
乌鲁木齐达坂城山区地衣生态位特征的研究
2023-11-01 04:51:20李作森艾尼瓦尔·吐米尔
广西植物 2023年9期
李作森 艾尼瓦尔·吐米尔







摘 要:為揭示达坂城山区地衣对生境资源的利用状况及竞争程度,该研究在野外设置30个样方并调查其物种盖度数据,利用生态位宽度、生态位重叠指数及排序分析,探讨达坂城山区地衣的生态位特征及环境影响因子。结果表明:(1)达坂城山区生态位最宽的为丽黄鳞衣(Rusavskia elegans)和内卷野粮衣(Circinaria contorta),对不同环境具有较强的适应性,其他地衣物种生态位宽度较窄,对环境资源利用程度低。(2)物种间生态位重叠值整体偏低,生态位重叠值较高的物种对极少,地衣物种生态位分化程度高,物种间竞争不激烈。(3)生态位较宽的物种与多数物种之间都存在生态位重叠,但重叠值较低;一些生态位较窄的物种与其他物种之间的生态位重叠值较高;生态位重叠和生态位宽度之间并无直接的线性关系。(4)地衣物种沿海拔梯度分布具有差异,海拔、光照强度、湿度、风速和人为干扰是本地区地衣群落物种生态位特征差异的主导环境因子。综上所述,达坂城山区地衣物种由于生境资源的竞争占据各自独特的生态位,形成了生态位的分化,群落较为稳定,沿海拔梯度的生境条件差异是物种差异分布的重要原因。该研究可为地衣群落构建研究提供理论依据,并对该地区地衣物种的多样性和生境保护具有重要意义。
关键词: 地衣, 生态位宽度, 生态位重叠, 海拔梯度, 达坂城山区
中图分类号:Q948
文献标识码:A
文章编号:1000-3142(2023)09-1636-10
收稿日期:2022-05-07
基金项目:国家自然科学基金(32160046,31660009)。
第一作者: 李作森(1996-),硕士研究生,主要从事地衣生态学研究,(E-mail)751886659@qq.com。
*通信作者:艾尼瓦尔·吐米尔,博士,教授,主要从事地衣生态学研究,(E-mail)anwartumursk@xju.edu.cn。
Niche characteristics of lichen in Dabancheng
mountainous area, Urumqi
LI Zuosen, TUMUR Anwar*
( College of Life Sciences and Technology, Xinjiang University, Urumqi 830046, China )
Abstract:In order to reveal the status of habitat resource utilization and the degree of competition among lichens in Dabancheng mountainous areas. In this work, 30 plots were sampled in the field and the species cover data of the plots were investigated, and the niche characteristics and environmental factors of lichen community species were analyzed by using niche width, niche overlap index and ordination analysis in this Dabancheng mountainous area. The results were as follows: (1) The niche width of Rusavskia elegans and Circinaria contorta were the widest in Dabancheng mountainous area, indicating that they were more adaptable to different environments. The niche widths of other lichen species were narrow, indicating that the species used the environmental resources to a less extent. (2) The niche overlap value between species was generally low, with only a few species pairs having a high niche overlap value. The degree of niche differentiation between lichen species was high, indicating that the competition between species was not intense. (3) Although niche overlap was common among species with wider niche widths, niche overlap values were lower. However, some species with narrower niche widths had higher niche overlap values with other species. As a result, there was no clear linear relationship between niche overlap and niche width. (4) The distribution of lichen species varied along the altitudinal gradient, and altitude, light intensity, humidity, wind speed, and human disturbance were the dominant environmental factors for the different niche characteristics of lichen community species in the study area. In conclusion, as a result of habitat resource competition in Dabancheng mountainous area, lichen species occupied their own distinct niches, resulting in niche differentiation and relatively stable communities. The distribution of species differed significantly due to differences in habitat conditions along the altitudinal gradient. The results of this paper provides a theoretical basis for the study of lichen community assembly, which are of great significance to the conservation of lichen species diversity and habitat in the region.
Key words: lichen, niche width, niche overlap, altitudinal gradient, Dabancheng mountainous area
生态位理论一直是群落生态学的主流理论,对物种生态位特征的研究能够反映物种在群落中的地位和作用。Johnson(1910)首次使用“生态位”概念,但并未定义生态位,Grinnell(1917)正式定义生态位为不同的种或亚种在不同环境中所占据的位置特征。国外学者开始系统地研究生态位的定义和定量计测方法(Gause, 1934; Keddy, 1992; Vannette & Fukami, 2014),对生态位理论进行了多方面的阐述和验证,认为生态位分化是物种共存的基础。尽管生态位的定义、量化和计测方法在生态学研究进程中经历诸多波折,但生态位理论在群落生态学的研究中已取得了诸多成果(Sales et al., 2021;牛克昌等,2022),是研究群落物种共存和多样性维持机制的重要理论。
生态位的定量研究主要是通过计算生态位宽度和生态位重叠值,反映物种在群落中的资源利用能力和相似程度(庞久帅等,2022)。目前,国内外有大量研究量化分析群落中物种占据的空间范围和利用生境资源的能力,从而探讨群落物种共存的生态位机制,多集中于草本、灌木、乔木和苔藓等高等植物群落(陈旭等,2009;Sun et al., 2021;曹华等,2021;庞久帅等,2022)。早期关于地衣群落生态学的研究,主要集中于用传统的物种多样性指数探讨群落物种分布格局。此外,由于其特殊的结构,对环境及气候变化的高度敏感,一些研究通过地衣物种多样性和空气污染、环境气候变化及干扰之间的关系来评估森林、城市、保护区等区域的污染程度、气候变化及干扰程度的情况(Lubek et al., 2018;Koch et al., 2019;Lucheta et al., 2019)。随着生态位特征量化方法在高等植物中的广泛应用,此方法逐渐应用于地衣,但研究较少(艾尼瓦尔·吐米尔等,2012;Giordani et al., 2015;靳文婷和艾尼瓦尔·吐米尔,2017;Rolshausen et al., 2018;李作森等,2021;田雅楠和艾尼瓦尔·吐米尔,2021)。田雅楠和艾尼瓦尔·吐米尔(2021)通过计算地衣生态位宽度和重叠值,分析了博格达峰北坡地衣生态位特征,发现由于资源利用能力的差异导致地衣形成了生态位分化。拜合提妮萨·依明等(2022)分析了天山一号冰川岩面生地衣的生态位特征,表明物种资源利用能力差异较小,物种间存在普遍的竞争关系。综上表明,地衣物种生态位可能受种间竞争和环境作用的影响,但并未结合具体的环境因子分析物种生态位特征,而种群生态位受人类活动等生物因子和非生物环境因子的影响(孙成等,2022),并且地衣对外界环境变化敏感。因此,环境因子对地衣生态位可能具有重要的影响。
达坂城山区为严重的干旱半干旱区,相较于其他研究区,光照较强,裸露岩石居多。地衣被认为是荒漠拓荒者,在植物群落演替早期阶段具有重要的作用。在该地区植被中地衣占据绝对优势,尤其在高等植物无法生长的岩石表面上可见大量地衣覆盖,因此,地衣在该地区生态系统中具有重要作用。本研究通过计算达坂城山区地衣生态位宽度和生态位重叠值,结合环境因子,定量分析地衣的生态位特征,以期揭示以下科学问题:(1)地衣物种的生态位有何特征;(2)环境因子对物种生态位特征有何影响。在传统的地衣物种多样性、物种分布格局的基础上,从生态位角度探讨地衣物种多样性的维持机制,从物种与环境、物种之间和物种自身特性方面提供更全面的信息,以期为该地区地衣物种的多样性保护提供依据。
1 材料与方法
1.1 研究区概况
达坂城区位于西天山和东天山最高峰博格达峰南麓之间,在乌鲁木齐市的南郊,距乌鲁木齐市区86 km,为三面环山的半封闭谷地。达坂城区北部山区为东天山最高峰博格达峰南坡,北部山区约占辖区总面积的三分之一,位于88°19′—88°32′ E、44°21′—44°38′ N。该地区地形地貌比较复杂,山势呈西南向东北升高,随着海拔高度的变化,其气候变化差异显著。由于山体构造差异,导致达坂城山区海拔1 800 m以下遭受严重的荒漠影响,属中温带大陆性干旱气候,四季干旱少雨,属于严重干旱、半干旱地区,为欧亚大陆腹地典型的荒漠、半荒漠区,也是乌鲁木齐的重要生态屏障和生态恢复带,由于其特殊的气候条件,使该地区生物呈现出特殊的荒漠气候适应机制(Thomas et al., 2003;任泉等,2011)。
1.2 研究方法
1.2.1 野外调查和采样 在达坂城区6个地点进行。在海拔1 500~3 500 m之间,选择30个面积为20 m × 20 m的样地。在样地内随机设置5个1 m × 1 m的小样方,筛查样方内所有地衣种类,并记录基物类型。用50 cm × 50 cm的铁筛计测每种地衣所占的网格数,根据网格所占比例得出样地中每种地衣的平均盖度。依据地衣和植被分布带特点,将采样区域分为4个海拔分布区域,即低海拔(<2 000 m,9个样方)、中低海拔(2 000~2 500 m,9个样方)、中高海拔(2 500~3 000 m,3个样方)、高海拔(>3 000m,9个样方)。将采集的地衣带回实验室制成标本,在实验室用形态学和解剖学,并结合化学显色反应等鉴定地衣种类。标本鉴定参考新疆地衣(阿不都拉·阿巴斯和吳继农,1998),中国地衣综览第二版和北美地衣,以及最新发表文献等鉴定物种。所采集的标本保存在新疆大学地衣标本馆(XJU-L)。
1.2.2 Levins生态位宽度和Shannon-Wiener生态位宽度
根据张金屯(2004)的方法计算其宽度值,计算公式如下。
式中: Bα为种i的Levins生态位宽度;Bi为种i的Shannon-Wiener生态位宽度;Pij =nij /Ni+,代表种i在第j个资源状态下的个体数占该种所有个体数的比例,nij 为种群i利用资源状态j的数量(以种i在第j个样方中的物种盖度表示),Ni 为种群i的总数量;r为样方数。
1.2.3 Pianka生态位重叠 生态位重叠值的计算参考张金屯(2004)的方法,计算公式如下。
式中: Oik为种i和种k间的生态位重叠值,具有值域 [0,1];Pkj =nkj /Nk+,代表种k在第j个资源状态下的个体数占该种所有个体数的比例,nkj为种群k利用资源状态j的数量,Nk为种群k的总数量;r为样方数。样地全部种群间生态位重叠值的总平均值=样地内全部种群间生态位重叠值总数/总种对数。
1.2.4 物种丰富度(S)、Shannon-Wiener多样性指数(H)、Simpson指数(D)和Pielou均匀度指数(J)
4个参数值的计算参考张金屯(2004)的方法,计算公式如下。
式中:S为样方中的物种数,即物种丰富度;Pi表示第i个种的多度比例,即Pi=Ni/N0,Ni为种群i的总数量,N0为S个物种的数量之和。
1.3 数据处理
生态位宽度值和生态位重叠值,物种多样性指数的计算及聚类分析均在R 4.0.2 软件中的“spaa”和“cluster”包完成。生态位特征与环境因子的CCA排序、RDA排序和PCoA排序分析均在 CANOCO 5.0软件中完成。
2 结果与分析
2.1 达坂城山区地衣的生态位宽度
达坂城山区地衣物种的Levins生态位宽度(Bα)和Shannon-Wiener生态位宽度(Bi)的趋势一致(表1)。Levins生态位最宽的物种为内卷野粮衣(Circinaria contorta),Levins和Shannon-Wiener生态位宽度值分别为Bα=12.093 4和Bi=2.600 7;Shannon-Wiener生态位宽度最宽的物种为丽黄鳞衣(Rusavskia elegans),Levins和Shannon-Wiener生态位宽度值分别为Bα=11.665 5和Bi=2.692 9。这两个种的生态位最宽,在所调查的大多数样方中分布,分布范围广,对环境资源的利用充分。生态位次宽的物种为碎茶渍(Lecanora argopholis)、方斑网衣(Lecidea tessellata)、怀俄明黄梅(Xanthoparmelia wyomingica)、莲座微孢衣(Acarospora rosulata)、粉瓣茶衣(Lobothallia alphoplaca)、盾原类梅(Protoparmeliopsis peltata)等,适应范围较广,能够较充分地利用生境资源。
生态位较窄的物种有泡状微孢衣(Acarospora bullata)、斯塔妃微孢衣(A. stapfiana)、朝鲜黄梅(Xanthoparmelia coreana)、软鹿石蕊(Cladonia mitis)、矮石蕊(C. humilis)等38个物种,其Levins和Shannon-Wiener生态位宽度值分别为Bα=1.000 0和Bi=0.000 0,他们在调查中仅出现在1个样方中,其生态适应范围窄。
统计不同生态位宽度的物种数,Levins生态位宽度Bα<2.000 0的物种共有64种,Shannon-Wiener生態位宽度Bi<1.000 0的物种有69种(表2),由此可知,达坂城山区大多数地衣生态位宽度较小,生态适应范围窄,资源利用能力较弱,他们分布在不同环境区域的个别样方中,分布相对集中,导致其生态位宽度指数较低。
2.2 达坂城山区地衣的生态位重叠
达坂城山区地衣物种间的重叠值偏低,生态位重叠值小于0.100的种对占所有种对的80.07%,其中生态位重叠值为0.000的种对占所有种对的66.81%,生态位重叠值在0.100~0.500之间的种对占所有种对的15.51%,生态位重叠值在0.500~1.000之间的种对仅占4.42%。这说明不同地衣种群间资源利用的相似程度较低,各物种对生境资源的适应存在差异,物种在生境资源的利用上形成了生态位的分化,种间竞争不激烈。
一般认为,生态位较宽的物种分布越广,与其他物种之间生态位重叠的机会就大,越容易出现较大的生态位重叠。Levins生态位最宽的内卷野粮衣与77个种之间存在生态位重叠,但重叠值小于0.650,Shannon-Wiener生态位最宽的丽黄鳞衣与94个物种之间都存在生态位重叠,与黑小极衣(Lichinella nigritella)的生态位重叠值为0.840,但与其他物种之间的重叠值较低。生态位较窄的物种仅与较少物种之间存在生态位重叠,但生态位重叠值都较高。这说明生态位宽的物种与其他物种间出现重叠的概率较大,但重叠值不一定较高,而生态位宽度较窄的物种与其他物种出现重叠的概率较小,但重叠值可能较高。
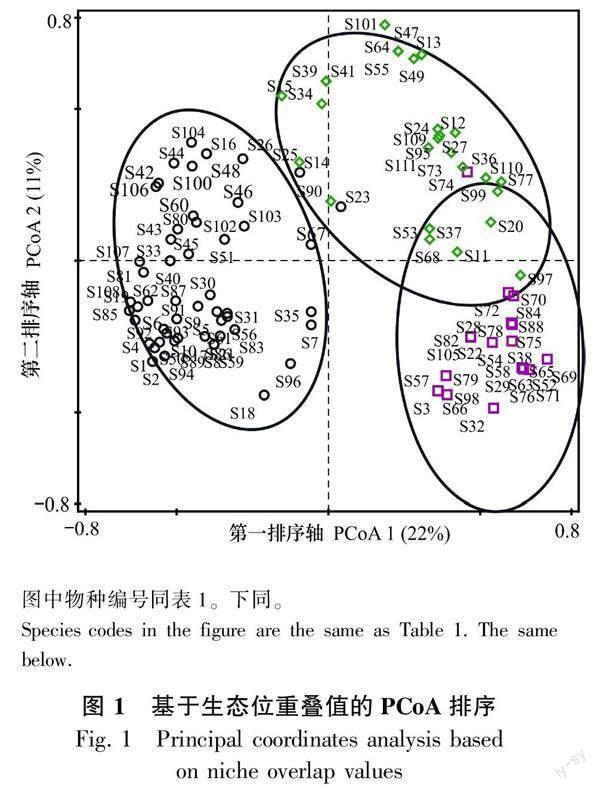
以生态位重叠值为指标,对达坂城山区调查的111种地衣物种进行聚类分析和排序分析,将聚类结果与PCoA排序结果相结合,可将达坂城山区的111种地衣物种划分为3个生态类群(图1)。
第一个生态类群,包括丽黄鳞衣、内卷野粮衣、碎茶渍、莲座黄茶渍、垫微孢衣等55种地衣,这些地衣在所调查的不同海拔区域的样方中均有分布,分布范围较广且Levins生态位最宽的内卷野粮衣和Shannon-Wiener生态位最宽的丽黄鳞衣也属于该生态类群,与大部分地衣物种之间都存在生态位重叠。第二个生态类群,包括鳞饼衣、亚兰灰平茶渍、桃红平茶渍、疣平茶渍等27种地衣,这些地衣大部分主要分布在3 000 m以上的高海拔区域,无人为干扰,风速较强、湿度较高、光照较弱的向阴区域。第三个生态类群,包括方斑网衣、盾原类梅、淡肤根石耳、金黄茶渍、亚洲多孢衣等29种地衣,这些地衣主要分布在2 500~3 000 m的中高海拔区域,具有较弱的人为干扰且湿度较高的区域。
由此可知,达坂城山区地衣物种在不同海拔区域之间具有差异,主要是由不同海拔区域生境条件的差异而导致。该地区低海拔区域较为干旱,光照较强,随着海拔的升高,湿度增加,能够提供更多的生态位供物种生存。
2.3 不同海拔梯度地衣物种的分布
在达坂城山区采样海拔区域范围之间,根据本地区植被分布带和样地分布特点,以500 m间隔将采样区域分为4个海拔分布区域,分析不同海拔范围地衣的物种多样性及分布格局。基于达坂城山区30个样方中111个物种的盖度数据,计算样地间的Bray-Curtis距离,利用PCoA主坐标分析并绘制排序图(图2)。由图2可知,第一轴和第二轴解释了总方差的34%,不同海拔样地沿第一轴和第二轴有序排列,低海拔样地之间离散程度较大,高海拔区域分布相对聚集,说明低海拔区域各样地地衣群落物种差异性较大。但是,该地区低海拔区域至高海拔区域样地沿第一轴有序排列,高低海拔梯度间样地地衣群落物种之间具有差异。
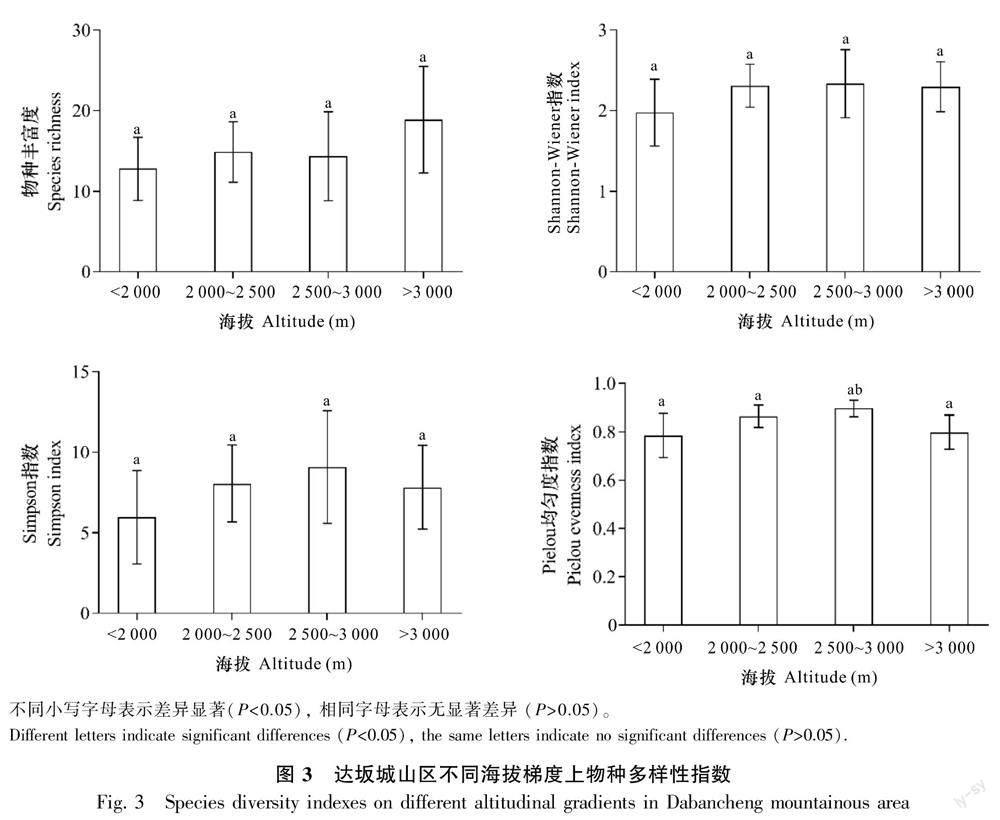
计算不同海拔区域地衣物种的多样性指数, 并进行单因素方差分析, 结果见图3。由图3可知,物种丰富度在3 000 m以上的高海拔区域最高,低海拔区域(<2 000 m)最低。Shannon-Wiener多样性指数、Simpson指数和Pielou均匀度指数在中高海拔区域(2 500~3 000 m)较高,在低海拔(<2 000 m)和高海拔区域(<3 000 m)较低。单因素方差分析表明,Pielou均匀度指数在3 000 m以上区域和2 500 m以下海拔区域之间差异显著(P<0.05),其他物种多样性指数在不同海拔区域之间无显著差异(P>0.05)。
2.4 生态位特征的环境解释
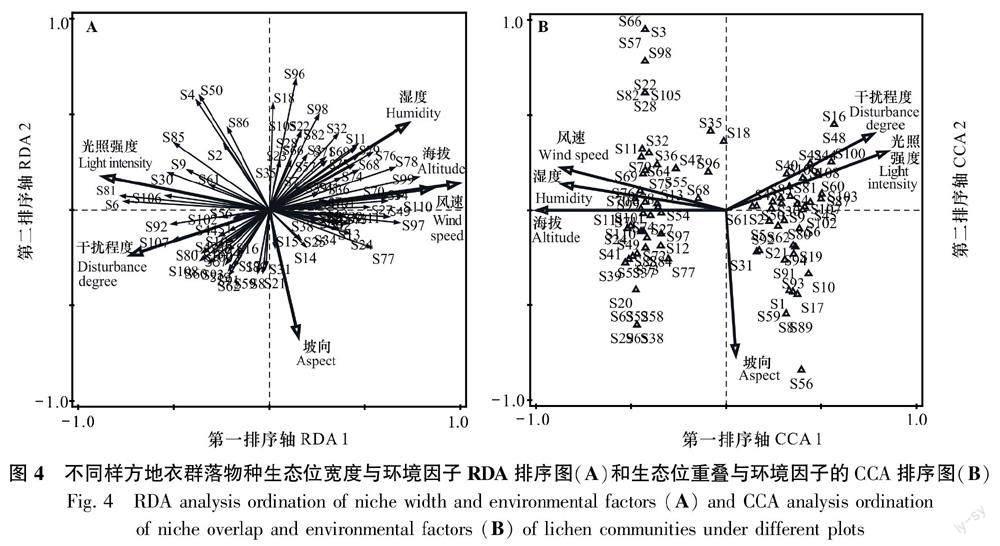
分别对不同样方地衣群落物种的生态位宽度和生态位重叠值与环境因子进行DCA分析,结果显示生态位宽度第一排序轴的梯度长度介于3~4之间, 并且RDA前四排序轴累计方差解释率均比CCA排序大,因此生态位宽度和环境因子的关系用RDA分析,而生态位重叠第一排序轴的梯度长度大于4,因此选择CCA排序,并用二维投影图显示(图4)。
由生态位宽度的RDA排序图(图4:A)可知,海拔、风速和湿度与生态位宽度呈正相关,光照强度、干扰和坡向与生态位宽度呈负相关。由生态位重叠的CCA排序图(图4:B)可知,光照强度和干扰与生态位重叠呈正相关,海拔、湿度、风速和坡向与生态位重叠值呈负相关。总体来看,与地衣群落物种生态位宽度和生态位重叠相关性最强的环境因子为海拔,其次为光照强度、风速、湿度、干扰程度、坡向,其中,坡向对生态特征的影响不显著。
3 讨论与结论
生态位宽度量化反映了物种对生境资源利用的多样化水平或特化水平,以及对环境的适应状况差异(聂莹莹等,2021)。也就是说,生态位宽度越宽,对环境资源的利用能力越强,往往会形成群落中的优势物种(孙成等,2022)。本研究发现,丽黄鳞衣广泛分布于不同海拔和生境中的岩石表面,对不同的环境具有较强的适应性,为该地区的优势物种,这与丽黄磷衣自身对环境的高耐受性有关。丽黄鳞衣对污染元素等具有较高的富集能力(赵丽丽等,2019),对恶劣生境具有耐受性,致使其泛化程度较高,对生境资源利用充分。此外,内卷野粮衣生态位宽度也较大,这可能是在生态位研究中将群落调查中的每个取样样地作为资源状态,从而反映物种对资源的综合利用狀况(罗文等,2021),内卷野粮衣分布在大部分样地中,虽然其多度较低,但同样能利用不同的生境资源,说明这两个物种对生存环境要求较低,能够适应该地区较恶劣的生境,对该地区地衣群落构建起着重要的作用。该地区大部分地衣生态位宽度较窄,这与该地区生境条件有密切联系,低海拔区域荒漠化严重,物种多为耐干旱的岩面生地衣,而在高海拔区域,具有显著较高比例的叶状地衣等,不同生境中生态需求相似物种的聚集分布,致使其特化程度较高,对环境资源利用程度低。
生态位重叠侧重反映了物种在空间中资源利用的交叉重叠程度,以及物种间的生物和生态学特性的相似度,可在一定程度上解释物种间的共生和竞争作用(肖艳梅等,2021)。生态位重叠值越高,种间资源需求越相似,种间竞争越激烈。本地区地衣种对间生态位重叠值整体偏低,说明地衣物种的生态位普遍存在差异,生态位分化程度高,物种间竞争不激烈,群落较稳定。虽然研究区整体干旱,生境异质性较低,但由于地衣生物体极小,小生境对其极为重要,地衣会占据自身独特的小生境,使种间关系处于协调和稳定状态。Gause(1934)认为,生态位重叠仅仅是物种竞争的一个不必要的前提条件,即物种间是否存在竞争要取决于具体生境,只有在资源缺乏或不足的生境条件下,生态位重叠才会产生竞争排斥。该地区整体环境较为恶劣,资源相对匮乏,可以认为地衣物种在定殖早期由于资源争夺竞争激烈,随后,物种逐渐占据各自适宜的生态资源位,形成了生态位分化,导致其重叠值整体偏低。另外,由于将调查样地视为生境资源位的综合,物种分布的交错程度较大也会导致其较低的生态位重叠值(宗宁等,2021)。
物种生态位宽度值越大,分布范围越广,越容易与其他物种发生生态位重叠(景升利等,2020)。该地区地衣物种大都为岩面生壳状地衣,对环境适应能力强,分布在不同海拔区域,与其他物种之间出现生态位重叠的概率较大。如生态位较宽的内卷野粮衣和丽黄鳞衣与多数物种之间都存在生态位重叠,但重叠值较低;而一些生态位较窄的物种,如软鹿石蕊、矮石蕊、小美衣、蜡黄橙衣、斑点野粮衣等,由于其特殊的生境需求,主要分布在无人为干扰, 较高湿度的高海拔区域,本研究中只在单个样方中有记录,却与其他物种之间存在较高的生态位重叠。这与Sun等(2021)的一些研究结果一致,认为生态位重叠和生态位宽度之间并没有直接的线性关系,这可能与物种的生物学和生态学特性有关(郭永清等,2009),导致生境资源利用的互补性,或是因为其只在同一样方中存在。
生态位特征的差异是群落各环境因子共同作用的结果,已有研究表明,地衣物种分布与光照、湿度、温度、海拔(Baniya et al., 2010;Sahu et al., 2019;Nascimento et al., 2021)等环境因素有关。达坂城山区荒漠化严重,生境干旱,资源较为匮乏。通过排序分析可以看出海拔是地衣生态位特征的重要影响因素。随着海拔的升高,湿度增加,人为干扰减弱,更适宜物种定殖。同时,水分增加更利于叶状和枝状等水分需求较高的地衣生长(Gauslaa, 2014),在2 500~3 000 m中高海拔区域,物种多样性最高。在高海拔区域,地衣物种多样性降低,主要是由于高海拔地区生境条件严酷,风速较强,可能不利于地衣生长(Pottier et al., 2013)。本研究发现,海拔、湿度、光照强度对地衣物种生态位特征的影响较大,主要是由于海拔影响湿度和光照条件,因此,沿海拔梯度的生境条件的差异导致物种生态位差异。总体而言,达坂城山区地衣物种沿海拔梯度分布具有差异,物种间由于生境资源竞争形成了生态位的分化,群落较为稳定。但是,由于相对恶劣的生境条件,群落较为脆弱,物种对外部干扰较为敏感,干扰可能会影响地衣物种多样性,从而影响该地区生态系统稳定。因此,避免对该地区进行较大的人为干扰,对该地区的荒漠生境研究和生態环境的保护具有重要意义。
参考文献:
ABBAS A, WU JN, 1998. Lichens of Xinjiang [M]. Urumqi: Xinjiang Science-Technology and Health Publishing House: 1-161.[阿不都拉·阿巴斯, 吴继农, 1998. 新疆地衣 [M]. 乌鲁木齐: 新疆科技卫生出版社: 1-161.]
BANIYA CB, SOLHY T, GAUSLAA Y, et al., 2010. The elevation gradient of lichen species richness in Nepal [J]. Lichenologist, 42(1): 83-96.
CAO H, ZHANG CY, GONG XF, et al., 2021. Niche characteristics and interspecific associations of dominant tree species in secondary evergreen broad leaf forest of Suichang County [J]. J Zhejiang Sci Technol, 41(5): 15-21.[曹华, 张川英, 龚笑飞, 等, 2021. 遂昌县次生常绿阔叶林主要物种的生态位特征及种间联结 [J]. 浙江林业科技, 41(5): 15-21.]
CHEN X, BU ZJ, WANG SZ, et al., 2009. Niches of seven bryophyte species in Hani peat land of Changbai Mountains [J]. Chin J Appl Ecol, 20(3): 574-578.[陈旭, 卜兆君, 王升忠, 等, 2009. 长白山哈泥泥炭地七种苔藓植物生态位 [J]. 应用生态学报, 20(3): 574-578.]
GAUSE GF, 1934. The struggle for existence [M]. New York: Hafner: 12-26.
GAUSLAA Y, 2014. Rain, dew, and humid air as drivers of morphology, function and spatial distribution in epiphytic lichens [J]. Lichenologist, 46(1): 1-16.
GRINNELL J, 1917. The niche-relationships of the California Thrasher [J]. Auk, 34(4): 427-433.
GUO YQ, LANG NJ, JIANG QC, et al., 2009. Niche characteristics of the plants in the dry and hot valley of Yuanmou County [J]. J NW For Univ, 24(2): 13-17.[郭永清, 郎南军, 江期川, 等, 2009. 元谋干热河谷植物生态位特征研究 [J]. 西北林学院学报, 24(2): 13-17.]
GIORDANI P, BENESPERI R, MARIOTTI MG, 2015. Local dispersal dynamics determine the occupied niche of the red-listed lichen Seirophora villosa (Ach.) Frdén in a mediterranean Juniperus shrubland [J]. Fungal Ecol, 13: 77-82.
IMIN B, LI ZS, TUMUR A, 2022. Niche characteristics of saxicolous lichens in Tianshan No. 1 Glacier, Xinjiang, China [J]. J Arid Land Resour Environ, 36(1): 159-166.[拜合提妮萨·依明, 李作森, 艾尼瓦尔·吐米尔, 2022. 天山一号冰川岩面生地衣群落主要种群生态位特征的研究 [J]. 干旱区资源与环境, 36(1): 159-166.]
JIN WT, TUMUR A, 2017. Niche of saxicolous lichens communities in Tomur Peak National Nature Reserve in Xinjiang, China [J]. Ecol Sci, 36(1): 89-94.[靳文婷, 艾尼瓦尔·吐米尔, 2017. 托木尔峰国家级自然保护区岩生地衣群落生态位研究 [J]. 生态科学, 36(1): 89-94.]
JING SL, YANG HL, WANG JT, 2020. Niche characteristics of plant communities in shallow mountain plantations inloess alpine region [J]. Open J Soil Water Conserv, 8(4): 23-37.[景升利, 杨海龙, 王佳庭, 2020. 黄土高寒区浅山人工林植物群落生态位特征研究 [J]. 水土保持, 8(4): 23-37.]
KEDDY PA, 1992. Assembly and response rules: two goals for predictive community ecology [J]. J Veg Sci, 3(2): 157-164.
KOCH NM, MATOS P, BRANQUINHO C, et al., 2019. Selecting lichen functional traits as ecological indicators of the effects of urban environment [J]. Sci Total Environ, 654(1): 705-713.
LI ZS, LUO P, TUMUR A, 2021. Niche of saxicolous lichens in Haxionggou Forest Park in Xinjiang, China [J]. J NW For Univ, 36(4): 257-265.[李作森, 雒鹏, 艾尼瓦尔·吐米尔, 2021. 新疆哈熊沟森林公园岩面生地衣生态位的研究 [J]. 西北林学院学报, 36(4): 257-265.]
LUBEK A, KUKWA M, JAROSZEWICZ B, et al., 2018. Changes in the epiphytic lichen biota of Biaowiea Primeval Forest are not explained by climate warming [J]. Sci Total Environ, 643: 468-478.
LUO W, ZHONG YF, XUE SL, et al., 2021. The study on niche of Cinnamomum rigidissimum and dominant populations in different communities in Jianfengling [J]. For Resour Manage, 5: 160-167.[羅文, 钟育飞, 薛少亮, 等, 2021. 尖峰岭不同群落的卵叶桂与优势种群的生态位研究 [J]. 林业资源管理, 5: 160-167.]
LUCHETA F, KOCH NM, KFFER MI, et al., 2019. Lichens as indicators of environmental quality in southern Brazil: an integrative approach based on community composition and functional parameters [J]. Ecol Indicators, 107: 105587.
NASCIMENTO E, MAIA LC, CCERES M, et al., 2021. Phylogenetic structure of lichen meta communities in Amazonian and Northeast Brazil [J]. Ecol Res, 36(3): 440-463.
NIE YY, CHEN JQ, XIN XP, et al., 2021. Responses of niche characteristics and species diversity of main plant populations to duration of enclosure in the Hulun Buir meadow steppe [J]. Acta Pratacul Sin, 30(10): 15-25.[聂莹莹, 陈金强, 辛晓平, 等, 2021. 呼伦贝尔草甸草原区主要植物种群生态位特征与物种多样性对封育年限响应 [J]. 草业学报, 30(10): 15-25.]
NIU KC, CHU CJ, WANG ZH, 2022. Dynamic niche: a new foundation for rebuilding theory of community ecology [J]. Sci Sin (Vit), 52(3): 403-417.[牛克昌, 储诚进, 王志恒, 2022. 动态生态位: 构建群落生态学理论的新框架 [J]. 中国科学: 生命科学, 52(3): 403-417.]
PANG JS, YUAN YC, ZHOU MM, et al., 2022. Species niche of Clematis acerifolia Maxim. community [J]. Acta Ecol Sin, 42(8): 3449-3457.[庞久帅, 原阳晨, 周苗苗, 等, 2022. 槭叶铁线莲(Clematis acerifolia Maxim.)群落中物种生态位研究 [J]. 生态学报, 42(8): 3449-3457.]
REN Q, CAI XT, MA WH, 2011. Changing features of temperature and precipitation in recent 52 years in Dabancheng region [J]. J Arid Land Resour Environ, 25(10): 116-121.[任泉, 蔡新婷, 馬文惠, 2011. 新疆达坂城地区52a来气温和降水变化特征分析 [J]. 干旱区资源与环境, 25(10): 116-121.]
POTTIER J, DUBUIS A, PELLISSIER LC, et al., 2013. The accuracy of plant assemblage prediction from species distribution models varies along environmental gradients [J]. Glob Ecol Biogeogr, 22(1): 52-63.
ROLSHAUSEN G, DAL GF, SADOWSKA-DES' AD, et al., 2018. Quantifying the climatic niche of symbiont partners in a lichen symbiosis indicates mutualist-mediated niche expansions [J]. Ecography, 41(8): 1380-1392.
SAHU N, SINGH SN, SINGH P, et al., 2019. Microclimatic variations and their effects on photosynthetic efficiencies and lichen species distribution along elevational gradients in Garhwal Himalayas [J]. Biodivers Conserv, 28: 1953-1976.
SALES LP, HAYWARD MW, LOYOLA R, 2021. What do you mean by “niche”? modern ecological theories are not coherent on rhetoric about the niche concept [J]. Acta Oecol, 110: 103701.
SUN C, QIN FC, LI L, et al., 2022. Environmental interpretation of ecological niche characteristic of herbaceous populations under different site types of Pinus tabuliformis forest in feldspathic sandstone region [J]. Acta Ecol Sin, 42(9): 3613-3623.[孙成, 秦富仓, 李龙, 等, 2022. 砒砂岩区不同立地类型人工油松林下草本种群生态位特征及其环境解释 [J]. 生态学报, 42(9): 3613-3623.]
SUN JC, LI YH, DENG DT, et al.,2021. Niche analysis of dominant species in alpine desert grassland communities in qaidam basin [J]. E3S Web Conf, 257(3): 03021.
THOMAS CD, CAMERON A, GREEN RE, et al., 2003. Extinction risk from climate change [J]. Nature, 427: 145-148.
TIAN YN, TUMUR A, 2021. Niche characteristics of saxicolous crustose lichens in the northern slope of Bogda peak, Xinjiang [J]. J Arid Land Resour Environ, 35(1):108-113.[田雅楠, 艾尼瓦尔·吐米尔, 2021. 新疆博格达峰北坡岩面生壳状地衣的生态位特征 [J]. 干旱区资源与环境, 35(1): 108-113.]
TUMUR A, SATTAR M, MUHAMAT M, et al., 2012. Niche of ground lichens in southern mountainous region of Urumqi, Xinjiang, China [J]. J Arid Land Resour Environ, 26(7): 116-120.[艾尼瓦尔·吐米尔, 买买提·沙塔尔, 马衣拉·莫合买德, 等, 2012. 新疆乌鲁木齐南部山区地面生地衣生态位研究 [J]. 干旱区资源与环境, 26(7): 116-120.]
VANNETTE RL, FUKAMI T, 2014. Historical contingency in species interactions: towards niche-based predictions [J]. Ecol Lett, 17(1): 115-124.
XIAO YM, XIE JY, YAO YP, et al., 2021. Niche dominant species in arbor layer of evergreen deciduous broad-leaved mixed forest in karst hills of Guilin, southwestern China [J]. Acta Ecol Sin, 41(20): 8159-8170.[肖艳梅, 解婧媛, 姚义鹏, 等, 2021. 桂林岩溶石山常绿落叶阔叶混交林乔木层优势物种生态位研究 [J]. 生态学报, 41(20): 8159-8170.]
ZHANG JT, 2004. Quantitative ecology [M]. Beijing: Science Press: 94-130.[张金屯, 2004. 数量生态学 [M].北京: 科学出版社: 94-130.]
ZHAO LL, YANG MJ, LUO ZD, et al., 2019. Lichen (Xanthoria elegans) biomonitoring of harmful element pollution in Taiyue Mountains, Shanxi Province, China [J]. Life Sci Res, 23(1): 28-34.[赵丽丽, 杨梦杰, 罗治定, 等, 2019. 山西省太岳山区有害元素大气污染的地衣( 丽石黄衣) 监测 [J]. 生命科学研究, 23(1): 28-34.]
ZONG N, SHI PL, ZHU JT, 2021. Changes of plant community composition and niche characteristics during desertification process in an alpine steppe [J]. Ecol Environ Sci, 30(8): 1561-1570.[宗宁, 石培礼, 朱军涛, 2021. 高寒草地沙化过程植物群落构成及生态位特征变化 [J]. 生态环境学报, 30(8): 1561-1570.]
(责任编辑 周翠鸣 李 莉)
猜你喜欢
散文诗(2021年22期)2022-01-12 06:14:04
落叶果树(2021年6期)2021-02-12 01:29:10
环球时报(2019-05-23)2019-05-23 06:17:36
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
北京文学(2017年12期)2017-12-12 20:23:37
体育科技(2016年2期)2016-02-28 17:06:07
吐鲁番(2015年1期)2015-11-17 19:53:54
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
河北遥感(2014年4期)2014-07-10 13:54:58
新晨(2013年1期)2014-04-03 03:40:48
