
不同光质与光周期对粗肋草生长、叶片色素和颜色的影响
2023-11-01 04:51:20李文杨张光辉李月凤梁祥鹏尹娟
广西植物 2023年9期
李文杨 张光辉 李月凤 梁祥鹏 尹娟
摘 要:為改良粗肋草叶色和优化粗肋草设施栽培体系,该研究以粗肋草品种‘吉利红’的水培苗为材料,设置6种光质(白光、R∶B=1∶1、R∶B=1∶2、R∶B=2∶1、R∶B=1∶3、R∶B=3∶1,其中R、B分别代表红光、蓝光)和2种光周期(8、12 h·d)交叉培养,测定粗肋草的生长量、生物量、土壤和作物分析仪器开发(SPAD)值、花色素苷含量和叶片颜色参数(色相值a*、色相值b*、明度值L*、色调角h*),研究粗肋草对红蓝光质和光周期互作的响应。结果表明:(1)12 h·d光周期更有利于粗肋草生物量的积累,其中LP11(R∶B=1∶3×12 h·d)处理的粗肋草苗木干重、鲜重均为最高,最有利于植物生长及生物量积累;其次是LP5(R∶B=1∶3×8 h·d)处理。(2)相同光质条件下,8 h·d光周期处理的粗肋草叶片SPAD值比12 h·d光周期处理高,12 h·d光周期处理的粗肋草叶片花色素苷含量高于8 h·d光周期的处理,LP11处理叶片SPAD值最低,花色素苷含量最高。12 h·d光周期培养的粗肋草叶片颜色参数a*、b*值比8 h·d光周期高,h*值比8 h·d光周期低。(3)主成分分析结果表明,LP11处理在促进粗肋草生长及叶色改良方面效果最好。综上认为,12 h·d光周期更有利于粗肋草的生长和叶片颜色变化,其中LP11处理为最佳光质和光周期组合。
关键词: 光质, 光周期, 粗肋草, 叶色参数, 色素含量
中图分类号:Q945
文献标识码:A
文章编号:1000-3142(2023)09-1725-12
收稿日期:2023-07-15
基金项目:河南省高等学校重点科研项目(19A22005); 信阳农林学院大别山林业资源创新理论与技术科技创新团队项目(XNKJTD-004)。
第一作者: 李文杨(1984-),硕士,副教授,主要从事经济林丰产栽培及逆境生理研究,(E-mail)wylee126@126.com。
*通信作者:尹娟,硕士,副教授,主要从事园林植物栽培与应用研究,(E-mail)wyxtkt@126.com。
Effects of different light qualities and photoperiods
on the growth, leaf pigment and color
of Aglaonema commutatum
LI Wenyang1, ZHANG Guanghui1, LI Yuefeng2, LIANG Xiangpeng1, YIN Juan1*
( 1. College of Forestry, Xinyang Agriculture and Forestry University/Key Laboratory of Tree Genetics and Breeding in Xinyang,
Xinyang 464000, Henan, China; 2. Forest Science Research Institute of Xinyang, Xinyang 464031, Henan, China )
Abstract:In order to improve the leaf color and optimize the facility of cultivation system of Aglaonema commutatum ‘Big apple’, its hydroponic seedlings were used as the test material in this study, six light qualities (white light, R∶B = 1∶1, R∶B = 1∶2, R∶B = 2∶1, R∶B = 1∶3, R∶B = 3∶1, in which R and B represent red light and blue light, respectively) and two photoperiods (8 h·d-1, 12 h·d-1) were cross cultured to determine the growth amount, biomass, soil and plant analyzer development (SPAD) value, anthocyanin content and leaf color parameters (hue value a*, hue value b*, luminosity L* and hue angle h*), and to investigate its response to red and blue light qualities and photoperiodic interactions. The results were as follows: (1) The dry weight and fresh weight of A. commutatum seedlings treated with LP11 (R∶B=1∶3×12 h·d-1) were the highest, which was most conducive to plant growth and biomass accumulation; followed by LP5 (R∶B=1∶3×8 h·d-1) treatment. (2) Under the same light quality condition, the SPAD value of 8 h·d-1 photoperiod was higher than that of 12 h·d-1 photoperiod, the anthocyanin content of 12 h·d-1 photoperiod was higher than that of 8 h·d-1 photoperiod, the SPAD value of LP11 treatment was the lowest and the anthocyanin content was the highest. The leaf color parameters a* and b* of A. commutatum cultured in 12 h·d-1 photoperiod were higher than those in 8 h·d-1 photoperiod, and the value of h* was lower than that in 8 h·d-1 photoperiod. (3) The principal component analysis showed that the LP11 treatment was the most effective in promoting the growth and leaf color improvement of A. commutatum. In conclusion, 12 h·d-1 photoperiod is more conducive to the growth and leaf color change of A. commutatum, and LP11 treatment is the best combination of light quality and photoperiod.
Key words: light quality, photoperiod, Aglaonema commutatum, leaf color parameters, pigment content
光是影响植物生长发育最重要的环境信号之一,主要通过光质、光周期及光强对植物形态建成以及植物生长发育的各个阶段起到显著作用(王冬雪等,2019)。光质会影响植物对营养物质的吸收以及气孔导度(Kim et al.,2004),不同波长的光对植物的影响不同(张水木等,2016;Shafiq et al., 2021),与其他光质相比,红蓝光被认为是对植物光合作用的最佳光谱(陈钢,2020)。植物开花对昼夜周期的适应反应称为光周期现象。虽然植物处在昼夜交替的环境中,但在光周期诱导中光期和暗期的作用是不相等的,是影响植物生长发育的关键因素。Hayama和Coupland(2004)研究表明,一般长日照能促进植物生长,短日照可抑制枝的伸长生长,促进芽的形成,影响植物开花。光质与光周期互作会造成斑叶植物的植株形态改变及叶斑形成的差异,使植物的叶色出现变化(Mohamed, 2016;Wang et al., 2016;刘敏竹等,2021)。有研究发现,不同光质及光周期可以提高叶山白兰(Paramichelia baillonii)叶片面积(韦秋梅,2018);光质×补光时间互作能有效提高东方百合(Lilium oriental)植株叶片光合色素含量和抗氧化酶活性(胡绍泉,2018);不同光周期处理对草莓(Fragaria × ananassa)光合特性影响不同,草莓叶片的气孔导度和叶片色素含量随着光周期的增加而增加(刘庆,2015);不同光质与光照时间配比,对冰菜(Mesembryanthemum crystallinum)生长形态、光合色素、光合参数等有重要作用(赵明伟,2020)。由此可见,调控适宜植物生长的光环境是苗木培育和规模化生产的关键环节。
粗肋草(Aglaonema commutatum)是天南星科(Araceae)粗肋草属(Aglaonema)多年生草本植物,原产于亚洲热带地区,品种多样,株型和叶色丰富,并具有净化空气、少病害、观赏期长等优点,是一种优良的可在室内外种植的观叶植物,深受人们喜爱,市场需求量大(李林山等,2022)。目前,粗肋草的生产以无性繁殖设施栽培为主,其中组织培养是有效的繁殖手段之一,可以快速获得大量试管苗。粗肋草叶片色彩及条纹美丽,但光照过弱、过强都不利于它的生长,会表现出长势不良、枝叶徒长或衰弱、叶色变淡、焦枯等现象,降低其观赏价值。光作为重要的环境因子对植物叶色转变过程和鲜艳程度具有非常重要的影响(郭力宇等,2018)。不同品种粗肋草的特征特性多体现为叶色的差异(王树茵等,2017),所需适宜受光条件不同,有针对性地筛选光环境来调控粗肋草植株的生长发育和形态建成、缩短培养周期、提高组培苗质量,减少能耗,降低成本,是为了满足市场对优质粗肋草的迫切需求。目前关于粗肋草的研究主要集中在新品种培育(李冬梅等,2022)、离体培养及快繁体系的建立和优化(周佐葡等,2018)等方面,光質与光周期互作对粗肋草品种‘吉利红’(Aglaonema commutatum ‘Big apple’)生长发育及叶片颜色的变化研究较少。本研究以‘吉利红’水培苗为对象,采用发光二极管(light emitting diode,LED)光源设置6种不同光质组合和2种光周期的方法,测定粗肋草的形态指标、生物量、土壤和作物分析仪器开发(soil and plant analyzer development,SPAD)值、花色素苷含量和叶片颜色参数。拟探讨以下问题:(1)不同光质和光照时间对粗肋草生长和叶片颜色的影响以及叶色变化规律;(2)改良粗肋草生长和叶色的最佳光质和光周期组合,优化粗肋草设施栽培体系。本研究以期为叶色对光质与光周期的响应机理提供理论依据。
1 材料与方法
1.1 试验材料
以生长发育良好且长势一致的粗肋草品种‘吉利红’(平均地径为3.98 mm,平均苗高为54.35 mm)为研究对象,使用定植篮(规格为内径43.5 mm、外径60 mm、高50 mm)培养在装有水培液(史丹利植物营养液,水培液∶水=1∶500,V/V)的培养瓶中,进行LED光源[高度为(50±5) cm]光照培养处理,为防止不同光源相互干扰,不同处理之间均设挡光材料。试验在河南省信阳农林学院、信阳市林木遗传育种重点实验室培养间进行,温度(25±2)℃,湿度(50±5)%。
1.2 试验设计
试验采用双因素交叉式分组设计,设置6个光质和2个光周期配比。光质分别为红光(red)、蓝光(blue),记为R、B,光质配比为白光(white,记为W)、R∶B=1∶1、R∶B=1∶2、R∶B=2∶1、R∶B=1∶3、R∶B=3∶1;光周期为8、12 h·d;两两组合共12个处理(LP1:W×8 h·d,LP2:R∶B=1∶1×8 h·d,LP3:R∶B=1∶2×8 h·d,LP4:R∶B=2∶1×8 h·d,LP5:R∶B=1∶3×8 h·d,LP6:R∶B=3∶1×8 h·d,LP7:W×12 h·d,LP8:R∶B=1∶1×12 h·d,LP9:R∶B=1∶2×12 h·d,LP10:R∶B=2∶1×12 h·d,LP11:R∶B=1∶3×12 h·d,LP12:R∶B=3∶1×12 h·d),每个处理10株粗肋草,重复3次。光周期调控采用定时器调控,8 h·d光周期光照时间为9:00—17:00,12 h·d光周期光照时间为7:00—19:00。
1.3 指标测定与处理
1.3.1 生长量、生物量指标 叶长、叶宽采用0.01 mm电子数显游标卡尺测量,测量粗肋草植株最大叶的叶长、叶宽。生物量的测量:每个处理随机选取3株粗肋草,将其从水培液中取出,冲洗干净,并用滤纸吸干粗肋草表面水分后,用0.01 g电子天平分别称量全株、根、茎、叶的鲜重,烘干至恒重后再次称量。计算公式如下:
叶重比=叶干重/全株干重;根重比=根干重/全株干重;茎重比=茎干重/全株干重;根冠比=根干重/(茎干重+叶干重)。
1.3.2 色素含量
1.3.2.1 叶绿素相对含量的测定 选取植株叶面干净、生长正常的叶片,用LYS-B便携式叶绿素测定仪测定粗肋草叶片叶绿素相对含量,用SPAD值表示。
1.3.2.2 花色素苷的提取与测定 参考李力(2016)的方法,称取粗肋草样品0.5 g,研磨至粉末状后置于10 mL具塞试管中,加入5 mL的1%盐酸乙醇,摇匀,确保全部粉末都被完全浸泡,盖上试管塞,于32 ℃恒温水浴锅中避光提取4 h。将提取液倒入离心管中,5 000 r·s-1离心10 min,上清液即为花色素苷提取液。以1%盐酸乙醇为空白对照,使用分光光度计在520 nm波长下测定粗肋草提取液的吸光度(OD)值。以叶片在10 mL提取液中改变0.1个OD值作为一个色素单位U,则花色素苷相对含量A=(OD520/0.1FW)(U·g)。
式中:OD为在 520 nm波长下测定提取液的OD值;FW为鲜叶片质量(g)。
1.3.3 叶片颜色参数
利用NR110色差仪测定叶片正面主脉与叶缘中间位置两侧脉间区域,测定明度值L*、色相值a*、色相值b*、色度值c*和色调角h* 5个参数。其中,L*是颜色明度和暗度的指标(取值范围为0~100);a*是红度和绿度的指标,其为+时表示偏红,为-时表示偏绿(取值范围为-128~127);b*是衡量黄蓝程度的指标,其为+时表示偏黄色,为-时表示偏蓝色,绝对值越大表示黄色或蓝色越深(取值范围为-128~127);c*是衡量色彩饱和度的指标,c*越大,颜色越纯;h*表示在色环中的颜色(孙彦琳等,2021)。
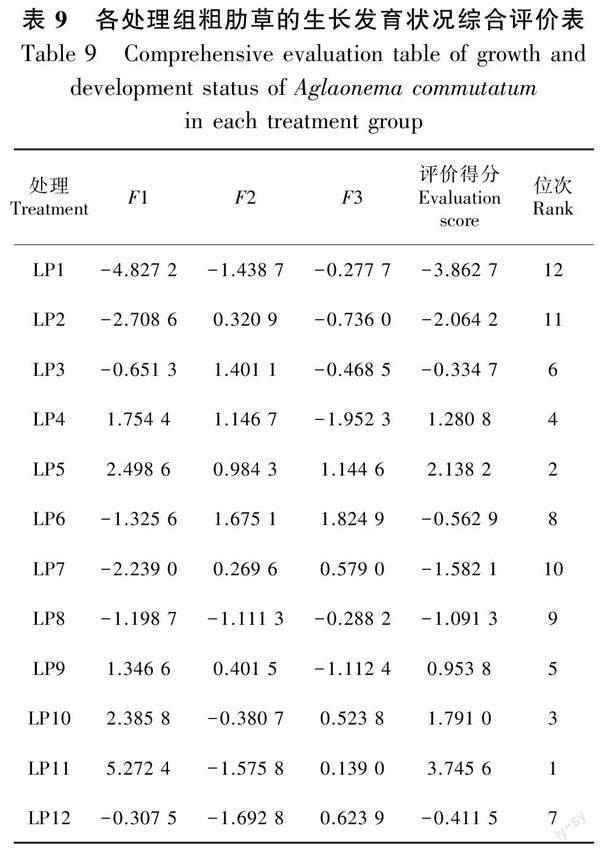
1.3.4 数据处理 采用Microsoft Excel 2016软件对数据进行整理汇总与制图,采用SPSS 22.0软件进行数据分析。双因素方差分析使用一般线性模型方法,定义固定因子和因变量,其中光质和光周期为固定因子,粗肋草生长、叶片色素和颜色指标为因变量。各处理之间采用Duncan检验法进行多重比较和差异显著性分析。对粗肋草生物量、叶片SPAD值和叶片颜色参数测量指标进行Pearson相关性分析;在这些指标间均具有一定的相关性的基础上,对与粗肋草主要生长性状及叶色性状相关的12个测量指标进行主成分分析。根据累积贡献率选出了前3个主成分,分别构建这3个主成分与原12项指标的线性组合方程,之后依据这3个主成分的方差贡献率,对这3个主成分进行线性加权从而构建粗肋草生长发育状况综合评价函数,用该函数计算各处理组粗肋草的综合评价得分,分值越高说明其改良效果越好(谢慈江等,2023;陶红等,2023)。
2 结果与分析
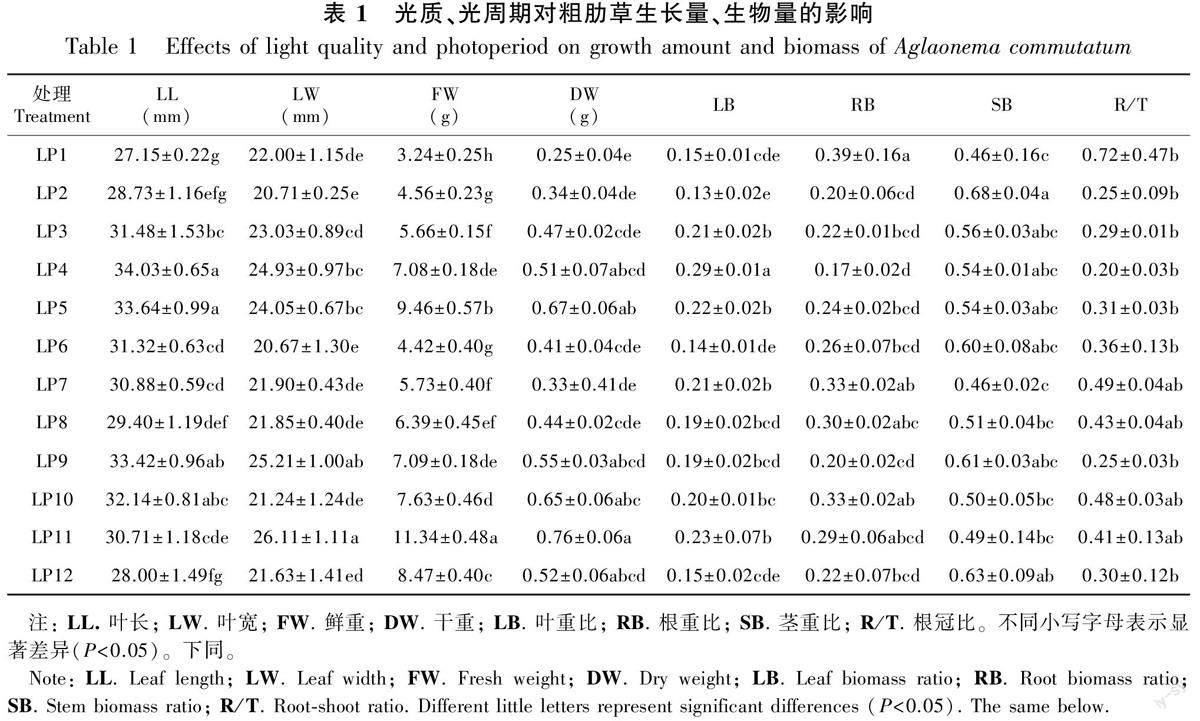
2.1 不同光质与光周期处理对粗肋草生长量、生物量的影响
2.1.1不同光质、光周期下粗肋草生长量、生物量的差异 在相同光周期下,不同光质对粗肋草生长量、生物量影响差异显著(表1)。当光周期为8 h·d-1时,LP4(R∶B)=2∶1光质下粗肋草叶长、叶宽和叶重比最大,LP2(R∶B)=1∶1光质下粗肋草茎重比最大。当光周期为12 h·d-1时,LP9(R∶B)=1∶2光质下粗肋草叶长最大,LP11(R∶B)=1∶3光质下粗肋草叶宽、叶重比最大,LP12(R∶B)=3∶1光质下粗肋草茎重比最大。当光周期为8、12 h·d-1时,根重比和根冠比均在白光条件下最大。
在相同光質下,12 h·d-1光周期粗肋草的鲜重、干重均大于8 h·d-1光周期的处理,说明12 h·d-1光周期处理更有利于粗肋草生物量的积累;粗肋草在8 h·d-1光周期和12 h·d-1光周期的鲜重和干重均在光质为R∶B=1∶3时达到最大值;其中12 h·d-1光周期条件下LP11处理的鲜重和干重最高,说明光质、光周期交互作用下LP11处理最有利于粗肋草生物量积累,在促进生物量积累中表现出优势。
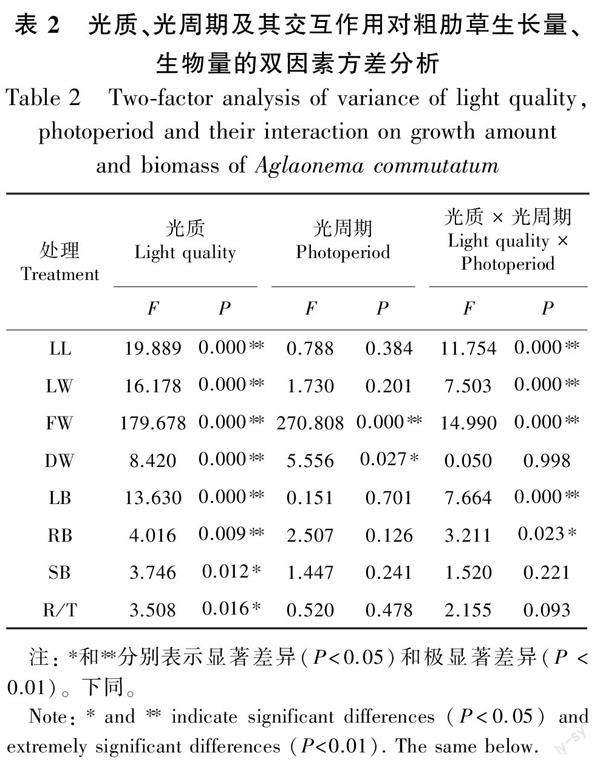
2.1.2 不同光质、光周期及其交互作用对粗肋草生长量、生物量的双因素方差分析 光质、光周期及光质×光周期交互作用对粗肋草叶片生长量、生物量的影响不同。光质对粗肋草叶长、叶宽、鲜重、干重、叶重比、根重比的影响极显著(P<0.01);光周期对粗肋草鲜重的影响达到极显著水平(P<0.01),对粗肋草干重的影响达到显著水平(P<0.05);光质×光周期交互作用对粗肋草叶长、叶宽、鲜重、叶重比的影响达到极显著水平(P<0.01),对根重比的影响达到显著水平(P<0.05),对干重、茎重比和根冠比影响不显著(P>0.05)(表2)。
2.2 光质与光周期对粗肋草叶片色素含量的影响
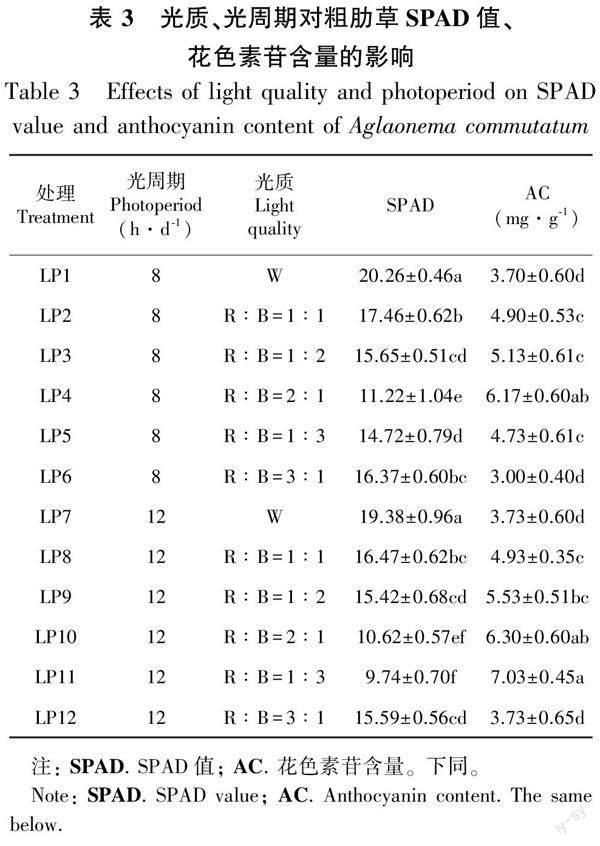
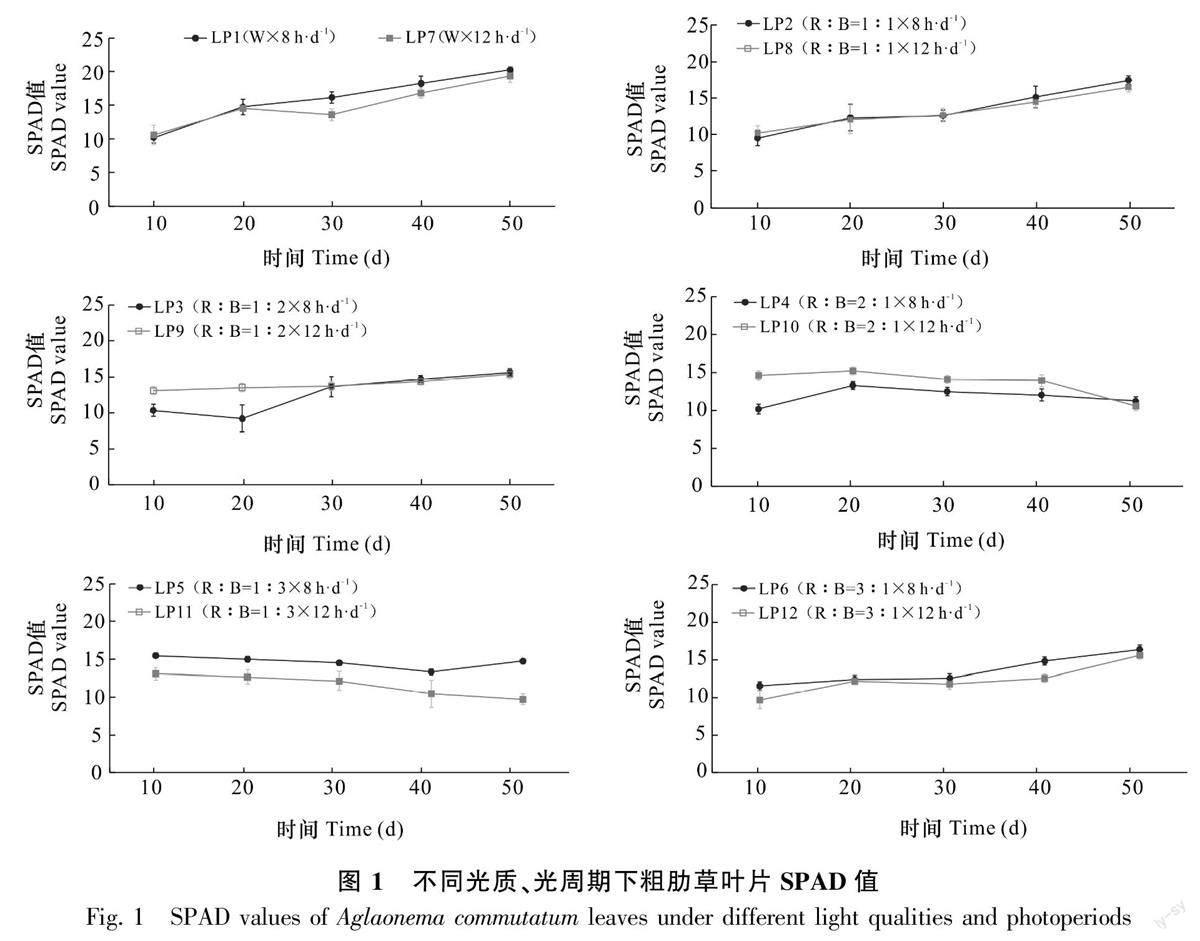
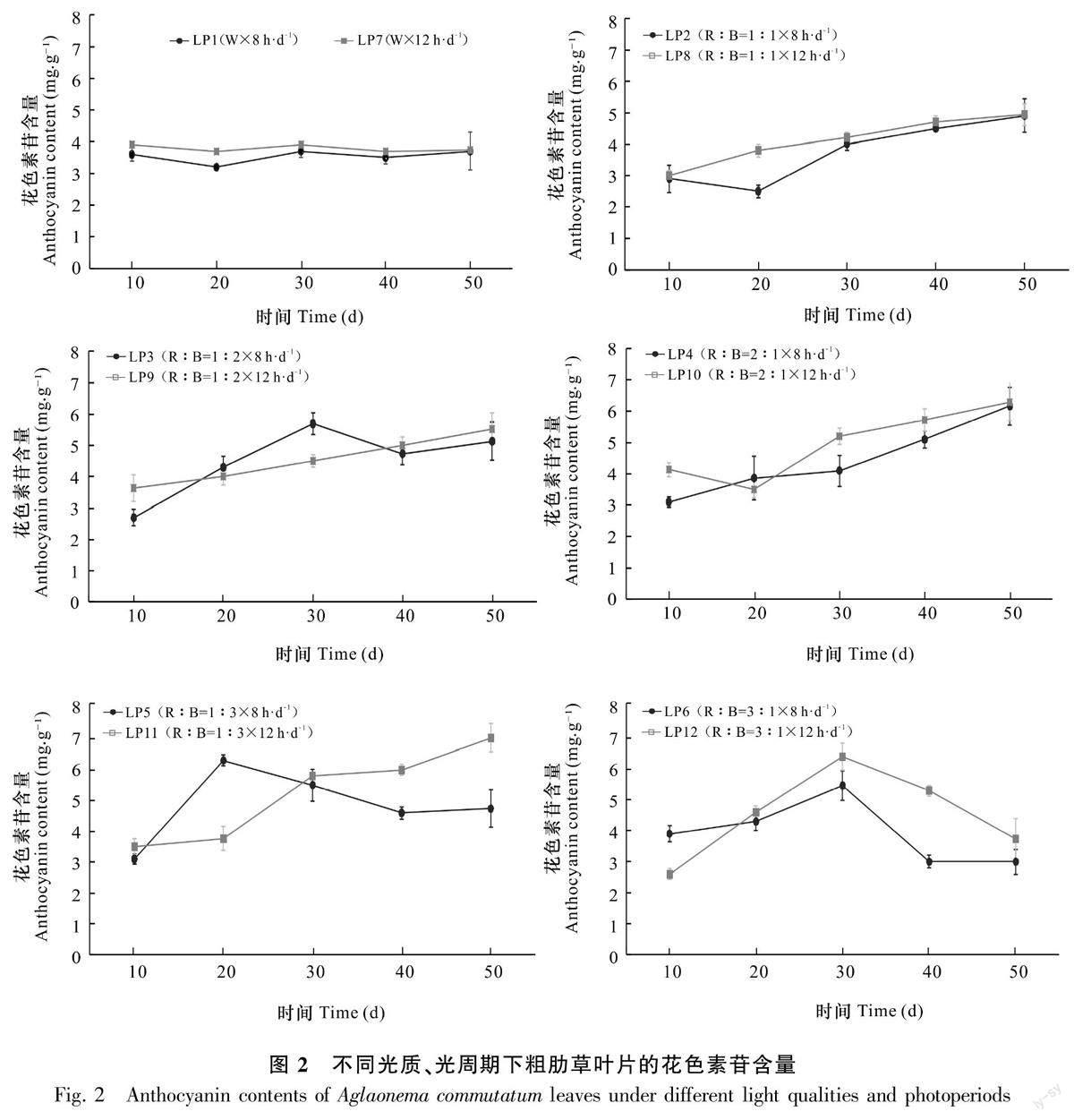
2.2.1不同光质、光周期下粗肋草叶片色素含量的差异 相同光周期培养50 d,不同光质对粗肋草SPAD值、花色素苷含量的影响差异显著(表3)。当光周期为8 h·d-1时,R∶B=2∶1光质下粗肋草叶片SPAD值最低,而此时粗肋草叶片花色素苷的含量最高。光周期为12 h·d-1时,R∶B=1∶3光质下粗肋草叶片SPAD值最低,花色素苷含量最高。在相同光质下,不同光周期对粗肋草叶片SPAD值、花色素苷含量的影响同样存在差异。在相同光质下,12 h·d-1光周期处理下粗肋草叶片SPAD值均低于8 h·d-1光周期的,而粗肋草叶片花色素苷含量均高于8 h·d-1光周期的。
光质×光周期交互作用下粗肋草叶片SPAD值、花色素苷含量存在显著差异。粗肋草叶片SPAD值按照从高到低依次为LP1>LP7>LP2>LP8>LP6>LP3>LP12>LP9>LP5>LP4>LP10>LP11;粗肋草叶片花色素苷含量按照从高到低依次为LP11>LP10>LP4>LP9>LP3>LP8>LP2>LP5>LP7>LP12>LP1>LP6;LP11处理的SPAD值最低,花色素苷含量最高。
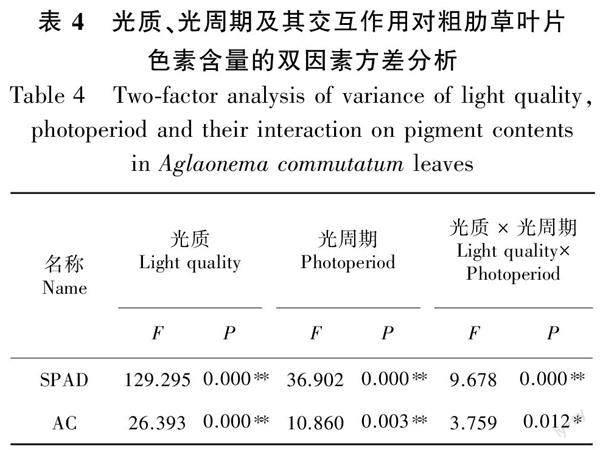
2.2.2 光质、光周期及其交互作用对叶片色素含量的双因素方差分析 由表4可知,光质、光周期均极显著影响粗肋草叶片SPAD值和花色素苷含量(P<0.01),而光质与光周期的交互作用对SPAD值的影响极显著(P<0.01),对花色素苷的影响显著(P<0.05)。
2.2.3 不同光质、光周期下粗肋草叶片SPAD值的变化 由图1可知,随着培养时间的延长,LP1、LP2、LP6、LP8、LP9处理的粗肋草叶片SPAD值逐渐增加,在R∶B=1∶1光质下,8 h·d-1光周期和12 h·d-1光周期粗肋草叶片SPAD值趋于一致,无明显变化;LP3、LP7、LP12处理的粗肋草叶片SPAD值存在先降低后逐渐上升趋势;LP10、LP11处理粗肋草叶片SPAD值逐渐降低,其中在LP11处理(R∶B=1∶3光质12 h·d-1光周期)下,粗肋草叶片SPAD值降幅均最大,粗肋草叶片SPAD值减少25.08%。
2.2.4 不同光质、光周期下粗肋草叶片花色素苷含量的变化 在白光光质下,8 h·d-1光周期和12 h·d-1光周期粗肋草叶片花色素苷含量趋于平稳,无明显变化。红蓝光质组合中LP4、LP8、LP9、LP11处理下粗肋草叶片花色素苷含量随培养时间的增加而逐渐增加。培养50 d,不同光质处理中LP11的粗肋草叶片花色素苷含量增加最高,增加了100.95%,其次是LP4处理,粗肋草叶片花色素苷含量增加了98.92%;LP2、LP5、LP10处理的粗肋草叶片花色素苷含量除在培养20 d突然降低外,整体呈上升趋势;LP6、LP12处理的粗肋草叶片花色素苷含量随时间的变化先增加后降低(图2)。
2.3 不同光质与光周期对粗肋草叶片颜色参数的影响
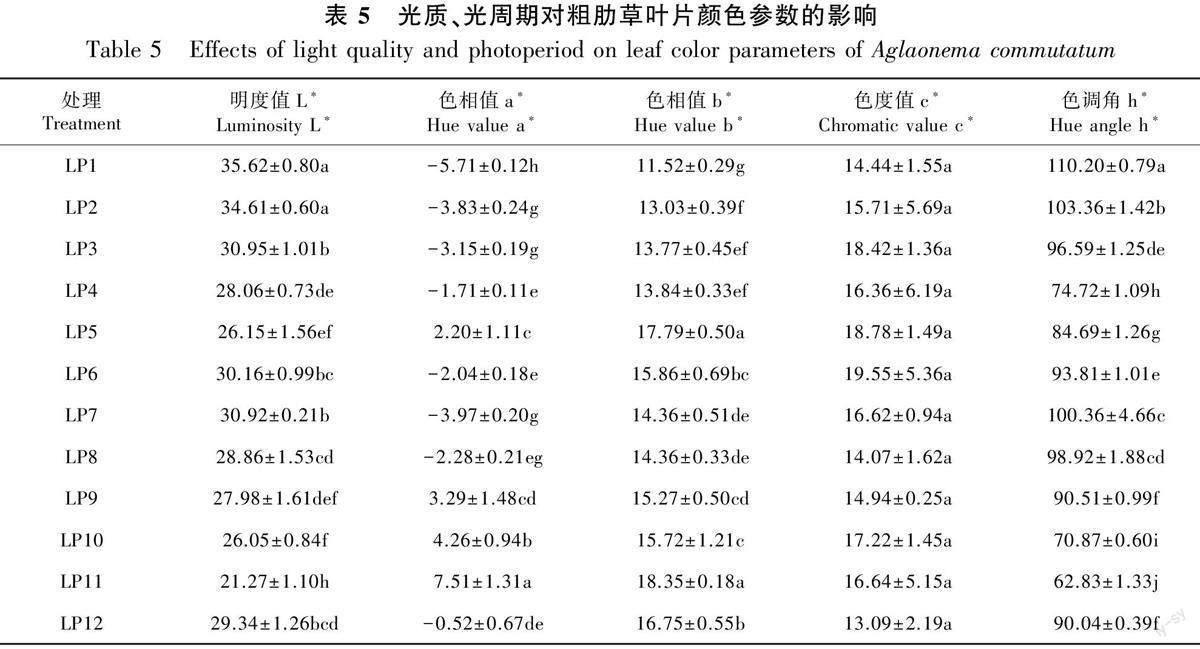
2.3.1 不同光质、光周期影响下粗肋草叶片颜色参数 由表5可知,不同光质对粗肋草叶片明度值L*、色相值a*、色相值b*、色调角h*的影响存在显著差异,对色度值c*影响不显著。8 h·d-1光周期和12 h·d-1光周期处理,R∶B=1∶3光质下粗肋草叶片的色相值a*、b*均最高。在相同光质下,不同光周期对粗肋草叶片色相值a*、b*的影响同样存在差异。在相同光质下,12 h·d-1光周期处理下粗肋草叶片色相值a*、b*均高于8 h·d-1光周期的。粗肋草叶片色相值a*从大到小依次为LP11>LP10>LP9>LP5>LP12>LP4>LP6>LP8>LP3>LP2>LP7>LP1,粗肋草叶片色相值b*从大到小依次为LP11>LP5>LP12>LP6>LP10>LP9>LP7>LP8>LP4>LP3>LP2>LP1。在相同光质下,12 h·d-1光周期处理下粗肋草叶片色调角h*均小于8 h·d-1光周期处理下粗肋草叶片色调角h*;在光质与光周期交互作用下,粗肋草叶片色调角h*从小到大依次为LP11<LP10<LP4<LP5<LP12<LP9<LP6<LP3<LP8<LP7<LP2<LP1,LP11处理下叶片色调角h*最低,此时叶片偏向橙色(40°<h*≤75°)。
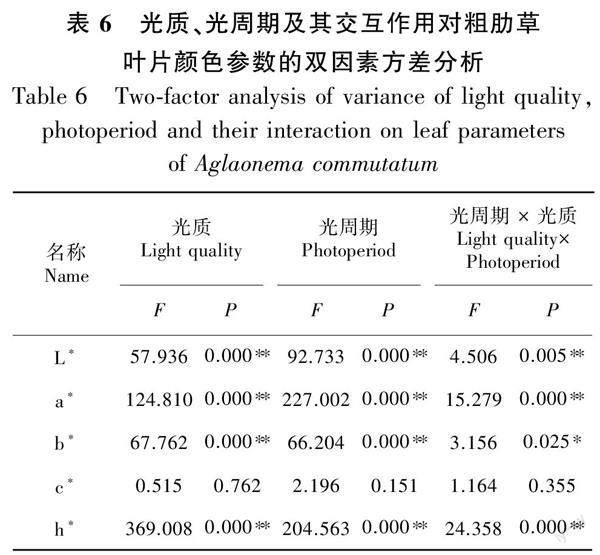
2.3.2 不同光质、光周期及其交互作用对叶片参数的雙因素方差分析 基于不同光质与光周期的双因素方差分析结果(表6)表明,本试验中光质、光周期及光质×光周期交互作用对粗肋草叶片颜色参数的影响不同。可见,光质、光周期及光质×光周期交互作用对粗肋草叶片颜色参数L*、a*、b*、h*的影响均极显著(P<0.01),而对色度值c*的影响均不明显(P>0.05)。
2.4 粗肋草生物量、叶片色素含量和叶片颜色参数的相关性分析和主成分分析
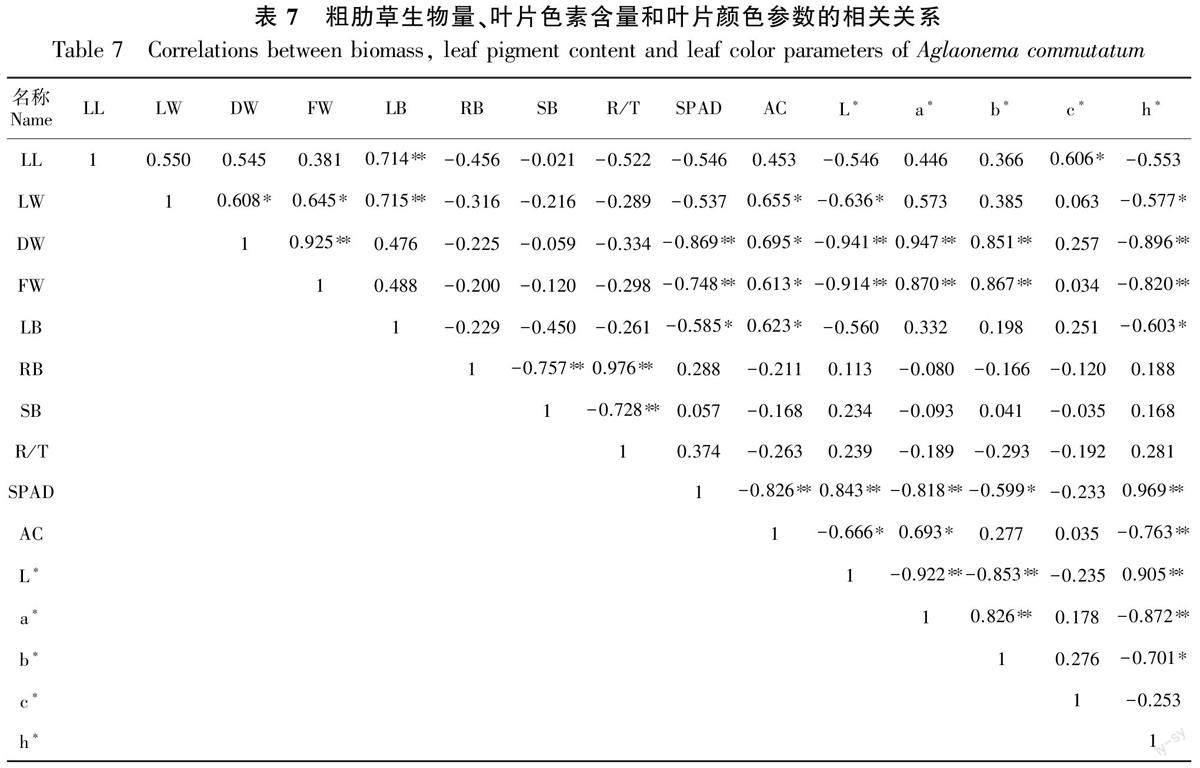
2.4.1 粗肋草生物量、叶片色素含量和叶片颜色参数的相关性分析 粗肋草生物量、叶片色素含量和叶片颜色参数存在相关性:粗肋草叶长与叶重比呈极显著正相关,与色度值c*呈显著相关,相关系数分别为0.714和0.606;粗肋草叶宽与干重、鲜重、叶重比和花色素苷含量均呈显著正相关,与明度值L*和色调角h*呈显著负相关;粗肋草干重和鲜重均与明度值L*呈极显著负相关,与花色素苷含量、色相值a*和色相值b*呈显著正相关;叶片SPAD值与干重、鲜重、叶重比、花色素苷含量、色相值a*和色相值b*呈显著负相关,与明度值L*和色调角h*呈显著正相关(表7)。
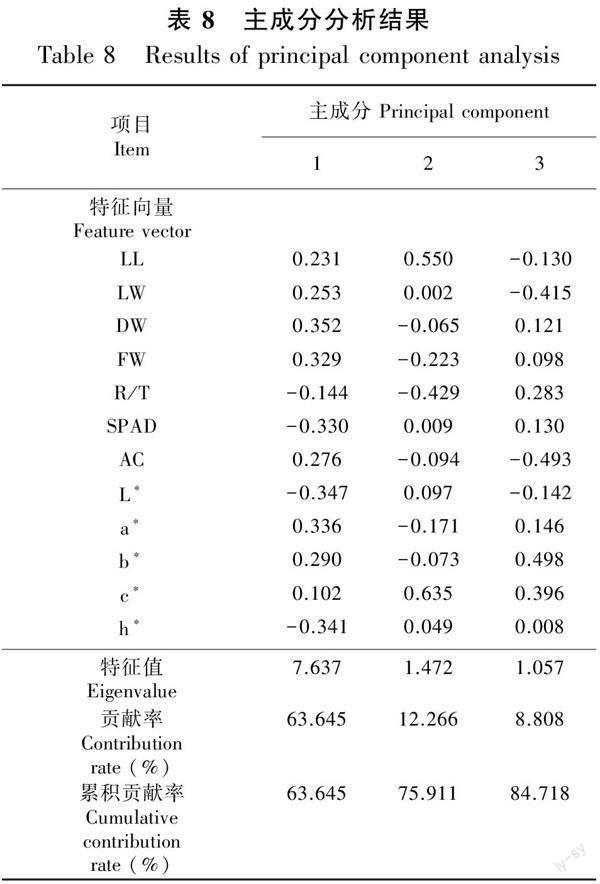
2.4.2 粗肋草生长性状和叶色性状相关指标的主成分分析 对粗肋草12个指标的主成分分析结果(表8)表明,前3个主成分的贡献率分别为63.645%、12.266%和8.808%,累积贡献率达到84.719%,从而表明这3个综合指标可以反映原指标的绝大部分信息,可以代替原来12个单项指标对不同处理下粗肋草生长发育状况进行综合评价,并且各综合指标所代表的信息不再重叠。根据各综合指标的标准化特征向量值及各单项指标的标准化值(Xi),可得到粗肋草生长发育指标的3个主成分与原12项指标的线性组合方程:F1=0.231X1+0.253X2+0.352X3+0.329X4-0.144X5-0.330X6+0.276X7-0.347X8+0.336X9+0.290X10+
0.102X11-0.341X12;F2=0.550X1+0.002X2-0.065X3-0.223X4-0.429X5+0.009X6-0.094X7+0.097X8-0.171X9-0.073X10+0.635X11+0.049X12;F3=-0.130X1-0.415X2+0.121X3+0.098X4+0.283X5+0.130X6-0.493X7-0.142X8+0.146X9+0.498X10+0.396X11+0.008X12。
2.4.3 不同光质、光周期下粗肋草生长发育状况的综合评价 以各主成分对应的方差贡献率作为权重,对主成分得分和相应的权重进行线性加权,构建粗肋草的生长发育状况评价函数为F=0.751F1+0.145F2+0.104F3,计算不同处理下粗肋草的生长发育状况综合评价得分,分值越高表示该处理下粗肋草的生长发育状况越好。本研究结果表明,在所有处理中,LP11的生长发育状况综合评价得分最高,LP1的生长发育状况综合评价得分最低。由此可见,光质与光周期交互作用下LP11(R∶B=1∶3×12 h·d-1)处理对促进粗肋草生长发育效果最好(表9)。
3 讨论
3.1 光质、光周期对粗肋草生长的影响
光环境对植物的影响贯穿植物的整个生命周期。本研究发现光质显著影响粗肋草生长量及生物量的积累,光周期对粗肋草鲜重、干重的影响显著,在相同光质下,光周期为12 h·d-1的粗肋草鲜重、干重比8 h·d-1光周期培养的更大,与12 h·d-1光周期更有利于萝卜(Raphanus sativus)幼苗鲜重、干重积累的研究报道是一致的(刘文科和姜偲倩,2016)。这可能是不同植物对日照长短的需求不同,延长光周期可以提高植物光系统Ⅱ反应中心的活性(姚宁等,2022),促进植物光能利用效率;光周期变短会降低植物的原初光能转化效率,使叶片的转化能力降低,从而導致植株生物量的差异。红蓝光质不同组合对粗肋草生物量积累均有促进作用,不同红蓝光质配比与不同光周期交互作用提高植物生长量的效果不同。8 h·d-1光周期和12 h·d-1光周期的粗肋草鲜重、干重均在光质R∶B=1∶3达到最大值,表明红光和蓝光比例为1∶3时最能促进粗肋草生物量的积累。不同光质对植物光合作用的影响不同,红蓝光处理下植株叶片的气孔导度、光系统Ⅱ的实际光化学效率、净光合速率等均高于单色光的处理(任毛飞等,2023)。蓝光的非光化学猝灭系数最高,其光系统Ⅱ在暗适应和光适应下的最大光化学效率等叶绿素荧光特性均高于其他单色光源(Gao et al., 2020);光系统Ⅱ的适宜光源,能促进细胞分裂和生长(Li et al.,2020)。因此,粗肋草生物量在蓝光比例最高的光质配比处理下积累最多。
3.2 光质、光周期对粗肋草叶片色素含量和颜色的影响
本研究中光照时间和光质均显著影响粗肋草叶片SPAD值的变化,其中白光8 h·d-1光周期条件下的粗肋草SPAD值最高。SPAD值与叶绿素总含量、叶绿素a、叶绿素b含量呈显著正相关(项倩等,2022)。叶绿素作为绿色植物生长发育的指标之一,不同光质配比和光周期调控绿色植物光合色素的形成。对杉木(Cunninghamia lanceolata)(汪星星等,2022)和西瓜(Citrullus lanatus)(秦伟等,2018)的研究均发现白光下幼苗的叶绿素含量比红蓝光组合处理高。白光更有利于粗肋草叶绿素的合成,但光照时间过长,会降低叶绿素合成。这可能是植物面对光周期不足的环境时可以通过提高光合色素合成来增强光合作用抵抗逆境;光周期过长会对植物叶绿素含量产生逆境效应,阻碍叶绿素合成;同时,植物具有表型可塑性,可以随着光环境的变化调整其形态和光合生理,以提高对光能捕捉的能力和利用效率(Lei et al., 2013)。
在相同红蓝光质配比条件下,光周期12 h·d-1更有利于粗肋草叶片花色素苷含量提高,当红蓝光质为R∶B=1∶3时,花色素苷含量最高。这可能与促进花色素苷合成最有效的是蓝光和紫外光有关(高飞等,2014)。光作为一种环境因子,其调控花色素苷合成主要通过对相关酶基因的直接或间接调控来完成(Jaakola, 2013)。红光可以显著增加草莓果实花色素苷的含量和相关酶的活性(Miao et al., 2016);蓝紫光可以诱导茶树花青苷关键合成酶查尔酮合成酶、类黄酮-3-羟化酶和二氢黄酮醇还原酶的表达,红光调控花色素苷合成酶(李智,2014)。光照条件为16 h·d-1的甘薯(Ipomoea batatas)叶片类黄酮合成途径中结构基因的表达量和花色素苷含量比8 h·d-1的高(魏永赞等,2017)。因此,在长时间光照条件下,蓝光为粗肋草的花色素苷合成和转录调控的主要效能光质,以信号转导途径调控花色素苷合成和转运相关基因的高效表达,加强花色素苷的积累。本研究中不同处理下粗肋草的L*均为正值,L*与花色素苷含量呈显著负相关,与SPAD值呈极显著正相关;色相值a*与花色素苷含量呈显著正相关,与SPAD值呈显著负相关,这与郭欢欢等(2017)对黄连木(Pistacia chinensis)叶色研究的结果一致。不同红蓝光质×光周期组合下培养的粗肋草叶片SPAD值和花色素苷的色素含量不同,叶片呈现的颜色也不同,其中LP11处理的粗肋草叶片花色素苷含量最高,SPAD值最低,L*最小,色度值a*、b*最大,实际观察到此处理的粗肋草叶色偏向橙红色,推测光质和光周期对粗肋草叶片色泽的影响,主要通过提高叶片花色素苷含量和降低叶绿素含量的变化来实现。在粗肋草设施栽培中可以通过控制光周期和红蓝光在光源中所占比例,影响叶片花色素苷和叶绿素的含量,从而改变叶色。
4 结论
本试验从不同光质与光周期交互作用对粗肋草叶片颜色影响的角度,证明了不同光质和光周期交互作用对粗肋草叶片颜色影响显著。在相同光质下,12 h·d-1光周期比8 h·d-1光周期对提高粗肋草生物量和花色素苷含量、降低叶绿素含量的作用更显著。从提高植物生长量和改良叶色角度来考虑,光质与光周期交互作用下LP11(R∶B=1∶3×12 h·d-1)处理对粗肋草生长量、生物量和叶片颜色的改变最大,为最佳光质和光周期组合。
参考文献:
CHEN G, 2020. Effects of different light quality ratios on photosynthetic characteristics of Cunninghamia lanceolata seedling Clones [D]. Fuzhou: Fujian Agriculture and Forestry University.[陈钢, 2020. 不同光质配比对杉木幼苗优良无性系光合特性的影响 [D]. 福州: 福建农林大学.]
GAO F, KE L, JIN T, et al., 2014. Effects of light on anthocyanin synthesis in plants [J]. Chin Agric Sci Bull, 30(34):6-10.[高飞, 柯燚, 金韬, 等, 2014. 光照对植物合成花色素苷的影响研究进展 [J]. 中国农学通报, 30(34): 6-10.]
GAO S, LIU XN, LIU Y, et al., 2020. Photosynthetic characteristics and chloroplast ultrastructure of Welsh onion (Allium fistulosum L.) grown under different LED wavelengths [J]. BioMed Cent, 2: 78.
GUO HH, LIU Y, YAO F, et al., 2017. Relationship between pigment contents and leaf color parameters of Pistacia chinensis Bunge of different provenances in autumn [J]. Acta Bot Boreal-Occident Sin, 37(10):2003-2009.[郭欢欢, 刘勇, 姚飞, 等, 2017. 不同种源黄连木秋季色素含量与叶色参数的关系 [J]. 西北植物学报, 37(10): 2003-2009.]
GUO LY, SHI M, WU YF, et al., 2018. Shading responses to leaf color and physiology during discoloration period of Carpinus betulus [J]. J Cent S Univ For Technol, 38(8):26-34.[郭力宇, 施曼, 吴驭帆, 2018. 遮阴对欧洲鹅耳枥变色期叶色及生理的影响 [J]. 中南林业科技大学学报, 38(8):26-34.]
HAYAMA R, COUPLAND G, 2004. The molecular basis of diversity in the photoperiodic flowering responses of Arabidopsis and rice [J]. Plant Physiol, 135(2):677-684.
HU SQ, 2018. Effects of supplemental light quality and durations of illumination on growth, physiological characteristics of Lilium oriental [D]. Hangzhou: Zhejiang University.[胡绍泉, 2018. 不同光质及补光时间对东方百合生长及生理特性的影响 [D]. 杭州: 浙江大学.]
JAAKOLA L, 2013. New insights into the regulation of anthocyanin biosynthesis in fruits [J]. Trends Plant Sci, 9(18):477-483.
KIM HH, GOINS GD, WHEELER RM, et al., 2004. Stomatal conductance of lettuce grown under or exposed to different light qualities [J]. Ann bot, 94: 691-697.
LEI G, BO L, LIU WY, et al., 2013. Inhibition effects of daughter ramets on parent of clonal plant Eichhornia crassipes[J]. Aquatic Bot, 107: 47-53.
LI DM, ZHAN QC, HUANG D, et al., 2022. A new Aglaonema cultivar ‘Baibaoshi’[J]. Acta Hortic Sin, 49(S1):157-158.[李冬梅, 詹启成, 黄丹, 等, 2022. 粗肋草属花卉新品种‘白宝石’ [J]. 园艺学报, 49(S1):157-158.]
LI L, 2016. Study on the formation and regulation of leaf color from Acer rubrum L. [D]. Chongqing: Southwest University.[李力, 2016. 北美红枫呈色生理机制及叶色调控 [D]. 重庆: 西南大学.]
LI LS, ZHAI YM, ZHANG L, 2022. Effect of light regulation on callus induction system of test-tube plantlets of Aglaonema commutatum Schott [J]. Mol Plant Breed, 20(14):4755-4764.[李林山, 翟懿銘, 张黎, 2022. 光调控对粗肋草试管苗愈伤组织诱导的影响 [J]. 分子植物育种, 20(14): 4755-4764.]
LI YF, LI L, LIU JG, et al., 2020. Light absorption and growth response of Dunaliella under different light qualities [J]. J Appl Phycol, 32 (2):1041-1052.
LI Z, 2014. Effect of the main environmental factors on anthocyanin content and related genes expression of purple tea shoots [D]. Taian: Shandong Agricultural University. [李智, 2014. 不同环境因子调控茶树紫色芽叶形成的分子机制研究 [D]. 泰安: 山东农业大学.]
LIU MZ, LI Q, YANG C, et al., 2021. Effect of LED light quality on growth and development and chlorophyll fluorescence characteristics of red orange seedlings [J]. S Chin Fruits, 50(2): 1-7. [刘敏竹, 李强, 杨超, 等, 2021. LED光质对红桔幼苗生长发育和叶绿素荧光特性的影响 [J]. 中国南方果树, 50(2): 1-7.]
LIU Q, 2015. Effects of different photoperiods and different light quality on the physiological characteristic and quality in strawberry [D]. Taian: Shandong Agricultural University.[刘庆, 2015. 不同光周期及光质对草莓生理特性及品质的影响 [D]. 泰安: 山东农业大学.]
LIU WK, JIANG SQ, 2016. Effects of light quality and intensity of LEDs on growth and biomass accumulation of radish seedlings [J]. Chin Light Light, 381(12):24-26.[刘文科, 姜偲倩, 2016. LED红蓝光质及其光周期对萝卜苗生长及生物量的影响 [J]. 中国照明电器, 381(12): 24-26.]
MIAO LX, ZHANG YC, YANG XF, et al., 2016. Colored light-quality selective plastic films affect anthocyanin content, enzyme activities, and the expression of flavonoid genes in strawberry (Fragaria × ananassa) fruit [J]. Food Chem, 207: 93-100.
MOHAMED NMA, 2016. The effects of light intensity and time on plant growth [D]. Fuzhou: Fujian Agricultural and Forestry University.[ MOHAMED NMA, 2016. The effects of light intensity and time on plant growth [D]. 福州: 福建农林大学.]
QIN W, CHEN K, ZHAO YF, et al., 2018. Effects of different LED light sources on SPAD value, photosynthetic parameters and physiological quality of watermelon seedlings [J]. Tianjin Agric Sci, 24(11): 8-10.[秦伟, 陈昆, 赵跃锋, 等, 2018. 不同LED光源对西瓜幼苗SPAD值、光合参数及生理品质的影响 [J]. 天津农业科学, 24(11): 8-10.]
REN MF, MAO GL, LIU SZ, et al., 2023. Research progress on the effects of light quality on plant growth and development, photosynthesis, carbon and nitrogen metabolism [J]. Acta Phytophysiol Sin,59(7):1221-1228.[任毛飞, 毛桂玲, 刘善振, 等, 2023. 光质对植物生长发育、光合作用和碳氮代谢影响的研究进展 [J]. 植物生理学报, 59(7): 1221-1228.]
SHAFIQ I, HUSSAIN S, RAZA MA, et al., 2021. Crop photosynthetic response to light quality and light intensity [J]. J Integr Agric, 20(1):4-23.
SUN YL, CHENG BX, ZHOU LJ, et al., 2021. Studies on the correlation between leaf coloration and flower coloration in F1 population of Rosa spp. [J]. J Sichuan Agric Univ, 39(2):173-179.[孫彦琳, 程璧瑄, 周利君, 等, 2021. 现代月季F1代群体叶色与花色相关性研究 [J]. 四川农业大学学报, 39(2): 173-179.]
TAO H, HANG H, FENG TM, et al., 2023. Adaptability evaluation of Prunus persica cultivars in Gansu provincebased on principal component analysis [J]. Non-Wood For Res, 41(1): 236-244. [陶红,韩宏,冯廷敏,等,2023. 基于主成分分析的不同碧桃品种在甘肃地区的适应性评价 [J]. 经济林研究,41(1): 236-244.]
WANG DX, SUN HJ, DE YJ, et al., 2019. Change of leaf color of Liquidambar formosana seedlings under different light quality treatments [J]. For Res, 32(4):158-164.[王冬雪, 孫海菁, 德永军, 等, 2019. 不同光质处理对枫香幼苗叶色的影响 [J]. 林业科学研究, 32(4): 158-164.]
WANG SA, WANG XD, SHI XB, et al., 2016. Red and blue lights significantly affect photosynthetic properties and ultrastructure of mesophyll cells in senescing grape leaves [J]. Hortic Plant J, 2(2):82-90.
WANG SY, LIU JM, ZHAO CY, et al., 2017. Chromosome numbers analyses of Aglaonema cultivars [J]. Chin J Trop Crops, 38(3):456-462.[王树茵, 刘金梅, 赵超艺, 等, 2017. 粗肋草品种的染色体数目分析 [J]. 热带作物学报, 38(3): 456-462.]
WANG XX, CHEN G, CAO GQ, et al., 2022. Effect of different photo qualities on antioxidase activity and chlorophyll content of Chinese fir seedlings [J]. J Gansu Agric Univ, 57(4):137-146.[汪星星, 陈钢, 曹光球, 等, 2022. 不同光质对杉木幼苗抗氧化酶活性和叶绿素含量的影响 [J]. 甘肃农业大学学报, 57(4):137-146.]
WEI QM, 2018. Effect of LED light quality and photoperiods on growth and photosynthetic characteristics of Paramichelia baillonii seedlings [D]. Nanning: Guangxi University.[韦秋梅, 2018. LED不同光质及光周期对山白兰苗木生长及光合特性的影响 [D]. 南宁: 广西大学.]
WEI YZ, LI WC, DONG C, et al., 2017. Regulation and mechanism of light on anthocyanin biosynthesis in plants [J]. Acta Phytophysiol Sin, 53(9):1577-1585.[魏永赞, 李伟才, 董晨, 等, 2017. 光照对植物花色素苷生物合成的调控及机制 [J]. 植物生理学报, 53(9): 1577-1585.]
XIANG Q, WU L, XU RH, et al., 2022. Correlation between SPAD and chlorophyll content in infected tomato leaves at different temperatures [J]. Acta Hortic Sin, 513(18):8-15.[项倩, 吴磊, 徐若涵, 等, 2022. 不同温度下染病番茄叶片SPAD和叶绿素含量的相关性 [J]. 北方园艺, 513(18): 8-15.]
XIE CJ, HE FY, LIU L, et al., 2023. Effects of light quality and photoperiod on the growth and physiology of Paramichelia baillonii seedlings [J/OL]. Guihaia: 1-14 [2023-07-26]. http://kns.cnki.net/kcms/detail/45.1134.Q.20221227.1646.008.html. [谢慈江, 何福英, 刘莉, 等, 2023. 光质和光周期对山白兰苗木生长、生理的影响 [J/OL]. 广西植物: 1-14 [2023-07-26]. http://kns.cnki.net/kcms/detail/45.1134.Q.20221227.1646.008.html.]
YAO N, LIU JF, JIANG ZP, et al., 2022. Effects of photoperiod and light quality on seedling growth and chlorophyll fluorescence kinetics of Quercus L. [J]. For Res, 35(1):59-69.[姚宁, 刘建锋, 江泽平, 等, 2022. 光周期与光质对栎属幼苗生长及叶绿素荧光的影响 [J]. 林业科学研究, 35(1): 59-69.]
ZHANG SM, PENG YY, LI L, 2016. Effect of different light quality treatments of leaf color change of Selaginella uncinata [J]. Acta Hortic Sin, 363(12):75-79.[张水木, 彭媛媛, 李林, 2016. 不同光质处理对翠云草叶色变化的影响 [J]. 北方园艺, 363(12): 75-79.]
ZHAO MW, 2020. Effects of different temperature, light quality and light time on the growth and quality of ice plant [D]. Shihezi: Shihezi University.[赵明伟, 2020. 不同温度、光质和光照时间对冰菜生长及品质的影响 [D]. 石河子: 石河子大学.]
ZHOU ZP, SHI SL, SONG T, et al., 2018. Optimization of callus and multiple shoots induction medium of Aglaonema commutatum Schott ‘Red Valentine’[J]. Genom Appl Biol, 37(12):5429-5436.[周佐葡, 施苏丽, 宋婷, 等, 2018. 如意皇后粗肋草愈伤组织及丛生芽诱导培养基优化 [J]. 基因组学与应用生物, 37(12): 5429-5436.]
(责任编辑 邓斯丽 蒋巧媛)
猜你喜欢
湖北农业科学(2024年6期)2024-07-04 12:59:05
遗传(2023年9期)2023-09-25 09:31:44
农业科技通讯(2023年1期)2023-02-12 07:08:50
四川林业科技(2022年1期)2022-03-22 07:48:32
上海农业学报(2022年1期)2022-03-08 00:09:12
浙江农业学报(2019年7期)2019-08-14 01:26:12
浙江农业学报(2018年9期)2018-10-11 07:26:22
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
上海蔬菜(2017年3期)2017-08-28 09:43:18
