
黑果枸杞XTH转录因子家族鉴定与生物信息学分析
2023-11-02 08:47:26贾西贝李效雄胡晓桐马彦军
草地学报 2023年10期
贾西贝,李效雄,杜 雨,胡晓桐,刘 筠,马彦军*
(1.甘肃农业大学林学院,甘肃 兰州 730070;2.兰州资源环境职业技术大学,甘肃 兰州 730030)
自然界中有许多不利因素影响着植物的生长发育过程,这些因素主要分为生物胁迫和非生物胁迫两大类。而盐胁迫和干旱胁迫是非生物胁迫中影响植物生长发育的主要原因,对植物的遗传潜力存在较大的影响[1]。盐胁迫会对植物细胞渗透产生影响,引发离子毒害,从而诱发植株内活性氧的产生,不仅导致各组织细胞内的离子失衡,还会使植物发生生理性干旱,缺少生长发育必需的营养元素等许多问题[2-3]。面对盐胁迫,多数植物通过自身合成能够渗透调节的物质,或是通过清除活性氧等方法,增强自身应对盐胁迫时的调节能力,从而减小盐胁迫对其生长发育带来的危害与影响[4]。此外,干旱也可以迫使植物的生理生化指标发生异常[5],引起细胞壁的机械损伤、叶片的光合效率降低或气孔闭合,以及植物细胞脱水等许多问题,最终导致植株的代谢紊乱[4]。由于盐胁迫或干旱胁迫导致的植物细胞脱水,会使其细胞壁的伸展性受到直接影响。面对此类问题,植物会对自身细胞大小进行调节,或是通过改变细胞壁的延展性来保障细胞膨压,从而将细胞失水的影响降到最低,适应失水条件[6]。因此,明确植物的耐盐耐旱等抗逆胁迫机制,对提高植物的育种及栽培技术有着重要意义[7]。
木葡聚糖内糖基转移酶/水解酶(Xyloglucan endotransglucosylase/hydrolase,XTH)是一种细胞壁操作酶,属于糖苷水解酶16(GH16)家族[8],具有两种不同的催化活性,一种为水解酶(Xyloglucan endohydrolase,XEH),另一种为转糖基酶(Xyloglucan endotransglucosylase,XET)。XTH酶多数的保守基序都是DEIDFEFLG,该基序被认为是XET和XEH活性的催化位点[9]。根据结构的不同可将XTH蛋白分为I/II和III组,而III组分为两个亚组IIIA和IIIB[10]。研究表明,具有糖基转移酶活性的XTH主要属于I/II组,而具有水解酶活性的XTH主要属于III组。XTH蛋白中有两个保守结构域,名为Glyco_hydro_16和XET_C。XET(C-XET)的C端结构域是区分XTH成员和其他GH16家族的主要依据[11-12]。在植物的生长发育及整个生命周期中,XTH基因参与植物许多抗逆性调控过程,具有极其重要的作用。例如,在辣椒(Capsicumannuum)中,CaXTH1,CaXTH2以及CaXTH3盐胁迫处理后表达均呈现升高趋势,同时CaXTH3过表达有利于番茄(Solanumlycopersicum)和拟南芥(Arabidopsisthaliana)耐盐能力的提升[13-14]。在较高浓度的NaCl胁迫下,蒺藜苜蓿(Medicagotruncatula)的MtXTH3基因表达显著上调[15]。柿子(DiospyroskakiThunb)中DkXTH1基因的过表达有利于增强拟南芥的抗盐胁迫、抗干旱胁迫以及对脱落酸(Abscisic acid,ABA)的耐受能力[16]。上述研究表明XTH基因家族在植物耐盐和抗干旱等非生物胁迫的生理过程中发挥一定的重要作用。
黑果枸杞(LyciumruthenicumMurr.)属于茄科(Solanceae)枸杞属(LyciumL.),多棘刺灌木,无主枝,多分枝,主要分布在西北地区的戈壁滩或盐碱地中[17]。黑果枸杞生命力顽强,耐寒耐旱、抗盐碱,是我国防风固沙、改良土壤荒漠化的先锋植物[18],同时因其果实中丰富的营养物质价值较高,因此也是荒漠地区主要的经济物种[19-20]。黑果枸杞虽然是西北地区重要的耐盐植物之一,但对其在分子层面对逆境胁迫的研究还较少。当前对植物抗逆性的研究多集中在模式植物上,如拟南芥、水稻(Oryzasativa)和烟草(Nicotianatabacum)[21-22]等。因此,本研究通过分析盐胁迫下黑果枸杞的转录组数据库,对黑果枸杞XTH家族成员进行了系统的鉴定及生物信息学分析,为更加深入了解黑果枸杞响应胁迫机制及XTH家族成员功能的进一步研究提供理论依据和参考。
1 材料与方法
1.1 试验材料
本试验材料为甘肃农业大学林学院组培室枸杞组培苗。材料生长环境为:昼夜温度控制在(22±2)℃/(20±2)℃的范围内,光照每天保持16 h,相对湿度保持在65%,光照强度为700 μmol·m-2·s-1。1/2MS培养基配方主要包括:1/2MS培养基2.47 g·L-1,蔗糖20 g·L-1,琼脂5 g·L-1,NAA 0.2 mg·L-1,IBA 0.2 mg·L-1。2周后,取长势良好、条件相同的实验苗进行炼苗,经过5天后移栽培养至1/2 Hoagland营养液中,培养液(pH=5.7)的配制成分包括:2 mmol·L-1KNO3,0.5 mmol·L-1MgSO4·7H2O,10 μmol·L-1MnCl2·4H2O,0.5 mmol·L-1Ca(NO3)2·4H2O,0.5 mmol·L-1KH2PO4,0.05 μmol·L-1Na2MoO4·2H2O,1.6 μmol·L-1ZnSO4·7H2O,0.6 μmol·L-1CuSO4,50 μmol·L-1H3BO3,0.06 μmol·L-1Fe-citrate·2H2O[23]。在营养液中培养14天,然后分别用50 mmol·L-1和250 mmol·L-1的NaCl进行盐胁迫处理。在加入NaCl的0 h,1 h和12 h三个时间段分别取样,并设置空白对照组。以相同的处理方式用样品做3次重复。之后将样品材料的根、茎、叶各采集0.1 g,液氮速冻,并在-80℃的条件下冷藏备用。
1.2 方法
1.2.1盐胁迫下黑果枸杞XTH基因家族的筛选及理化性质分析 对数据库NR,KOG和Swiss-Prot分别注释,将筛选出的候选基因蛋白序列导入NCBI(https://www.ncbi.nlm.nih.gov/orffinder/)进行预测,利用在线软件Pfam(https://Pfam.xfam.org/)以及NCBI的CDD(https://www.ncbi.nlm.nih.gov/cdd)进行保守结构域分析,然后利用InterProScan(v5.17-56.0)进行进一步的验证,剔除不含保守结构域的序列,剩余序列即为黑果枸杞的XTH蛋白序列。使用ExPASy(http://web.expasy.org/protparam/)对XTH转录因子进行预测,分析其氨基酸的分子量、脂肪系数、不稳定数以及等电点等相关数据。
1.2.2盐胁迫下黑果枸杞XTH基因家族保守域验证与基序分析 通过开放阅读框(Open reading frame,ORF)(https://www.ncbi.nlm.nih.gov/orffinder)对LrXTHs的氨基酸序列进行初步预测。LrXTHs氨基酸序列的比对则使用DNAMAN 8.0软件。用在线软件MEME(https://meme.nbcr.net./meme/tools/meme)对XTH基因的保守域单元(Motif)进行检测,其中数目最大的保守基序为5,设其余参数值为默认。
1.2.3盐胁迫下黑果枸杞XTH系统发育和表达谱分析 从TAIR(http://www.arabidopsis.org/)、UniPort(https://www.uniprot.org)和NCBI上获取拟南芥、水稻、番茄、烟草和葡萄(Vitisvinifera)五种植物的序列。使用MEGA-X软件(版本为10.0.2),采用极大似然法(ML,Piosson model),bootstrap1000构建黑果枸杞、拟南芥、水稻、番茄、烟草和葡萄的系统发育树。构建的进化树则通过在线软件iTOL(http://itol.embl.de/)进行美化。利用软件TBtools内的Heatmapper程序构建黑果枸杞叶部和根部组织的基因表达谱。
2 结果与分析
2.1 基因筛选
通过Pfam和NCBI Blast,筛选含有PF00722(Glycohydro_16)和PF06955(XET_C)保守结构域的蛋白序列,共筛选得到19个LrXTHs基因家族成员,并命名为LrXTH1~LrXTH19(表1)。使用在线软件ExPASy预测分析19个LrXTHs氨基酸序列(表1),结果显示氨基酸数目介于258~330 aa之间;分子量在29.770 62~37.980 83 kDa之间;理论等电点介于4.98~8.92之间,LrXTHs的基因长度在954 bp~1 814 bp之间。其中稳定蛋白有11个,不稳定蛋白有8个,通过脂肪系数可知19个LrXTHs蛋白属于亲水性蛋白。

表1 黑果枸杞XTH转录因子的理化性质Table 1 The physicochemical properties of XTH transcription factors in Lycium ruthenicum
2.2 保守域验证与基序分析
通过对19个LrXTHs蛋白序列进行多序列比对,结果表明(图1):在LrXTHs蛋白中代表催化活性残基的活性位点(ExDxE)高度保守,且所有LrXTHs蛋白都具有DE(I/L/F/V)D(F/I/M)EFLG保守结构域。DE(I/L/F/V)D(F/I/M)EFLG结构域氨基酸数量在103~160个之间,氨基酸序列中多个氨基酸位点都是稳定不变的,为DEXDXEFLG(图1)。19个LrXTHs蛋白的催化残基附近具有潜在的N-连接糖基化位点:N(Y/H)V(R/L/T/I/S/K/E/A)T(S/K/R/P)G(A/D/R)。
将黑果枸杞XTH导入MEME软件,对其家族成员的保守序列验证分析。结果显示,得到5个LrXTH基因家族的保守基序,并为其从Motif 1依次命名至Motif 5(图2),长度在21~50 aa之间,且LrXTH中都含Motif 1~Motif 5的基序。这5个保守基序通过Intreproscan(v5.17-56.0)工具功能注释(表2),发现Motif 1~Motif 4为Glyco_hydro_16,Motif 5为XET-C。

图1 黑果枸杞XTH转录因子的多序列对比Fig.1 Multi-sequence comparison of XTH transcription factors in L. ruthenicum

图2 黑果枸杞XTH转录因子的蛋白保守基序Fig.2 The protein conserved motif of LrXTH transcription factor in L. ruthenicum
2.3 系统进化树分析
为进一步解析LrXTH基因家族的系统发育关系,选择19个黑果枸杞的XTHs成员,33个拟南芥的XTHs成员、29个水稻的XTHs成员、32个番茄的XTHs成员、56个烟草的XTHs成员和34个葡萄的XTHs成员构建系统发育树。结果显示,五种植物的XTHs基因家族成员分为4个亚族(图3),除III A和Ancestral亚族不含有LrXTH成员外,III B亚族含有基因LrXTH2,LrXTH13和LrXTH14,其余成员则位于I/II亚族。同时I/II亚族的XTH蛋白数量较多,共有16名成员,AtXTH1,AtXTH2,AtXTH3和AtXTH11基因则被归入Ancestral亚族。同时在系统发育树中部分XTH基因形成了相关姐妹对,如LrXTH14和NtXTH47,LrXTH1和NP_001 234369.2,LrXTH3和NtXTH28,NtXTH28,LrXTH4和XP_004250701.1等(图3)。总而言之,XTH基因家族成员在不同植物进化过程有所差异,并且在这五种植物中黑果枸杞与番茄的家族成员关系更加密切。
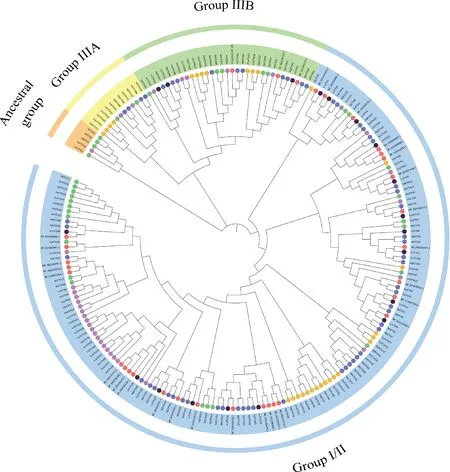
图3 拟南芥、水稻、烟草、葡萄、番茄和黑果枸杞中LrXTH基因的系统发育进化树Fig.3 Phylogenetic tree of LrXTH genes in Arabidopsis thaliana,Oryza sativa,Nicotiana tabacum,Vitis vinifera,Solanum lycopersicum and L. ruthenicum
2.4 基因的表达谱分析
为初步了解LrXTH基因的功能,在NaCl处理下对黑果枸杞叶部和根部组织的19个LrXTHs基因表达进行分析。结果显示,LrXTH基因家族成员在黑果枸杞不同组织中的表达量存在明显差异。在盐胁迫下黑果枸杞根部组织的多数XTHs表达量较高,其中LrXTH3,LrXTH9,LrXTH18和LrXTH19是受盐胁迫诱导而表达量上调的基因。在250 mmol·L-1浓度的盐处理下,根部组织中LrXTH3和LrXTH19基因分别在1 h和12 h时(R250_1和R250_12)表达量显著上调;同一时间内(1 h),LrXTH9表达量在50 mmol·L-1的NaCl处理下显著上调,LrXTH18表达量在250 mmol·L-1的NaCl处理下显著上调,推测这4个基因可能参与盐胁迫反应并发挥功能。在盐胁迫下,黑果枸杞叶部组织的LrXTH9表达量在50 mmol·L-1的NaCl处理下随盐处理时间的增加而升高,其属于盐胁迫诱导上调基因。盐胁迫处理下LrXTH5和LrXTH8表达量在根部和叶部组织中均表现出较明显的下降趋势,其中LrXTH5在1 h和12 h两组处理时间内,其表达量随NaCl浓度的升高而降低;在浓度为50 mmol·L-1和250 mmol·L-1的两组NaCl处理下,其表达量随时间的增加而降低,它属于盐胁迫诱导下调基因。在叶片和根部中,LrXTH9表达量在1 h和12 h时,50 mmol·L-1的NaCl处理下表现为上调,250 mmol·L-1的NaCl处理下表现为下调(图4)。通过与对照组比较,推测LrXTH3,LrXTH9,LrXTH18和LrXTH19在黑果枸杞抗盐胁迫的生理过程中起到了重要的作用。

图4 LrXTH基因在不同组织中的表达模式Fig.4 Expression patterns of LrXTH genes in different tissues注:L表示叶;R表示根.R50-1表示50 mmol·L-1NaCl处理下1小时所取根样;L50-1表示50 mmol·L-1 NaCl处理下1小时所取叶样;R250-1表示250 mmol·L-1 NaCl处理下1小时所取根样;L250-1表示250 mmol·L-1NaCl处理下1小时所取叶样;R50-12表示50 mmol·L-1 NaCl处理下12小时所取根样;L50-12表示50 mmol·L-1NaCl处理下12小时所取叶样;R250-12表示250 mmol·L-1NaCl处理下12小时所取根样;L250-12表示250 mmol·L-1NaCl处理下12小时所取叶样,R0表示CK根;L0表示CK叶。下同Note:L denotes the leaf,R the root.R50-1:root sample under 50 mmol·L-1 of NaCl treatment for 1 hour;L50-1:leaf sample under 50 mmol·L-1 of NaCl treatment for 1 hour;R250-1:root sample under 250 mmol·L-1 of NaCl treatment for 1 hour;L250-1:leaf sample under 250 mmol·L-1 of NaCl treatment for 1 hour;R50-12:root sample under 50 mmol·L-1 of NaCl treatment for 12 hours;L50-12:leaf sample under 50 mmol·L-1 of NaCl treatment for 12 hours;R250-12:root sample under 250 mmol·L-1 of NaCl treatment for 12 hours L250-12:leaf sample under 250 mmol·L-1 of NaCl treatment for 12 hours;R0:root sample under root control treatment;L0:leaf sample under leaf control treatment.The same as below

图5 LrXTH基因响应盐处理的表达分析Fig.5 Expression analysis of LrXTH genes in response to salt treatments
3 讨论
对XTH基因的鉴定有助于了解植物面对耐盐和抗旱等环境胁迫时耐受性基因的表达和调控机制。已有研究通过转基因技术证实了XTH基因在提高机体对盐胁迫耐受性中的重要作用[24-25]。目前,XTH基因家族已经在多个植物中得到鉴定,其中包括拟南芥33个[26],水稻29个[27],大麦(HordeumvulgareL.)35个[28]和烟草56个[22]等。本研究基于黑果枸杞转录组数据鉴定出19个LrXTHs家族成员,并对其进行系统地生物信息学分析及功能预测。
验证保守结构域是研究基因家族及其功能的重要内容。通过多序列比对及保守基序的分析,19个LrXTHs成员都含DE(I/L/F/V)D(F/I/M)EFLG保守结构域,这与拟南芥[26]、水稻[27]、盐角草[10]等蛋白序列结构高度相似。与进化树结合分析发现,同一亚族的LrXTHs基因具有较高相似度的保守基序,说明黑果枸杞XTH基因拥有较为保守的蛋白结构。这为LrXTHs各序列之间的保守程度以及对进化树的亚族分类提供了较高保障,从而有助于更深层次地进行黑果枸杞XTH基因家族的功能分析。另外,在同一亚族中,LrXTHs基因的位置相近,Motif数目和排列顺序大致相同,这与凤梨[29](Ananascomosus)和胡杨[30](Populuseuphratica)等植物中XTH转录因子结果相似。这表明LrXTHs的分类不仅有保守基序的保障,而且亚族之间的序列也有着较高的保守性。
对19个LrXTHs进行系统发育分析,划分出4个亚族(图3),这与XTH家族中水稻、盐角草[10](Salicorniaeuropaea)、桃[31](Prunuspersica(L.) Batsch)和辣椒[32]等植物的分类结果一致。除Ancestral亚族外,其它亚族全都含有拟南芥、水稻、烟草及葡萄的XTH基因家族成员,说明黑果枸杞和它们的演化路径具有一定的相似性,通过比较这几种植物XTH基因家族的遗传演化过程,发现与黑果枸杞XTH家族同源程度最高的植物为番茄,表示黑果枸杞XTH家族成员与番茄的进化起源关系是十分相近的。部分研究表明Ⅰ/Ⅱ亚族和ⅢB亚族的XTH蛋白一般具有XET活性[33-35],而ⅢA亚族的XTH蛋白具有XEH活性[36-37]。通常位于同一亚族的基因具有相似的功能,因此可以通过对进化树亚族中基因家族成员的分析,推测相同分支的基因功能[38]。拟南芥XTH19和XTH23受BES1调控,参与侧根的生长发育并促进侧根的盐适应[16],因此推测LrXTH19具有类似的作用;AtXTH30作为负调控因子参与拟南芥对盐胁迫的响应[39],推测同一亚族的LrXTH2,LrXTH13,LrXTH14具有类似的功能;拟南芥的根部在生长发育过程中,AtXTH10起着重要作用,其缺失或过量表达都会对拟南芥幼苗根部生长产生影响[40],推测同分支的LrXTH10对黑果枸杞根部生长有着相同的作用;水稻在干旱胁迫下,OsXTH2表达明显升高并呈现出相关趋势[41],推测同分支的LrXTH1,LrXTH4,LrXTH5在黑果枸杞抗干旱胁迫过程中可能发挥重要作用;在盐胁迫和干旱胁迫下,葡萄的VvXTH3,VvXTH5,VvXTH31,VvXTH34表达量显著增加[42],推测同分支的LrXTH4,LrXTH9,LrXTH10和LrXTH12在黑果枸杞耐盐和抗干旱等非生物胁迫响应过程中发挥重要作用。通过对基因表达谱分析,能够对植物的生物学特性(如抗逆胁迫、生长发育以及亚族组织的特异性等)进行更加深入地探索,为下一步的功能分析打下坚实的数据基础[43-44]。
根据基因表达谱显示,经过盐胁迫处理,多数黑果枸杞LrXTHs基因的表达水平都高于空白对照,同时基因的表达模式在叶片和根部中也存在差异。其中LrXTH3,LrXTH18,LrXTH19仅根部上调表达,这在水稻中也有同样的现象,其中7个XTH基因(OsXTH1,OsXTH2,OsXTH4,OsXTH13,OsXTH15,OsXTH16和OsXTH25)主要在14日龄幼苗的根中表达,而在其他组织中未检测到表达[10]。葡萄的VvXTH20在盐胁迫处理3 h内根部表达量显著升高[42],且同一亚族的LrXTH18在R250_1 NaCl处理下显著升高,推测LrXTH18在黑果枸杞根部的抗盐胁迫过程中起到一定作用;在干旱和盐胁迫下,VvXTH31表达量在葡萄叶部组织中显著上调[42],而同一亚族中LrXTH9在黑果枸杞的叶中表达水平较高,推测LrXTH9在黑果枸杞叶部的抗盐胁迫过程中参与调控过程;同时LrXTH9在低浓度NaCl(L50和R50)处理下,分别在1 h和12 h两组不同时间处理中都表现为表达量上调,推测该基因在黑果枸杞根部和叶部组织中参与耐盐胁迫。在拟南芥中AtXTH14,AtXTH15和AtXTH31基因在盐胁迫处理下显著下调,表明其参与植物抗盐的调控过程[45],因此推测黑果枸杞的LrXTH15和LrXTH16参与对NaCl胁迫抗性的负调控过程。在水稻中OsXTH11在抗盐胁迫中发挥重要作用[46],推测同一亚族分支的LrXTH3,LrXTH18和LrXTH19可能在黑果枸杞抗盐胁迫中发挥重要作用。因此,可将这些LrXTHs作为候选基因为作物抗盐性研究提供参考,但仍需通过相关研究来完善LrXTHs在盐胁迫下的作用机理。
4 结论
本研究对盐胁迫下黑果枸杞转录组数据分析,得到19个LrXTHs家族成员,并对其理化性质、系统进化树以及表达谱模式等进行研究,从而预测其潜在的生物学功能。多序列比对分析表明LrXTHs的保守结构域为DE(I/L/F/V)D(F/I/M)EFLG。系统发育分析将LrXTHs分成4个亚族。表达谱分析表明LrXTH3,LrXTH18和LrXTH19在黑果枸杞根部表达水平较高,而LrXTH9基因在叶部和根部表达水平较高,推测它们可能在黑果枸杞抗盐胁迫过程中具有重要作用。研究结果为黑果枸杞和XTH基因家族潜在功能的进一步研究提供了参考。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
江苏农业科学(2022年17期)2022-09-24 11:42:26
四川大学学报(自然科学版)(2021年4期)2021-07-15 09:10:48
中国农业科学(2020年5期)2020-03-30 02:02:46
河北果树(2020年1期)2020-02-09 12:31:26
生命科学研究(2018年1期)2018-05-29 01:12:47
天然产物研究与开发(2018年3期)2018-05-07 06:38:29
辽宁林业科技(2017年4期)2017-06-22 10:23:36
中成药(2017年3期)2017-05-17 06:09:00
上海农业学报(2017年3期)2017-04-10 12:39:14
