[45]之前对海洋沉积物中微生物的调查研究结果相近。因此,柱状沉积物中的TOC 和TC 含量会对该区域微生物的多样性和群落结构特征产生重要的影响。尽管其中原因和涉及的微生物代谢机制可能很复杂且需要进一步的深入研究,但我们很容易得出这样的推测:TC 和TOC 作为微生物重要的营养物质和能量来源,微生物的代谢过程会因其所处环境中的含量不同而产生一定的偏好性,从而引起主导微生物类群的差异。
3.2 沉积物柱中的微生物多样性
在我们利用纯培养手段分离的64 个菌株中,Proteobacteria 在门分类水平上最丰富。据报道Proteobacteria 通常作为常见的栖息菌群存在于不同的热液环境中, 比如 Perez Rodriguez 等和Rajasabapathy 等[46-47]分别在不同热液区分离鉴定到了大量的Gammaproteobacteria。同时,有研究表明一些属于Proteobacteria 的Alcanivorax、Thalassospira、Halomonas、Paracoccus可能参与到产酸或产碱的硫氧化过程[11],本研究分离的菌株中也有相当的比例属于这些属,而这些属的菌株在其他热液环境中也曾分离到[47-48]。另外有研究表明,属于Gammaproteobacteria 的Marinobacter和Pseudoalteromonas的微生物类群在很大程度上参与了金属氧化[49],冲绳海槽中部热液区的沉积物中也分离到了大量的Marinobacter和Pseudoalteromonas属的菌株,但是遗憾的是本研究并没有分离到这两个属的菌株,尽管高通量测序数据表明它们在样品中是存在的。除了Proteobacteria 之外,Bacillota 是分离得到菌株中第二大门类的类群,这类细菌由于其独特的细胞结构和生理抗逆性可以耐受的温度范围很广,以不同的表型存在于不同的热液环境中[50]。研究人员曾从卡斯特罗海山热液区分离了15 种属于Bacillus的菌株,而铁锰等金属元素对它们的生长有明显的促进作用[51],同时分离自Guaymas 盆地的一些属于Bacillus的菌株参与了铜介导的锰氧化过程[52]。因此,我们推测本研究样品分离的属于Bacillota 的菌株也可能参与了周边生态系统中的金属氧化过程。
基于16S rDNA 高通量测序的微生物组成分析显示,本研究中沉积物样品中的微生物组成在门水平上,Proteobacteria 占据绝对的优势地位。在这个门中,Gammaproteobacteria、Alphaproteobacteria 是两个主要的类群,这一结果与冲绳海槽中部Iheya North 和Iheya Ridge 热液区以及太平洋富含金属的热液喷口附近沉积物中微生物组成的报道类似[21,53-55]。先前的研究表明Epsilonproteobacteria(ε-变形菌纲)在与热液喷口流体和烟囱结构相关的微生物栖息地中占主导地位,同时也是从活跃的热液喷口中鉴定出的优势细菌,在碳、硫的循环中发挥重要作用[56-57],而且在冲绳海槽的部分热液喷口处也发现Epsilonproteobacteria 占据了一定丰度比例[16,18-20]。然而,在本研究的样品中Epsilonproteobacteria 的丰度是极低的。另一方面,关于冲绳海槽非热液区沉积物中微生物的群落研究表明, Deltaproteobacteria(δ-变形菌纲)和Chloroflexi(绿弯菌门)通常为主要类群[18,58],而Deltaproteobacteria 和Chloroflexi的相对丰度在本研究的样品中也是非常低的(<1%)。通过综合比较,我们发现尽管在海洋热液区沉积环境中,Proteobacteria 通常作为主导的微生物类群,但是上述的研究内容支持了这样一种假设,由于海底热液的活跃程度不同以及与热液喷口的距离不同,不同位点的沉积物样品中微生物组成受热液活动的影响程度不同,从而造成了其微生物主要类群的差异。比如Epsilonproteobacteria 看起来更喜欢活跃的热液喷口流体和烟囱体周围的高温环境; Alphaproteobacteria 和 Gammaproteobacteria更喜欢不活跃的热液栖息地。进一步推测可以看出,HOBAB4-S2 柱状沉积物样品由于其具有高丰度的Alphaproteobacteria、Gammaproteobacteria 和低丰度的Epsilonproteobacteria,与冲绳海槽活跃的热液喷口和非热液区沉积物中微生物群落均有较大差异,表明尽管该位点附近的热液喷口可能是不活跃的,但是其沉积物中微生物群落组成还是受周边热液活动影响的。
3.3 柱状沉积物中微生物多样性的影响因素
由于热带西太平洋地理位置和水文条件的特殊,细菌群落的多样性很容易受到洋流的影响[59],这是一个非常复杂和动态的过程,主要涉及到沉积物的来源、盐度、有机质的生物利用率、pH 以及其他的地球化学和物理化学因素[60]。本研究的区域位于冲绳海槽南部热液区附近,其沉积和物质输运除了受热液活动影响,也在一定程度上受洋流[61]因素的影响。冗余分析结果显示相比于其他测定的环境因子,柱状沉积物中TOC、Zn 和Pb 的含量与微生物的群落组成有着更为密切的关系,因此环境特征对于特定环境下微生物的群落结构具有非常重要的影响。从我们研究的结果中发现,微生物的群落结构随沉积物深度的不同而不同,这可能是地球化学环境的一个反映,也可能决定了微生物的种类、数量和多样性分布。另外,不同的沉积事件、沉积过程也可能形成不同的微生物群落。因此推测微生物的群落结构和组成在一定程度上跟沉积事件的干扰和热液活动事件有关,而这一点在西菲律宾海盆和西太平洋暖池的研究中已经证实[59,62-63]。
4 结论
(1)冲绳海槽南部HOBAB4-S2 柱状沉积物主要由SiO2、Al2O3、Fe2O3等组成,这3 种组分含量为3.8%~69.3%。该位点的沉积物有机质含量较为贫乏,同时有两个富含Cu-Zn-Pb 的层。
(2)利用HOBAB4-S2 柱状沉积物,通过分离培养得到纯菌株64 株,Proteobacteria 约占总数的59%。基于16S rRNA 基因高通量测序的结果同样表明,在所有沉积物层位中变形菌门为占据绝对优势的类群,这种现象在更深层(>240 cmbsf)的沉积物中表现的更为明显;相对于深层的沉积物,表层沉积物表现出更高的微生物多样性。通过比较可培养和免培养方法获得的不同微生物群落多样性表明,该位点沉积物中存在大量有活力但不可培养的微生物。
(3)相比于其他测定的环境因子,柱状沉积物的TOC 含量与其微生物的群落组成有着更为密切的关系。
总的来说,本研究揭示了冲绳海槽南部热液区附近沉积环境中细菌的多样性和垂直分布特征,为微生物的群落结构及其与地质沉积过程的相关性提供了证据。随着现代分离培养技术和组学技术的快速发展,通过个性化分离培养手段培养难培养的微生物并从分子水平上开展研究,同时结合高通量测序的组学分析,将有助于进一步地理解海底热液环境中微生物的代谢机制及其与环境的协同演化过程以及它们在全球物质循环过程中的作用。
猜你喜欢
VOGUE服饰与美容(2023年5期)2023-05-10
高技术通讯(2021年6期)2021-07-28
石油与天然气地质(2021年3期)2021-06-29
高校地质学报(2020年6期)2021-01-11
环球时报(2017-05-16)2017-05-16
航空知识(2017年2期)2017-03-17
新疆地质(2016年4期)2016-02-28
应用海洋学学报(2015年1期)2015-11-22
江西煤炭科技(2015年1期)2015-11-07
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28
