
竹茶混交模式对表层土壤有机碳储量及组分的影响
2023-11-01 04:51:20曹意丁苏雅覃云斌何昕诺马姜明
广西植物 2023年9期
关键词:毛竹林
曹意 丁苏雅 覃云斌 何昕诺 马姜明



摘 要:為探究毛竹林下种植茶树对土壤有机碳储量与碳组分的影响,该研究以毛竹纯林、竹茶混交林和常绿阔叶林为研究对象,采集这3种林分类型的表层(0~10 cm)土壤,测定土壤有机碳(SOC)、碳组分、生物与非生物因素指标。结果表明:(1)竹茶混交林林下植物多样性相较于毛竹纯林显著降低,但其土壤有机碳密度(22.54±2.09) t·hm-2、碳组分与毛竹纯林无显著差异(P>0.05)。竹茶混交林的矿物结合态有机碳(MOC)为(20.13±1.83) g·kg-1,占总有机碳的92.66%。常绿阔叶林土壤有机碳密度比竹茶混交林和毛竹纯林高土壤有机碳密度分别高41.15%和41.00%(P<0.05)。(2)3种林分类型土壤微生物量碳(MBC)含量范围为0.58~3.08 g·kg-1,土壤16S rRNA丰度范围为2.18×1010 ~5.65×1010copies·g-1,固碳基因cbbL丰度范围为0.37×108~ 1.10 ×108 copies·g-1,土壤微生物碳利用效率范围为0.03~0.28;3种林分类型之间微生物相关指标不存在显著差异(P>0.05)。(3)3种林分类型SOC与土壤pH、砂粒含量和地上凋落物生物量呈显著负相关,与土壤黏粒含量、粉粒含量、总氮、C∶N、总磷和铵态氮含量呈显著正相关(P<0.05)。(4)就不同碳组分而言,颗粒有机碳(POC)和MOC均与土壤pH、砂粒含量和根系生物量呈显著负相关,与土壤含水量、黏粒含量、粉粒含量、总氮、C∶N、总磷和铵态氮含量呈显著正相关(P<0.05)。综上表明,竹茶混交改造会造成原生毛竹纯林林下植被多样性下降,但并未造成土壤碳储量下降;而相较于常绿阔叶林,毛竹经营措施需要改进,以提升其碳汇效益。
关键词: 颗粒有机碳, 矿物结合有机碳, 土壤有机碳密度, 毛竹林, 竹茶混交林
中图分类号:Q948
文献标识码:A
文章编号:1000-3142(2023)09-1668-10
收稿日期:2022-07-10
基金项目:广西科技重大专项课题(桂科AA20161002-1); 广西重点研发计划项目(桂科AB21220057); 广西科技基地和人才专项(桂科AD21220163)。
第一作者: 曹意(1996-),硕士研究生,主要从事恢复生态学研究,(E-mail)cywiyii@163.com。
*通信作者:覃云斌,博士,主要从事森林生态学研究,(E-mail)shuibaoqinyunbin@163.com。
Effects of bamboo-tea mixed model on surface soil
organic carbon storage and components
CAO Yi1,2,3, DING Suya1,2,3, QIN Yunbin 1,2,3*, HE Xinnuo1,2,3, MA Jiangming1,2,3
( 1. Key Laboratory of Ecology and Environmental Protection of Rare and Endangered Animals and Plants, Ministry of Education (Guangxi Normal
University), Guilin 541004, Guangxi, China; 2. Guangxi Key Laboratory for Conservation and Sustainable Utilization of Landscape
Resources in Lijiang River Basin, Guilin 541006, Guangxi, China; 3. Institute of Sustainable Development
and Innovation, Guangxi Normal University, Guilin 541006, Guangxi, China )
Abstract:In order to explore the effects of planting tea trees under Phyllostachys edulis forest on soil organic carbon storage and carbon components, we took pure bamboo forest, bamboo-tea mixed forest and evergreen broad-leaved forest as the study objects, collected the surface soil (0-10 cm) of these three forest types, and measured soil organic carbon(SOC), carbon components, biotic and abiotic factors. The results were as follows: (1) The diversity of understory plants in bamboo-tea mixed forest was significantly lower than that in pure bamboo forest, but there was no significant differences in soil organic carbondensity[ (22.54 ± 2.09) t·hm-2 ] and carbon composition between bamboo-tea mixed forest and pure bamboo forest (P>0.05). Mineral-associated organic carbon (MOC) was (20.13 ± 1.83) g · kg-1, accounting for 92.66% of total organic carbon. The soil organic carbon density of evergreen broad-leaved forest was 41.15% and 41.00% higher than that of bamboo-tea mixed forest and pure bamboo forest (P>0.05). (2) Soil microbial biomass carbon(MBC) content, 16S rRNA gene abundance, cbbL gene abundance and microbial carbon use efficiency of these forest types were (0.58 - 3.08) g·kg-1, (2.18×1010 - 5.65×1010) copies·g-1, (0.37×108-1.10×108) copies·g-1, 0.03 - 0.28, respectively. But there were significant differences about these microbial indicators between three forest types (P>0.05). (3) SOC of the three forest types was significantly negatively correlated with soil pH, gravel content and aboveground litter biomass, and significantly positively correlated with soil clay content, silt content, total nitrogen, C∶N, total phosphorus and ammonium nitrogen (P<0.05). (4) In terms of different carbon fractions, particulate organic carbon (POC) and MOC were significantly negatively correlated with soil pH, gravel content and root biomass, and significantly positively correlated with soil water content, clay content, silt content, total nitrogen, C∶N, total phosphorus and ammonium nitrogen (P<0.05). In conclusion, the mixed transformation of bamboo and tea will cause the decrease of understory vegetation diversity in the original pure bamboo forest, but it will not cause the decrease of soil carbon storage; compared with evergreen broad-leaved forest, it is necessary to improve the management measures of bamboo to improve its carbon sink efficiency.
Key words: particulate organic carbon,mineral-associated organic carbon,soil orgnaic carbon density,Phyllostachys edulis forest,bamboo-tea mixed forest
随着“碳中和”目标的提出,未来40年中国会实施更有力的政策减少碳排放、增加碳汇。但是,合理政策的制定依赖于对增汇减排的科学认知。土壤是陆地最大碳储存库,通过增加土壤碳储量被认为是实现“碳中和”目标的有效途径(杨元合等,2022)。近自然造林以及植被恢复的土壤碳汇效应已经被广泛认知(Don et al., 2011; Lu et al., 2018),如何提升经济林的土壤碳汇效益已成为当前关注的热点(Yang et al., 2021b; Villa et al., 2022)。
毛竹(Phyllostachys edulis)是我国价值较高的竹类品种,广泛分布于南方山地丘陵地区,约占全国竹林面积的74%,其主要经济价值在于生产竹材与竹笋(杨传宝等,2020)。毛竹在经济林种植中占据重要地位,以及快速生长特性,使其碳汇效益受到了广泛关注。毛竹地上部分的年均固碳量较高,为(8.13±2.15)Mg·hm-2(Yen & Lee, 2011)。目前,主要围绕毛竹入侵与经营管理措施两个方面开展毛竹土壤碳库研究。相较于常绿阔叶林,毛竹的种植与入侵往往会造成对土壤碳储量的下降(Wang et al., 2019;祁雪连等,2021),同时引起颗粒有机碳(particulate organic carbon, POC)与矿物结合态有机碳(mineral-associated organic carbon, MOC)含量与占比的变化(Yang et al., 2021b)。不同的管理措施(如间伐、林下植被清除)和混交模式(如竹-杉木混交)会对毛竹林土壤碳库产生不同的影响(漆良华等,2013;李光敏等,2019;Yang et al., 2021a)。Yang等(2021a)在浙江安吉的研究表明,粗放经营的毛竹林比无经营和集约经营的毛竹林具有更高的团聚体稳定性和有机碳储量。生物与非生物因素对毛竹土壤碳库影响因研究地区与经营管理措施不同也存在差异性(张厚喜等,2019;Yang et al., 2021a)。
目前,毛竹单一的经济收入模式难以满足农民收入水平提高的热切需求。同时,近年来受竹材加工业低迷的影响,毛竹种植户的积极性也备受打击。因此,发展林下经济成为了增加农户收入、推动毛竹产业可持续发展的重要举措(蔡春菊等,2018;Zhang et al., 2019)。在关注毛竹林林下种植经济效益的同时,其生态效益也是研究的重点,涉及土壤养分、微生物、水土保持等方面(蔡泽宇等,2017;王勤等,2020;曹小青等,2022)。王勤等(2020)对不同毛竹-多花黄精复合模式的研究显示,低毛竹立竹密度的竹-药复合模式土壤有机碳(soil organic carbon, SOC)含量要比高密度毛竹立竹模式高13%。曹小青等(2022)的研究结果显示,随着套种年限的增加,毛竹-白及复合模式的土壤微生物多样性逐渐提高。但是目前,毛竹林下复合经营的生态效益评估研究有限,并且多是关注毛竹与中草药复合经营带来的变化,缺乏对毛竹与茶树复合经营的碳汇/源作用研究。同时,竹茶种植模式同样是提升毛竹经济效益的重要林下经济举措,特别是具有特色茶资源的竹产区。因此,开展竹茶混交模式的土壤碳库变化研究对增进该改造模式的生态效益认知、促进毛竹林改造的经济与生态效益协同提升建设具有重要意义。
目前,竹茶混交改造对毛竹林土壤碳库储量及其碳组分的影响研究相对缺乏。鉴于此,本文以广西猫儿山地区毛竹纯林、竹茶混交林和天然常绿阔叶林为研究对象,探究竹茶混交模式对毛竹林土壤碳储量的影响,分析POC与MOC含量及其占比变化,揭示影响土壤碳储量与碳组分变化的关键因素。基于土壤碳储量与碳组分评估,本研究成果可为提高毛竹林林下竹茶改造提供理论依据与技术支撑。
1 材料与方法
1.1 研究区概况
研究区位于广西桂林市华江瑶族乡,地处110°27′ E、25°48′ N,海拔在300~500 m之间,属于中亚热带山地气候,年均气温16.40 ℃,年均降水量在2 100 mm以上。成土母质主要为花岗岩风化母质,土壤主要以红壤和黄壤为主。华江瑶族乡是全国十大毛竹生产基地之一,拥有毛竹林面积约(1.53×104)hm2且毛竹林面积有逐年扩张的趋势,该地区野生药用植物和野生茶树资源丰富(宜丽娜等,2012)。在乡村振兴的背景下,当地依托特色茶资源开展竹茶复合经营的毛竹纯林改造,以提升毛竹的经济效益。
1.2 研究方法
1.2.1 样地设置 本研究共设置3种林分样地类型,分别为毛竹纯林(pure bamboo forest, PBF)、竹茶混交林(bamboo-tea mixed forest, BMF)和天然常绿阔叶林(evergreen broad-leaved forest, EBF)。每种林分类型设置6个立地条件接近的重复样方(20 m × 20 m),并进行群落学调查。常绿阔叶林与毛竹林分布在相同的海拔位置。3种林分类型的样地之间至少间隔1 000 m,重复样方之间至少间隔500 m。选取的毛竹纯林样地一直为单一种植毛竹,每年都会进行周期性的间伐与林下植被清耕。选取的竹茶混交林样地的乔木层为单一毛竹,每年会进行毛竹间伐;其林下茶树为自然生长的野生茶树,不存在人为管理,但每年春季会進行采茶,茶树因有一定经济价值得以保留。本研究以该自然存在的竹茶混交林作为毛竹纯林改造的参照对象。3种林分样地基本信息见表1。
1.2.2 样地调查与样品采集 在2021年11月对样地进行调查与采样。调查内容包括海拔、坡度、坡向、植物组成、经营措施等。其中,乔灌植物调查采用每木检尺,草本植物调查采用小样方法。土壤样品采集则采用土钻在每个样方内随机采集5个0~10 cm子土样,再混合成一个土样,作为该样地的土样。同时,在每个采样点附近收集土壤测定土壤容重,用铝盒收集土壤测定含水率。在样方内随机设置5个50 cm × 50 cm的小样方收集地上凋落物,设置5个20 cm × 20 cm的小样方收集0~10 cm层土壤根系。收集的土样带回实验室后,一部分自然风干,一部分保存在4 ℃冰箱用于鲜土实验测定,一部分保存在-80 ℃下以便进行微生物DNA提取。
1.2.3 土壤理化性质与微生物碳代谢相关指标测定 土壤全碳与总氮含量采用碳氮元素分析仪(Elementar Vario EL Ⅲ,德国)测定。土壤容重采用环刀法测定。pH采用电位法测定(水土质量比为2.5∶1)。总磷(total phosphorus, TP)含量采用钼锑钪比色法测定。土壤速效磷(available phosphorus, AP)采用 1/2硫酸消煮法然后用钼锑抗比色法测定。土壤铵态氮(NH4+-N)与硝态氮(NO3--N)含量,采用2 mol·L-1 KCl溶液浸提及SKALAR SAN++连续流动分析仪测定。土壤机械组成测定采用比重计法,即50 g风干土样混合分散剂煮沸后,过滤至1 000 mL沉降筒内,采用比重计进行测定。采用国际制将土壤机械组成划分为砂粒、粉粒和黏粒。土壤有机碳含量采用浓硫酸-重铬酸钾高温外加热氧化法测定。地上凋落物与根系生物量采用烘干法测定。采用六偏磷酸钠分散法区分土壤颗粒态有机碳和矿物结合态有机碳。具体方法如下:自然风干的土壤样品加入0.5%的六偏磷酸钠溶液(土液质量比1∶3)和玻璃珠,振荡后充分分散;利用53 μm筛过滤分组为矿物结合态有机质(<53 μm)和颗粒态有机质(>53 μm);分离后的组分用去离子水洗后60 °C烘干,待测;结合土层厚度、容重等土壤理化性质来计算土壤有机碳密度(soil organic carbon density, SOCD),公式如下。
SOCD=SOC×ρ×D×10 (1)
式中: SOC为土壤有机碳的平均含量(g·kg-1); D为土层厚度(cm); ρ为土壤容重(g·cm-3)。
土壤微生物量碳(microbial biomass carbon, MBC)含量采用氯仿熏蒸法测定(汪亚芳等,2022)。16S rRNA与cbbL基因采用qPCR测定,引物序列分别为F515/R907和K2f/V2f(Qin et al., 2020)。采用 H218O 标记土壤样品中的微生物,测定微生物碳利用效率(carbon use efficiency, CUE),参考Zheng等(2019)方法。
1.2.4 数据处理与分析 采用 Excel 2016、SPSS 25和Origin 2021进行数据整理与分析。采用单因素方差分析(ANOVA)分析不同处理组间变量显著性差异。采用 Pearson 相关分析揭示变量间相互关系,通过多元线性回归模型逐步法揭示土壤理化因素与微生物指标对土壤有机碳的影响。所有统计的显著性水平设置为P<0.05。
2 结果与分析
2.1 不同林分类型土壤理化性质差异
3种林分类型中土壤pH,黏粒和砂粒含量,总磷及硝态氮含量均存在显著差异(P<0.05)。其中,3种林分类型的土壤均为酸性土壤且竹茶混交林的土壤pH值最高。毛竹纯林的土壤含水率最低且土壤的砂粒含量显著高于其他2个林分类型(P<0.05),其土壤养分含量(包括总磷和硝态氮)均低于竹茶混交林和常绿阔叶林。竹茶混交林及毛竹纯林的根系生物量、C∶N、总氮和铵态氮含量无显著差异,但与常绿阔叶林存在显著差异(P<0.05)。其中,二者的根系生物量显著高于常绿阔叶林,而其他指標则显著低于常绿阔叶林(P<0.05)(表2)。
2.2 不同林分类型SOC与SOCD变化特征
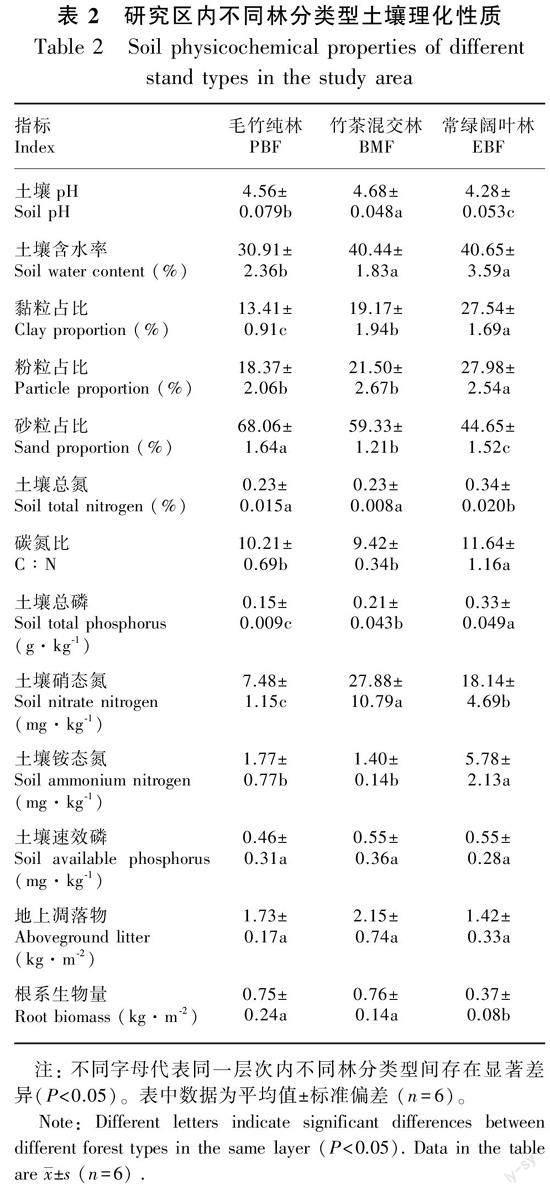
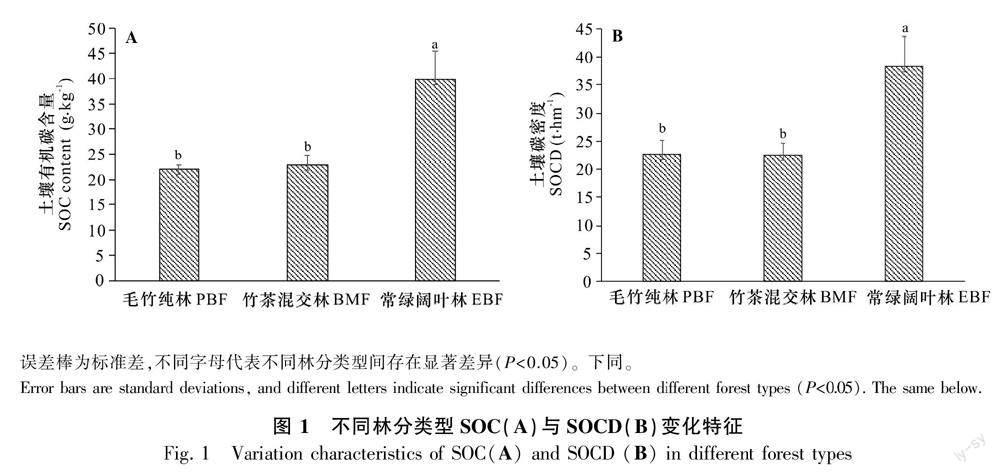
竹茶混交林表层SOC为(22.95±1.91)g·kg-1,与毛竹纯林SOC不存在显著差异(P>0.05),比常绿阔叶林(39.74±5.82)g·kg-1低42.25%(P<0.05)。3种林分类型SOCD变化范围为(19.65~44.09)t·hm-2。其中,竹茶混交林与毛竹纯林表层SOCD分别为(22.54±2.09)、(22.60±2.53)t·hm-2,均显著低于常绿阔叶林(38.31±5.40)t·hm-2(P<0.05)(图1)。
2.3 不同林分类型有机碳组分含量变化特征
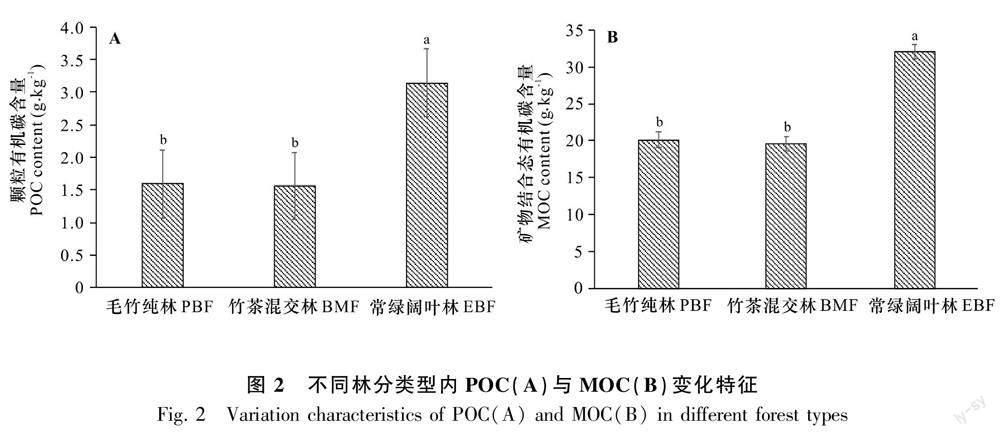
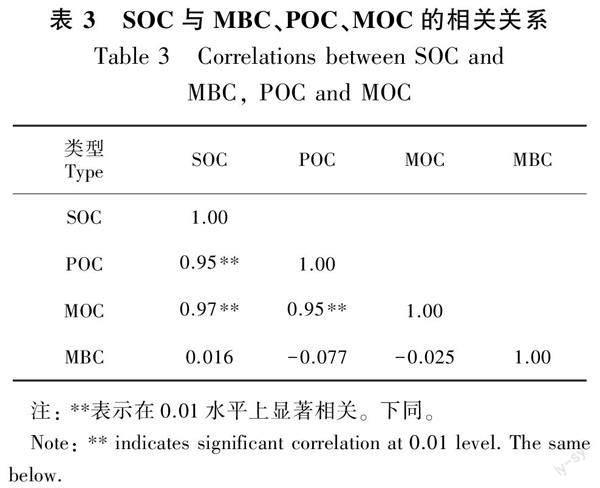
3种林分类型土壤均以矿物结合态有机碳为主,MOC的范围为17.98~36.83 g·kg-1,对SOC的贡献占90.19%~94.40%,剩余贡献部分为POC;不同林分类型之间POC和MOC对SOC的贡献不存在显著差异(P>0.05)。由表3相关性分析显示,SOC与POC、MOC呈极显著正相关,相关系数分别为0.95、0.97(P<0.05)。常绿阔叶林土壤POC和MOC分别为(3.14±0.42)g·kg-1和(32.09±3.54)g·kg-1,均分别显著高于其毛竹纯林和竹茶混交林(P<0.05)。竹茶混交林POC和MOC分别为(1.59±0.32)g·kg-1和(20.13±1.83)g·kg-1,与毛竹纯林不存在显著差异(P>0.05)(图2)。
2.4 不同林分类型微生物碳代谢相关指标变化特征
3种林分类型土壤微生物量碳含量范围为0.58~3.08 g·kg-1,其与SOC之间相关性并不明显(P>0.05)(表3)。不同林分类型之间微生物量碳含量不存在显著差异(P>0.05),其中竹茶混交林土壤微生物量碳含量为(1.47±0.34)g·kg-1。竹茶混交林土壤16S rRNA丰度平均为3.91×1010copies·g-1;固碳基因cbbL丰度为0.76×108copies·g-1,占总细菌丰度的0.21%。竹茶混交林与毛竹纯林和常绿阔叶林在16S rRNA丰度和cbbL丰度上不存在显著差异(P>0.05)。3种林分类型土壤微生物碳利用效率范围为0.03~0.28,3种林分类型之间微生物碳利用效率(carbon use efficiency, CUE)不存在显著差异(P>0.05)(表4)。
2.5 各有机碳组分与环境因子相关性
由表5可知,3种林分类型SOC与土壤pH、砂粒含量和地上凋落物生物量呈显著负相关, 与土壤黏粒含量、粉粒含量、总氮、C∶N、总磷和铵态氮含量呈显著正相关 (P<0.05)。就不同碳组分而言,POC和MOC均与土壤pH、砂粒含量和根系生物量呈显著负相关,与土壤含水率、黏粒含量、粉粒含量、总氮、C∶N、总磷和铵态氮含量呈显著正相关(P<0.05)(表5)。多元线性回归结果显示,土壤总氮含量与C∶N是预测土壤碳密度的关键因素,可解释土壤碳密度变化的97%,公式如下。
y=-22.12+11.66CN+1.81RC∶N (R2=0.97, P<0.01) (2)
式中:y为土壤碳密度(t·hm-2); CN为总氮含量(g·kg-1); RC∶N为碳氮质量比。
3 讨论
3.1 毛竹林改造对土壤碳储量和组分的影响
在研究区内开展毛竹纯林的竹茶混交模式改造,从长期来看并不会引起土壤有机碳储量的下降。研究结果显示,毛竹纯林与竹茶混交林SOCD不存在显著差异。以往对乔木树种与林下植被多样性研究表明,物种多样性与土壤碳储量多呈正相关(Zhao et al., 2014;Chen et al., 2020)。这种差异性可能是由于毛竹纯林与竹茶混交林的碳循环均主要受毛竹的支配。这两种林分类型乔木层只有毛竹,而毛竹根系发达,占据着表层土壤,影响着土壤有机碳与其他养分循环。刘骏等(2013)研究表明,毛竹林细根生物量是常绿阔叶林的5.86倍。虽然竹茶模式改造使毛竹的株密度下降了约20%,但两者的表层土壤根系生物量并不存在显著性差异。竹茶混交改造遗留的地下空间会迅速被其他毛竹根系占据。此外在毛竹砍伐期间,种植者会清除林下植被,这种周期性更新也削弱了毛竹纯林内植物源SOC的输入与积累。李光敏等(2019)的研究结果显示,毛竹林林下植被清除2年后其SOC与对照组之间不存在显著差异。在毛竹的支配性地位与当前管理方式的影响下,毛竹纯林的竹茶混交模式改造对表层土壤碳储量的影响有限。
虽然毛竹林林下改造不会造成土壤碳储量的下降,但这种种植模式并不利于SOC的固持。本研究显示,毛竹纯林与竹茶混交林的SOCD分别比常绿阔叶林低41.15%和41.00% (P<0.05)。这说明将常绿阔叶林转化为毛竹纯林和竹茶混交林均会造成土壤碳储量的流失,这与其他地区的研究结果一致(Lin et al., 2018;Wang et al., 2019;祁雪连等,2021)。Lin等(2018)通过对常绿阔叶林转为毛竹林的对比研究表明,这种土地变化会使0~40 cm层土壤碳储量下降12%。毛竹经营管理、乔木层树种差异及其影响下的土壤理化性质差异是造成毛竹纯林、竹茶混交林与常绿阔叶林土壤碳储量存在差异的主要原因。因此,在竹茶混交改造过程中探索如何提高土壤有机碳储量十分必要。
本研究表明,竹茶混交改造不会引起毛竹林土壤有机碳组分含量与占比的显著性变化。3种林分类型土壤均以MOC为主,占总有机碳的比例高达90%以上。这说明这些林地土壤有机碳库以稳定性碳为主,但也反映出POC较难保留。每年植物会通过地上凋落物、根系分泌物等形式向土壤释放碳源,再经分解转化为POC;而3种林分类型POC占比极小,大部分新输入的碳不可能马上转化为稳定性的MOC,极可能是被损失掉。POC的损失可能由该地区充沛的降雨造成。较强的降水既能将植物凋落物冲走,减少其原地转化为POC的可能,同时也能将不稳定的POC随径流带走,减少其保留。董玉清等(2020)在猫儿山相近海拔的常绿阔叶林研究中,也发现POC占总有机碳的比例在1.26%~14.44%之间。因此,有必要在这个地区开展碳示踪研究,以揭示碳的转化归趋。
3.2 影响土壤有机碳及其组分变化的主要因素
土壤有机碳的转化和积累与生物、非生物因素存在密切联系。本研究中土壤有机碳及其组分与总氮、C∶N、总磷和铵态氮含量呈极显著正相关(P<0.01),并且回归分析显示总氮是影响有机碳及其组分的最关键因素。微生物生长需要氮源,当氮源缺乏时其会选择降解凋落物与有机质以获取氮源。这个过程会造成有机碳含量的下降(Frey et al., 2014;Lu et al., 2021)。因此,充足的氮源有助于SOC的积累。在一些研究中,同样发现土壤总氮和无机氮含量与有机碳含量呈正相关的现象(李萍,2010;Yang et al., 2021b)。李萍(2010)对江西大岗山毛竹林的研究同樣发现土壤总氮是影响土壤有机碳变化的最大因素且彼此之间呈正相关。因此,在毛竹的管理中可以适当添加氮肥,改变毛竹林土壤有机碳较低的现象,以提高土壤碳汇能力。
整体而言,3种林分类型SOC和各碳组分含量与土壤pH呈极显著负相关(P<0.01)。这说明pH越低越有利于土壤有机碳的积累。土壤 pH可以通过影响微生物群落结构与活性而影响有机碳的分解与转化。pH过高与过低均不利于微生物的生长与活动,从而抑制有机碳的分解转化(Zhang et al., 2019)。张厚喜等(2019)对武夷山不同海拔梯度毛竹林土壤有机碳特征及影响因素中也发现相同现象。本研究中SOC及其组分与土壤黏粒含量、粉粒含量呈极显著正相关(P<0.01)。这主要是因为粉粒和黏粒对SOC存在保护作用,减少碳的矿化分解与流失(Lehmann & Kleber,2015;Kasmerchak et al.,2018)。但是,竹茶混交林比毛竹纯林具有更高的黏粒和粉粒含量,而两者之间的SOCD并无显著差异。这说明黏粒和粉粒含量可能需要达到一定阈值才会影响到有机碳的积累,也可能是其他关键因素抵消了竹茶混交林黏粒和粉粒对有机碳积累的贡献。因此,毛竹林林下种植茶树至少可以改善土壤机械组成,朝有利于有机碳积累的方向发展。
土壤微生物是影响土壤碳循环的关键因素,参与SOC的积累与分解。但是,基于对土壤微生物量碳含量、固碳基因丰度、碳利用效率指标的分析显示,本研究区内微生物活动不是影响土壤有机碳储量变化的决定因素。汪亚芳等(2022)研究同样发现毛竹林与常绿阔叶林土壤微生物量之间不存在显著差异。此外,Bai等(2016)的研究还发现毛竹入侵阔叶林会造成SOC储量下降,但微生物量碳得到了提高。因此,还需要开展动态和多季节性研究来揭示微生物对土壤碳库的影响。
4 结论
本研究结果表明,相较于毛竹纯林,毛竹林林下种植茶树会显著提高土壤pH、含水率、黏粒、粉粒含量、总磷与硝态氮含量。毛竹林下茶树的种植会造成林下灌木层植物多样性下降,但不会造成土壤有机碳密度的下降,以及有机碳组分比例的改变。毛竹纯林与竹茶混交林的土壤有机碳储量显著低于常绿阔叶林。3种林分的碳组分均以矿物结合态有机碳为主。因此,需要改进现有的毛竹纯林与竹茶混交林经营管理模式,在提升经济效益的同时,注重碳汇效益的协同发展。此外,本研究从土壤碳汇的角度反映生态效益,既有一定的代表性,也有一定的局限性。未来还需要开展更多元的生态效益评估,以揭示竹茶改造模式对毛竹纯林生态系统功能的影响,从而为竹茶改造模式提供科学依据与指导。
参考文献:
BAI SB, CONANT RT, ZHOU GM, et al., 2016. Effects of Moso bamboo encroachment into native, broad-leaved forests on soil carbon and nitrogen pools [J]. Sci Rep, 6(1): 31480.
CAI CJ, FAN SH, LIU GL, et al., 2018. Research and development advance of compound management of bamboo forests [J]. World Bamboo Ratt, 16(5): 47-52. [蔡春菊, 范少辉, 刘广路, 等, 2018. 竹林复合经营研究和发展现状[J]. 世界竹藤通讯, 16(5): 47-52.]
CAI ZY, ZHANG JF, SUN SY, et al., 2017. Influence of Phyllostachy edulis stand improvement on runoff and nitrogen and phosphorus losses in Anji, Zhejiang, China[J]. J Soil Water Conserv, 31(6): 46-51. [蔡泽宇, 张建锋, 孙士咏, 等, 2017. 安吉毛竹林分改造对地表径流及氮磷流失的影响[J]. 水土保持学报, 31(6): 46-51.]
CAO XQ, WANG L, SUN MY, et al., 2022. Soil microbial community diversity in Phyllostachys pubescens-Bletilla striata ecosystems with different intercropping years[J]. SoilFert Sci Chin, (1): 147-154. [曹小青, 王亮, 孙孟瑶, 等, 2022. 不同年限毛竹-白及复合系统土壤微生物群落多样性特点[J]. 中国土壤与肥料, (1): 147-154.]
CHEN XL, CHEN HYH, CHEN C, et al., 2020. Effects of plant diversity on soil carbon in diverse ecosystems: a global meta-analysis [J]. Biol Rev, 95: 167-183.
DON A, SCHUMACHER J, FREIBAUER A, 2011. Impact of tropical land-use change on soil organic carbon stocks — a meta-analysis [J]. Glob Change Biol, 17(4): 1658-1670.
DONG YQ, GUAN P, LU Y, et al.,2020. Fractionsand contents of soil organic carbon at different elevations in Maoer Mountain [J]. Chin J Soil Sci, 51(5): 1142-1151. [董玉清, 官鵬, 卢瑛, 等, 2020. 猫儿山不同海拔土壤有机碳组分构成及含量特征 [J]. 土壤通报, 51(5): 1142-1151.]
FREY SD, OLLINGER S, NADELHOFFER K, et al., 2014. Chronic nitrogen additions suppress decomposition and sequester soil carbon in temperate forests [J]. Biogeochemistry, 121(2): 305-316.
KASMERCHAK CS, MASON JA, LIANG MY, 2018. Laser diffraction analysis of aggregate stability and disintegration in forest and grassland soils of northern Minnesota, USA [J]. Geoderma, 338: 430-444.
LEHMANN J, KLEBER M, 2015. The contentious nature of soil organic matter [J]. Nature, 528: 60-68.
LI GM, CHEN FS, XU ZW, et al., 2019. Effects of thinning and understory removal on soil labile organic carbon in moso plantation [J]. Acta Agric Univ Jiangxi, 41(4): 733-740. [李光敏, 陈伏生, 徐志文, 等, 2019. 间伐和林下植被剔除对毛竹林土壤活性有机碳的影响 [J]. 江西农业大学学报, 41(4): 733-740.]
LI P, 2010. Soil organic carbon and its relationship with soil properties in Phyllostachys forests [D]. Beijing: Beijing Forestry University. [李萍, 2010. 毛竹林土壤有机碳变化及其与土壤性质的关系[D]. 北京: 北京林业大学.]
LIN ZW, LI YF, TANG CX, et al., 2018. Converting natural evergreen broadleaf forests to intensively managed Moso bamboo plantations affects the pool size and stability of soil organic carbon and enzyme activities [J] Biol Fert Soils, 54(4): 467-480.
LIU J, YANG QP, YU DK, et al., 2013. Contribution of fine root to soil nutrient heterogeneity at two sides of the bamboo and broadleaved forest interface [J]. Chin J Plant Ecol, 37(8): 739-749. [刘骏, 杨清培, 余定坤, 等, 2013. 细根对竹林-阔叶林界面两侧土壤养分异质性形成的贡献 [J]. 植物生態学报, 37(8): 739-749.]
LU F, HU HF, SUN WJ, et al., 2018. Effect of national ecological restoration projects on carbon sequestration in China from 2001 to 2010 [J]. PNAS, 115: 4039-4044.
LU XK, VITOUSEK PM, MAO QG, et al., 2021. Nitrogen deposition accelerates soil carbon sequestration in tropical forests [J]. PNAS, 118(16): e2020790118.
QI LH, FAN SH, DU MY, et al., 2013. Vertical distribution and seasonal dynamicsof soil organic carbon in Phyllostachy edulis plantations and P. edulis-Cunninghamia lanceolata mixed forests in the hilly region of Central Hunan, Southern China[J]. Sci Silv Sin, 49(3): 17-24. [漆良华, 范少辉, 杜满义, 等, 2013. 湘中丘陵区毛竹纯林、毛竹-杉木混交林土壤有机碳垂直分布与季节动态[J]. 林业科学, 49(3): 17-24.]
QI XL, GE XM, QIAN ZZ, et al., 2021. Differences of soil properties between natural mixed coniferous and broad-leaved forest and Moso bamboo plantation in Wuyi Mountains [J]. Ecol Environ Sci, 30(8): 1599-1606. [祁雪连, 葛晓敏, 钱壮壮, 等, 2021. 武夷山天然针阔混交林与毛竹人工林土壤性质差异 [J]. 生态环境学报, 30(8): 1599-1606.]
QIN YB, CHEN ZH, DING BJ, et al., 2020. Impact of sand mining on the carbon sequestration and nitrogen removal ability of soil in the riparian area of Lijiang River, China [J]. Environ Poll, 261: 114220.
VILLA MVE, CRISSTIANO PM, DE D, et al., 2022. Do selective logging and pine plantations in humid subtropical forests affect aboveground primary productivity as well as carbon and nutrients transfer to soil? [J]. For Ecol Manage, 503(1) : 119736.
WANG Q, SUN MY, DI JH, et al., 2020. Effects of different Phyllostachys pubescens-Polygonatum sibiricum intercropping models on soil physicochemical properties [J]. Ecol Sci, 39(6): 54-59. [王勤, 孙梦瑶, 遆建航, 等, 2020. 毛竹-多花黄精复合经营模式对土壤理化特性的影响[J]. 生态科学, 39(6): 54-59.]
WANG YF, LIU ZY, ZHANG BG, et al., 2022.Effects of invasive Moso bamboo removal on soil microbial biomass and enzyme activities in subtropical forests [J]. Chin J Appl Ecol, 33(5): 1233-1239. [汪亚芳, 刘宗悦, 张宝刚, 等, 2022. 入侵毛竹皆伐对亚热带森林土壤微生物生物量和酶活性的影响 [J]. 应用生态学报, 33(5): 1233-1239.]
WANG HB, JIN J, YU PY, et al., 2019. Converting evergreen broad-leaved forests into tea and Moso bamboo plantations affects labile carbon pools and the chemical composition of soil organic carbon [J]. Sci Total Environ, 711(1): 135225.
YANG CB, NI HJ, SU WH, et al., 2020. Effects of management measures on organic carbon, nitrogen and chemical structure of different soil fractions in Phyllostachys edulis plantations [J]. Chin J Appl Ecol, 31(1) : 25-34. [杨传宝, 倪惠菁, 苏文会, 等, 2020. 经营措施对毛竹林土壤不同组分有机碳、氮及化学结构的影响 [J]. 应用生态学报, 31(1): 25-34.]
YANG CB, WANG AK, ZHU ZX, et al., 2021a. Impact of extensive management system on soil properties and carbon sequestration under an age chronosequence of Moso bamboo plantations in subtropical China [J]. For Ecol Manage, 497(1): 119535.
YANG CB, ZHANG XP, NI HJ, et al., 2021b. Soil carbon and associated bacterial community shifts driven by fine root traits along a chronosequence of Moso bamboo (Phyllostachys edulis) plantations in subtropical China [J]. Sci Total Environ, 752(15): 142333.
YANG YH, SHI Y, SUN WJ, et al., 2022. Terrestrial carbon sinks in China and around the world and their contribution to carbon neutrality [J]. Sci Sin (Vit), 52(4): 534-574. [杨元合, 石岳, 孙文娟, 等, 2022. 中国及全球陆地生态系统碳源汇特征及其对碳中和的贡献 [J]. 中国科学(生命科学), 52(4): 534-574.]
YEN TM, LEE JS, 2011. Comparing aboveground carbon sequestration between Moso bamboo (Phyllostachys heterocycla) and China fir (Cunninghamia lanceolata) forests based on the allometric model [J]. For Ecol Manage, 261(6) : 995-1002.
YI LN, BAI KD, LI MX, et al., 2012. Comparison of physiology and biochemical index of some evergreen and deciduous broad-leaved species in Maoer Mountain, Guangxi, China [J]. J Guangxi Norm Univ (Nat Sci Ed), 30(1): 83-88. [宜麗娜, 白坤栋, 李明霞, 等, 2012. 猫儿山常绿和落叶阔叶树抗性生理生化指标的比较 [J]. 广西师范大学学报 (自然科学版), 30(1): 83-88.]
ZHAO FZ, KANG D, HAN XH, et al., 2014. Soil stoichiometry and carbon storage in long-term afforestation soil affected by understory vegetation diversity [J]. Ecol Eng, 74: 415-422.
ZHANG HX, LIN C, CHENG H, et al., 2019. Variation of soil organic carbon content of Moso bamboo forest along altitudinal gradient in Wuyi Mountain in China [J]. Soils, 51(4): 821-828. [张厚喜, 林丛, 程浩, 等, 2019. 武夷山不同海拔梯度毛竹林土壤有机碳特征及影响因素 [J]. 土壤, 51(4): 821-828.]
ZHANG XP, GUO GB, WU ZZ, et al., 2019. Agroforestry alters the rhizosphere soil bacterial and fungal communities of Moso bamboo plantations in subtropical China [J]. Appl Soil Ecol, 143: 192-200.
ZHENG Q, HU YT, ZHENG SS, et al., 2019. Growth explains microbial carbon use efficiency across soils differing in land use and geology [J]. Soil Biol Biochem, 128: 45-55.
(责任编辑 李 莉 王登惠)
猜你喜欢
天津农林科技(2022年2期)2022-04-19 10:48:14
浙江农林大学学报(2020年5期)2020-11-05 09:12:52
湖州职业技术学院学报(2020年4期)2020-03-30 13:13:02
乡村科技(2018年8期)2018-06-27 03:41:40
浙江农林大学学报(2018年1期)2018-01-16 09:52:52
现代农业科技(2017年15期)2017-09-15 18:59:29
绿色科技(2017年15期)2017-09-01 19:57:53
农家科技下旬刊(2017年2期)2017-03-27 12:01:29
湖南林业科技(2017年1期)2017-02-06 05:29:18
防护林科技(2013年9期)2013-08-08 07:29:40
