
1,8-cineole ameliorates colon injury by downregulating macrophage M1 polarization via inhibiting the HSP90-NLRP3-SGT1 complex
2023-10-24 02:40:22ShengsuoBingYngYngDuYiwenLvJirongLiuYucongShiTingHungHuchongXuLiDengXioyinChen
Shengsuo M ,Bing Yng ,Yng Du ,Yiwen Lv ,Jirong Liu ,Yucong Shi ,Ting Hung ,Huchong Xu ,*,Li Deng ,**,Xioyin Chen ,***
a School of Traditional Chinese Medicine,Jinan University,Guangzhou,510632,China
b Guangzhou Key Laboratory of Formula-Pattern of Traditional Chinese Medicine,Jinan University,Guangzhou,510632,China
Keywords:1,8-cineole Amomum compactum Sol.ex Maton NLRP3 HSP90 Ulcerative colitis Intestinal barrier function
ABSTRACT Ulcerative colitis(UC)is characterized by chronic relapsing intestinal inflammation.Currently,there is no effective treatment for the disease.According to our preliminary data,1,8-cineole,which is the main active compound of Amomum compactum Sol.ex Maton volatile oil and an effective drug for the treatment of pneumonia,showed remarkable anti-inflammatory effects on colitis pathogenesis.However,its mechanism of action and direct targets remain unclear.This study investigated the direct targets and mechanism through which 1,8-cineole exerts its anti-inflammatory effects using a dextran sulfate sodium salt-induced colitis mouse model.The effects of 1,8-cineole on macrophage polarization were investigated using activated bone marrow-derived macrophages and RAW264.7 cells.In addition,1,8-cineole targets were revealed by drug affinity responsive target stability,thermal shift assay,cellular thermal shift assay,and heat shock protein 90 (HSP90) adenosine triphosphatases (ATPase) activity assays.The results showed that 1,8-cineole exhibited powerful anti-inflammatory properties in vitro and in vivo by inhibiting the macrophage M1 polarization and protecting intestinal barrier function.Mechanistically,1,8-cineole directly interacted with HSP90 and decreased its ATPase activity,also inhibited nucleotide-binding and oligomerization domain-,leucine rich repeat-,and pyrin domain-containing 3(NLRP3) binding to HSP90 and suppressor of G-two allele of SKP1 (SGT1) and suppressed NLRP3 inflammasome activation in macrophages.These results demonstrated that 1,8-cineole is a potential drug candidate for UC treatment.
1.Introduction
Ulcerative colitis (UC) is a chronic immune inflammatory disease in the gastrointestinal tract that is characterized as a modern refractory disease by the World Health Organization [1].In recent decades,the global incidence and prevalence of UC have been increasing over time worldwide,especially in the Northern Europe,Canada,and Australia.Additionally,the incidence of UC has also been steadily increasing in Asia,the Middle East,and South America[2].Although it has gradually increased in recent years,the etiology of UC remains unknown.Most investigators concur that UC is caused by patient's genetics,microbiome,immune response,and the environment [3].With the development of genome-wide association and immunological studies,numerous studies have shown that aberrant immune responses may play a central role in the pathogenesis of UC [4].The abnormally activated innate and adaptive immune cells in the colon release proinflammatory mediators and antibodies,which drive the inflammatory response and lead to impaired intestinal barrier function.However,immunesuppressor or anti-inflammatory treatment often results in treatment failure or unacceptable side effects.Thus,the development of novel anti-inflammatory drugs or immunomodulators remains the main direction of UC therapy.
1,8-cineole is the main active compound ofAmomum compactumSol.ex Maton volatile oil (ACO) and is well known for its extensive pharmacological activities,including anti-inflammatory,antioxidant,antiviral and antimicrobial effects [5-7].In addition,1,8-cineole,a main compound of Eucalyptus limonene-pinene soft capsules,has also been approved by the National Medical Products Administration,China for the clinical treatment of pneumonia.Our preliminary results showed that ACO could improve colonic barrier function by upregulating the expression of the tight junction proteins zonula occludens-1 (ZO-1) and occludin in the intestines of pneumonia model mice (unpublished observations).We further found that 1,8-cineole could also reduce intestinal inflammation.Despite this,the specific mechanism remains unclear.Most previous studies have focused on its effects of 1,8-cineole on the regulation of transcription factor activity and intracellular inflammatory factor production [7,8].However,there is no evidence that reveals the direct binding target of 1,8-cineole.In this experiment,we demonstrated that 1,8-cineole has a suitable therapeutic effect on UC pathogenesis via inhibiting the heat shock protein 90(HSP90)-nucleotide-binding and oligomerization domain-,leucine rich repeat-,and pyrin domain-containing 3 (NLRP3)-suppressor of Gtwo allele of SKP1 (SGT1) complex.It strongly inhibits the development of intestinal inflammation by reducing M1 polarization of macrophages,which in turn inhibits the release of proinflammatory cytokine.Thus,1,8-cineole represents a therapeutic candidate for the treatment of UC.
2.Materials and methods
2.1.Materials and reagents
DriedAmomum compactumSol.ex Maton fruits were purchased from Kangmei Pharmaceutical (Puning,China).1,8-cineole(PHR1828),pronase (PRON-RO),and lipopolysaccharide (LPS;L2630) were purchased from Merck (Kenilworth,NJ,USA).Interferon γ(IFNγ;507209-MNAH)were purchased from Sino Biological(Beijing,China).Dextran sulfate sodium salt (DSS) (216 011080;molecular weight: 36000-50000 Da) was purchased from MP Biomedicals (San Diego,CA,USA).The details are present in the Supplementary data.
2.2.Extraction of ACO
Dried powder ofAmomum compactumSol.ex Maton fruits were directly subjected to hydrodistillation in a Clevenger-type apparatus for 4 h.The obtained oil was mixed with anhydrous sodium sulfate and stored in -80°C.The chemical components of ACO were analysis by gas chromatography-mass spectrometry (Agilent Technologies,Inc.,Santa Clara,CA,USA).The results are recorded in Table S1 and Fig.S1.
2.3.Animal care and use
Six-week-old male C57BL/6 J mice were obtained from Liaoning Changsheng Biotechnology Co.,Ltd.(Benxi,China).The animals were housed in a room maintained at 25°C,50% relative humidity and 12-h light/12-h dark cycle with free access to food and water.All animal experimental protocols were approved by the Animal Care Committee of Jinan University (Approval No.: IACUC-20201028-02),and all animal experiments were performed in accordance with the Guide for Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication No.85-23,revised in 1996).The details are present in the Supplementary data.
2.4.Induction of colitis and treatments
After 10 days adaptive feeding,all animals were randomly divided into four groups,namely,control (CON),2.5% DSS (DSS),2.5% DSS+1,8-cineole(Cineole),and 2.5% DSS+ACO(ACO).Except for the control mice,colitis was induced in the mice by adding 2.5%(V/V) DSS to the drinking water for 7 days.ACO and 1,8-cineole were dissolved in 1% (V/V) Tween 80 and administered daily by oral gavage.The detailed experimental procedure is shown in Fig.S2.
2.5.Cell culture and treatment
Bone marrow-derived macrophages (BMDMs) were isolated from 6-to 8-weeks-old C57BL/6 mice and cultured in Iscove's modified Dulbecco's medium containing 10% (V/V) fetal bovine serum (FBS),1% (V/V) penicillin/streptomycin,and 30% (V/V) L929 medium supernatant.RAW264.7 macrophages,Caco2 cells,and THP-1 cells were obtained from iCell Bioscience Inc.(Shanghai,China).RAW264.7 cells were cultured in Dulbecco's Modified Eagle Medium.THP-1 cells were cultured in Roswell Park Memorial Institute 1640 medium.All culture media contained 10% (V/V) FBS and 1% (V/V) penicillin/streptomycin.Caco2 cells were cultured in minimum essential medium containing 20%(V/V)FBS and 1%(V/V)penicillin/streptomycin.All cells were incubated at 37°C in a humidified atmosphere of 5% CO2.Macrophages and Caco2 cells were pretreated with different concentrations (50,100,and 200 μM) of 1,8-cineole for 12 h and then exposed to 1 μg/mL LPS and 50 μg/mL IFNγ to induce M1 polarization and cell damage.The details are present in the Supplementary data.
2.6.Drug affinity responsive target stability (DARTS)
As previously described [9,10],RAW264.7 cells were lysed by mammalian protein extraction reagent,and protein quantification was performed using a bicinchoninic acid kit.The cell lysates were mixed with 1,8-cineole or dimethyl sulfoxide(DMSO)overnight at 4°C for the binding reaction.After binding,the lysates were mixed with pronase at a ratio of 1:1000 (m/V) or 1:2000 (m/V).The mixture was incubated at 37°C for 30 min,and then 5× loading buffer was added for proteolysis.Then,the samples were analyzed by liquid chromatography-mass spectrometry (LC-MS;Thermo Fisher Scientific Inc.,Waltham,MA,USA) and sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
2.7.Cellular thermal shift assay (CETSA)
RAW264.7 cells were treated with 1,8-cineole or DMSO for 24 h and washed twice with phosphate buffered saline.Then,the cells were lysed using Pierce™immunoprecipitation (IP) lysis buffer supplemented with Pierce™protease and phosphatase inhibitors(Thermo Fisher Scientific Inc.).After centrifugation (12,000g,10 min,4°C),the supernatant was divided into ten equal parts and heated at 37,53,55,57,59,61,63,65,67,and 69°C for 3 min via a polymerase chain reaction(PCR)instrument(Bio-Rad Laboratories,Hercules,CA,USA),followed by cooling for 3 min at 4°C.The soluble supernatant was quantified by the Western blotting assay,and the HSP90 abundance level was analyzed.
2.8.Thermal shift assay (TSA)
Thermal shift reaction solution was added as follows: 2 μL of 8×SYPRO orange was mixed with 5 μL of 5×reaction buffer,12.5 μL of HSP90 protein solution,and 2.5 μL of 1,8-cineole or vehicle(DMSO).Then,thermal scanning (25-99°C at 1°C/min) was performed using a 7500 real-time PCR system (Thermo Fisher Scientific Inc.,Foster city,CA,USA),and carboxy-X-rhodamine dye was selected for the reporter.The melting curve and melting temperature (Tm) value of the protein were analyzed by TSA-CRAFT software.Tm shifts greater than 2°C were considered statistically significant.
2.9.HSP90 adenosine triphosphatases (ATPase) activity assay
As previously described [11],the recombinant human HSP90β protein was incubated with or without the indicated compounds at 37°C for 1 h,and then,the CellTiter-Glo assay was employed to measure the quantitation of adenosine triphosphate according to the manufacturer's instructions.
2.10.Co-IP
For immunoprecipitation,RAW264.7 cells were treated with LPS and IFNγ for 6 h and then lysed in ice-cold Pierce™IP lysis buffer supplemented with Pierce™protease and phosphatase inhibitors.Then,the lysates were incubated with anti-nucleotide-binding and oligomerization domain-,Leucine rich repeat-,and pyrin domaincontaining 3 (NLRP3),anti-HSP90,anti-SGT1 antibodies,or normal rabbit IgG at 4°C.After incubation overnight,5 μL of protein A/protein G magnetic beads were added to each tube and mixed gently at 4°C for 3 h.The beads were collected with a magnetic stand,and 100 μL of 1×loading buffer was added for elution.Then,the sample was loaded on SDS-PAGE for Western blotting.
2.11.Western blot
Protein was extracted from cells and colon tissue using radio immunoprecipitation assay lysis buffer,and the bicinchoninic acid method was used to determine the total protein concentration.Fifty micrograms of protein samples were loaded onto 10% SDSPAGE gels and then transferred to 0.45-μm polyvinylidene fluoride membranes.The membranes were blocked with NcmBlot blocking buffer for 10 min and incubated with the indicated primary antibody overnight at 4°C.Protein bands were detected with an enhanced chemiluminescence system (Bio-Rad ChemiDoc MP imaging system,Bio-Rad Laboratories) and analyzed by Image Lab 3.0 (Bio-Rad Laboratories).The details are present in the Supplementary data.
2.12.Statistical analysis
Statistical analyses were performed and figures were generated using GraphPad Software (San Diego,CA,USA).All data are presented as the mean ± standard deviation.Comparisons among groups was performed using one-way analysis of variance,followed by Dunnett's test where appropriate.
3.Results
3.1.ACO/1,8-cineole alleviated colon barrier damage in colitis mice
To verify the protective effect of ACO/1,8-cineole against UC,we first developed a chronic colitis mouse model induced by DSS and assessed UC severity by continuous body weight loss,severe disease activity index (DAI) score,and colon shortening.As we mentioned earlier,ACO and 1,8-cineole alleviated diarrhea symptoms,body weight loss,and decreased DAI scores and the spleen index (Figs.1A-D).Moreover,we found that neither ACO nor 1,8-cineole showed significant hepatotoxicity,and both of them reversed the increased release of alanine aminotransferase induced by DSS (Fig.S3).In addition,the ACO and 1,8-cineole also reduced DSS-induced death in mice (Fig.S4).Moreover,myeloperoxidase,a representative inflammatory marker of UC,was also downregulated after ACO or 1,8-cineole treatment.Then,we evaluated the barrier function of the colon.As shown in Fig.1E,fluorescein isothiocyanate-dextran 4 kDa level in plasma was higher in DSS group than in the CON group and significantly decreased after ACO and 1,8-cineole treatment.Moreover,both 1,8-cineole and ACO treatment reversed the downregulated expression of tight junction proteins (ZO-1 and occludin),which play important roles in maintaining intestinal barrier function(Fig.1F).In addition,hematoxylin and eosin staining revealed that ACO and 1,8-cineole remarkably alleviated the symptoms of muscularis propria in the DSS group,including the loss of crypts and surface epithelia,goblet cell loss,thickening and edema in the submucosa,mucosal damage,leukocyte infiltration,and inflammatory infiltrates (Fig.1G).Periodic acid-Schiff staining and immunohistochemistry (IHC) of MUC2 also confirmed these results.As shown in Figs.1G and H,after treatment with 1,8-cineole and ACO,the number of goblet cells was restored,and the expression of MUC2 was upregulated to a certain extent.These results suggested that 1,8-cineole and ACO could significantly restore colonic barrier function and prevent intestinal leakage,and we hypothesize that this effect may be a result of the renewal of intestinal epithelial cells and alleviation of intestinal inflammation by 1,8-cineole.Therefore,we analyzed the expression of Ki67,a proliferation-related targets,in the intestine using IHC staining to determine intestinal epithelial proliferation (Fig.1H).However,1,8-cineole and ACO did not significantly upregulate the expression of Ki67 in the colon,leading us to hypothesize that the restoration of intestinal barrier function by 1,8-cinele and ACO may be related to the inhibition of intestinal inflammation.
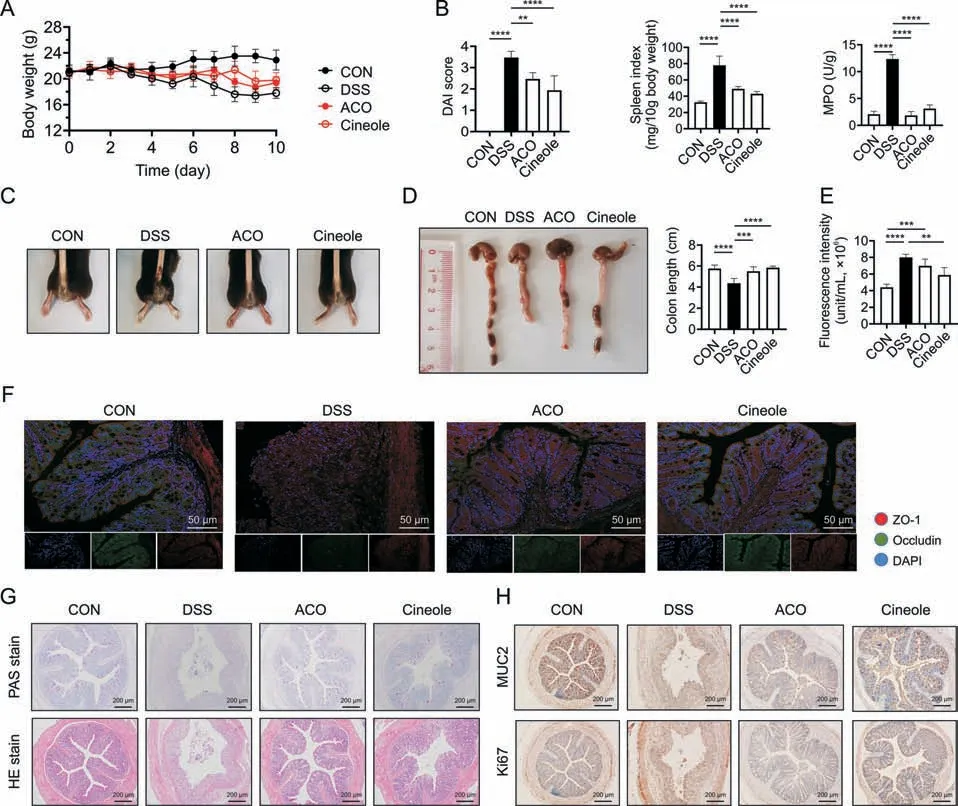
Fig.1. Amomum compactum Sol.ex Maton volatile oil (ACO) and 1,8-cineole (Cineole) inhibited dextran sulfate sodium (DSS)-induced intestinal inflammation and intestinal permeability changes.(A) Body weight.(B) Disease activity index (DAI) score,spleen index,and myeloperoxidase (MPO) level.(C) Representative pictures of the mouse anus.(D)Representative gross anatomical views of the mouse colon and the colon length.(E) Intestinal permeability of fluorescein isothiocyanate-dextran 4 kDa (FD4).(F) Immunofluorescence staining of zonula occludens-1 (ZO-1) (red) and of occludin integral membrane protein (occludin) (green) in mice colons.(G) Periodic acid-Schiff (PAS) staining and hematoxylin and eosin(HE)staining of mice colons.(H)Immunohistochemistry staining of mucin-2(MUC2)and of Ki67 in mice colons.All experiments were performed at least in quadruplicate,and the data represent mean ± standard deviation.ns: no statistical significance.**P <0.01,***P <0.001,and ****P <0.0001.CON: control;DAPI: 4′,6-diamidino-2-phenylindole.
3.2.1,8-cineole reduced DSS-induced increases in lymphocyte and monocyte numbers
Next,flow cytometry and routine blood examination were used to analyze the changes in immune cells after 1,8-cineole treatment.The proportion of CD3+T cells (CD45+and CD3+) in peripheral blood mononuclear cells and mesenteric lymph nodes(MLNs)was significantly decreased after treatment with 1,8-cineole (Fig.2A).However,the analysis of CD4+T cells and CD8+T cells showed that there was no significant change in the proportion of CD4+and CD8+lymphocytes between the DSS model group and the 1,8-cineole-treated group.However,the proportion of monocytes(CD11b+) was significantly increased in the DSS group,and treatment with 1,8-cineole reversed this phenomenon.Routine blood examination also verified this result.The number of total white blood cells,the number and proportion of monocytes,and the number and proportion of granulocytes in the blood were significantly increased after DSS induction,while the proportion and number of these cells were significantly decreased after 1,8-cineole treatment (Fig.2B),which indicated that 1,8-cineole may regulate the proportion of monocytes to inhibit the colitis inflammation.

Fig.2. 1,8-cineole (Cineole) treatment altered the proportion of several immune cell subsets in the peripheral blood mononuclear cells (PBMCs) and mesenteric lymph nodes(MLNs).(A)The percentage of CD3+T lymphocyte(CD45+and CD3+),CD4+T lymphocyte(CD4+,gated on CD45+and CD3+),CD8+T lymphocyte(CD8+,gated on CD45+and CD3+),and monocyte (CD11b+) in PBMCs and MLNs.(B) The number of total white blood cells (WBC),the number and proportion of monocytes,and the number and proportion of granulocytes in the blood were determined using routine blood tests.All experiments were performed at least in quadruplicate,and the data represent mean±standard deviation.ns:no statistical significance.*P<0.05,**P<0.01,***P<0.001,and ****P<0.0001.CON:control;DSS:dextran sulfate sodium;ACO:Amomum compactum Sol.ex Maton volatile oil.
3.3.1,8-cineole reduced the ratio of macrophages in the colonic lamina propria (LP) and MLN and inhibited the M1 polarization of macrophages in colitis model mice
Based on the previous flow cytometry and routine blood test results,we suggest that 1,8-cineole may relieve colonic inflammation by regulating the frequency and number of monocytes.Therefore,we further analyzed the proportion of monocyte subsets,such as macrophages (CD11b+,F4/80+),neutrophils (CD11b+,LY6G+),dendritic cells (CD11b+,CD11c+),and antigen presenting cells(CD11b+,MHCII+).The results showed that the proportions of most monocyte subsets in MLN and LP were significantly decreased after 1,8-cineole treatment,especially macrophages,which play a very important role in the development of colitis and are functionally heterogeneous (Fig.3A).Therefore,we used CD86 and CD206 antibodies to mark the M1 macrophages and M2 macrophages,respectively.As shown in Fig.3B,after 1,8-cineole treatment,the proportion of M1 macrophage was significantly reduced in the MLN and LP,but the proportion of M2 macrophages was only increased in the MLN and did not change much in the LP.Immunofluorescence staining also showed that the expressions of F4/80 and CD86 was significantly upregulated in the colon LP of the DSS group,but ACO and 1,8-cineole treatment significantly inhibited the infiltration and M1 polarization of macrophages(Fig.3C).The expression level of inducible nitric oxide (NO) synthase (iNOS),another key marker of M1 polarization,also verified this hypothesis.We found that ACO and 1,8-cineole significantly inhibited iNOS fluorescence intensity in the colon LP (Fig.3D).Then,we purified MLN macrophages through CD11b magnetic bead separation and measured the messenger RNA (mRNA) expression levels related to M1 and M2 polarization by quantitative PCR.Primer sequences are listed in Table S2.The results showed that 1,8-cineole significantly inhibited the mRNA expression levels ofinterleukin(Il)1β,tumor necrosis factor(Tnf)α,Il6,Il18,andIl23,all of which are considered to be key cytokines in M1 polarization of macrophages.Interestingly,the expression levels of some M2 polarization-related factors,such asC-X-C motif chemokine ligand22(Cxcl22)andIl10,were also inhibited by 1,8-cineole(Fig.3E).This result is consistent with the effect of ACO and 1,8-cineole on CD206 expression in the LP.Then,we analyzed the plasma cytokines related to the polarization of macrophages by Bio-Plex™(Bio-Rad Laboratories).As shown in Fig.S5,of the 18 cytokines measured,9 cytokines showed a significant change in the plasma of the colitis group mice compared with the CON group mice.They are mainly proinflammatory cytokines,including TNFα,macrophage inflammatory protein2 (MIP-2),monocyte chemoattractant protein1(MCP-1),KC (CXCL1),IL17A,IL6,IL1β,IFNγ,and granulocytemacrophage colony-stimulating factor (GM-CSF).Compared with the DSS group,the 1,8-cineole-treated group showed significantly inhibited expression of these cytokines.Therefore,we indicated that ACO and 1,8-cineole may relieve intestinal inflammation by inhibiting the polarization of macrophages to the M1 phenotype.
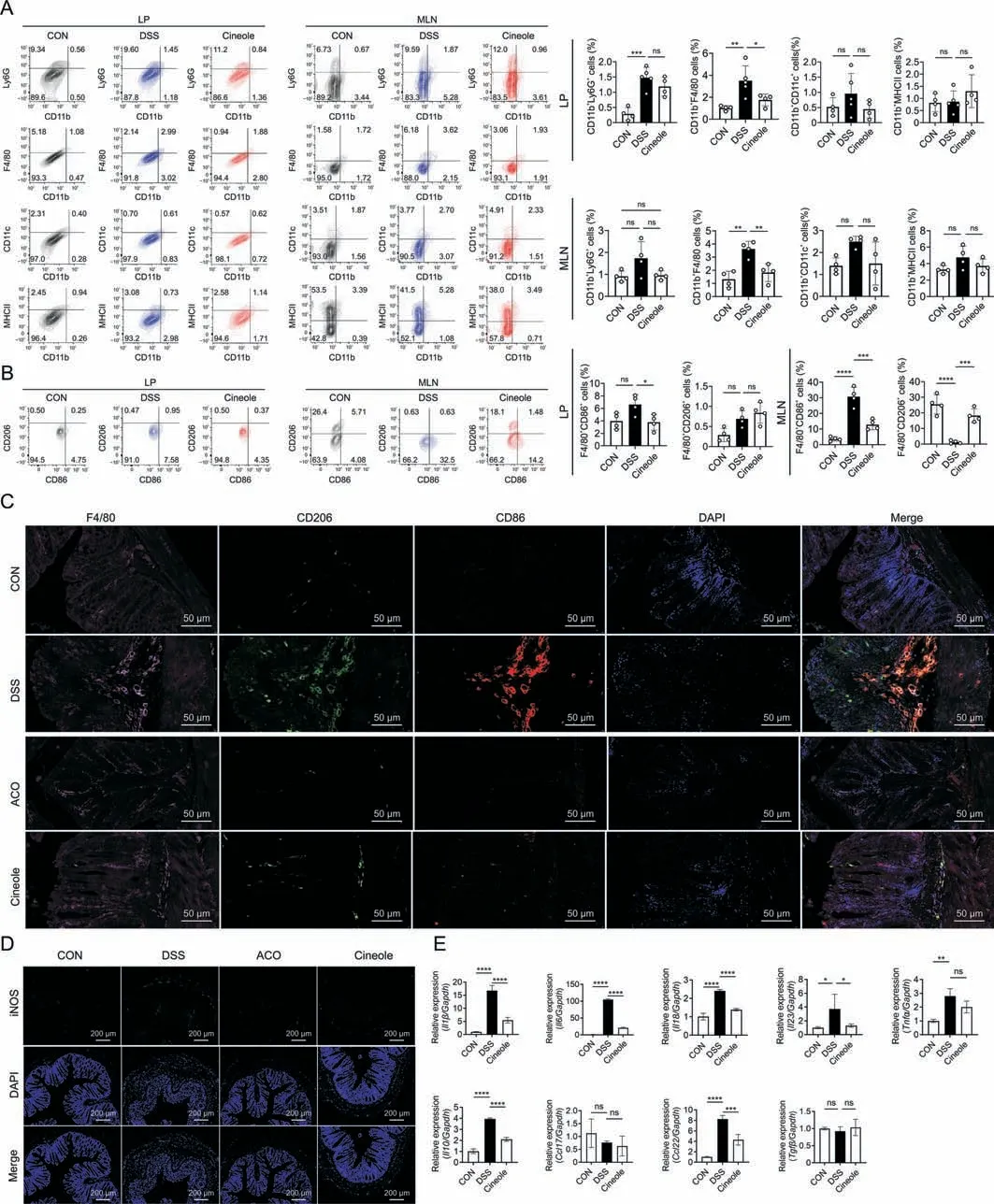
Fig.3. 1,8-cineole (Cineole) inhibited M1 polarization of macrophages in the mesenteric lymph node (MLN) and lamina propria (LP).(A) The percentage of neutrophils (CD11b+,Ly6G+),macrophages(CD11b+,F4/80+),dendritic cells(CD11b+,CD11c+),and antigen-presenting cell(CD11b+,MHCII+)in LPs and MLNs.(B)Flow cytometry for CD86 and CD206 on F4/80+cells in the MLNs and LPs.(C)Representative immunofluorescence staining of F4/80,CD86,CD206,and 4′,6-diamidino-2-phenylindole(DAPI)in colon sections(pink:F4/80;red:CD86;green:CD206;and blue:DAPI).(D)Representative immunofluorescence staining of inducible nitric oxide(NO)synthase (iNOS)and DAPI staining in colon sections(green:iNOS and blue:DAPI).(E)Expression levels of M1 macrophage-related genes(interleukin (Il)1β,tumor necrosis factor (Tnf)α,Il6,Il18,and Il23)and M2 macrophage-related genes(C-C motif chemokine ligand(Ccl)17,Ccl22,Il10,and transforming growth factor beta(Tgfβ)in CD11b+cells sorted by CD11b magnetic beads derived from MLNs.All experiments were performed at least in quadruplicate,and the data represent mean±standard deviation.ns:no statistical significance.*P<0.05,**P<0.01,***P<0.001,and ****P<0.0001.CON:control;DSS: dextran sulfate sodium;ACO: Amomum compactum Sol.ex Maton volatile oil. Gapdh: glyceraldehyde 3-phosphate dehydrogenase.
3.4.1,8-cineole inhibited LPS/IFNγ-induced M1 polarization in vitro
Next,we detect the toxicity of 1,8-cineole to macrophages(Fig.S6) and explored its activity in inhibiting M1 polarization of macrophages in vitro.As shown in Fig.4A,the flow cytometry results demonstrated that the expression of CD86 was significantly upregulated in RAW264.7 cells and BMDMs after LPS stimulation and reversed by 100 μM 1,8-cineole treatment.However,there were no significant changes in M2 polarization(CD206+)after LPS stimulation or treatment with 1,8-cineole (Fig.4A).In addition,cotreatment with 100 μM 1,8-cineole decreased the LPS-induced accumulation of NO and reactive oxygen species (ROS) and inhibited iNOS enzymatic activation in RAW264.7 cells and BMDMs(Figs.4B and C).Moreover,the release of the typical cytokines of M1 macrophage,such as IL6,IL1β,and TNFα,in the supernatant was significantly decreased by 1,8-cineole treatment (Fig.4D).The reverse transcription quantitative PCR results showed that LPS increased the transcription levels of M1 polarization-related genes,includingIl1β,Il6,Tnfα,Il23,andIl18.Additionally,treatment with 1,8-cineole significantly reduced the expression levels of these genes.In contrast,treatment with LPS or 1,8-cineole did not influence the expression level of most genes related to M2 polarization,such asIl10,transforming growth factor beta (Tgfβ),andCcl17.Only theCcl22 mRNA expression level was significantly increased after stimulated with LPS and IFNγ,and this increase was reversed by 200 μM 1,8-cineole(Fig.4E).Collectively,these results indicated that 1,8-cineole may inhibit the polarization of macrophages to the M1 phenotype in vitro.

Fig.4. 1,8-cineole (Cineole) inhibited M1 polarization and proinflammatory cytokine expression in macrophages in vitro.(A) The percentages of CD86+ cells and CD206+ cells in RAW264.7 cells and bone marrow-derived macrophages (BMDMs) after treatment with lipopolysaccharide (LPS) and interferon γ (IFNγ) with or without Cineole.(B) Effects of Cineole on nitric oxide production in LPS-activated RAW264.7 and BMDMs.(C) The levels of inducible nitric oxide (NO) synthase (iNOS) enzymatic activity and reactive oxygen species (ROS) in RAW264.7 cells and BMDMs were determined using flow cytometry.(D) The productions of interleukin (IL) 1 beta,IL6,tumor necrosis factor (TNF) alpha were measured using enzyme linked immunosorbent assay.(E) The expression levels of M1 macrophage-related genes (Il1β, Tnfα, Il6, Il18,and Il23) and M2 macrophage-related genes(C-C motif chemokine ligand(Ccl)17,Ccl22,Il10,and transforming growth factor beta(Tgfβ))in RAW264.7 cells and BMDMs were measured by real time quantitative polymerase chain reaction.All experiments were performed at least in triplicate,and the data represent mean±standard deviation.ns:no statistical significance.*P<0.05,**P<0.01,***P<0.001,and****P <0.0001.CON: control; Gapdh: glyceraldehyde 3-phosphate dehydrogenase.
3.5.1,8-cineole regulated Caco2 monolayer permeability in a differentiated Caco2/THP-1 coculture model
As previously mentioned,we found that 1,8-cineole could inhibit the increase in intestinal permeability and reverse the downregulation of intestinal tight junction protein expression in vivo.However,whether these effects are related to the inhibition of M1 macrophage polarization by 1,8-cineole remains unclear.Therefore,we established a Caco2/THP-1 coculture model [12] to explore the anti-inflammatory and barrier-stabilizing effects of 1,8-cineole (Fig.S7).The apparent permeability coefficient of fluorescein and leakage of FD4 were significantly increased after stimulation with LPS and IFNγ,which previously have been shown to be associated with the increased levels of proinflammatory cytokines released by THP-1 cells [12] (Figs.5A-C).Moreover,the significantly decreased trans-epithelial electrical resistance value after LPS stimulation also showed a diminished trend in the barrier function of the intestinal epithelial cell monolayers (Fig.5D).However,treatment with 1,8-cineole significantly reversed the increase in epithelial permeability(Figs.5A-D).As shown in Figs.5E and F,the protein expression of occludin and ZO1 were both decreased after stimulation with LPS and IFNγ,and the change in occludin expression was more obvious according to the result.Consistent with the previous intestinal permeability results,1,8-cineole significantly reversed the LPS-induced downregulation in tight junction protein expression in Caco2 cells,indicating that 1,8-cineole may maintain intestinal barrier function by inhibiting the release of proinflammatory cytokines from macrophages and regulating the expression of tight junction proteins.
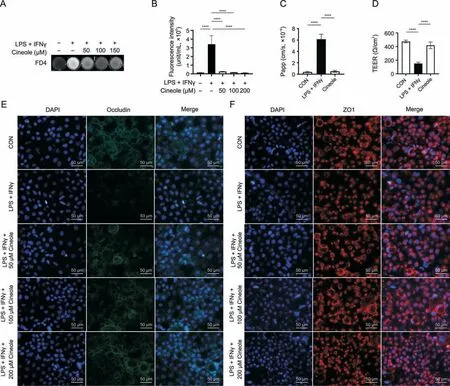
Fig.5. 1,8-cineole (Cineole) restored the Caco2 monolayer barrier function in a differentiated Caco2/THP-1 coculture model.(A) The leakage level of fluorescein isothiocyanatedextran 4 kDa(FD4)in culture medium was photographed by fluorescence imager(white shows fluorescence).(B)Effect of Cineole on leakage of FD4.(C,D)Apparent permeability coefficient(Papp)of fluorescein and transepithelial electrical resistance(TEER)of Caco2 monolayer.(E,F)Representative immunofluorescence staining of occludin(green)(E),ZO1(zonula occludens-1) (red) (F) and 4′,6-diamidino-2-phenylindole (DAPI) staining in Caco2 cells.All experiments were performed at least in triplicate,and the data represent mean ± standard deviation.ns: no statistical significance.****P <0.0001.LPS: lipopolysaccharide;IFNγ: interferon-γ;CON: control.
3.6.1,8-cineole bound to HSP90 and inhibited its ATPase activity
In previous experiments,we verified that 1,8-cineole could significantly inhibit M1 polarization in macrophages in vitro and in vivo,but its exact mechanism and target molecule are not clear.Therefore,the DARTS assay was used to explore the initial target molecule of 1,8-cineole (Fig.6A).According to the Coomassie brilliant blue staining result (Fig.S8),we found a band at approximately 70-95 kD that was significantly thickened after 1,8-cineole treatment compared with the vector.Then,the molecule of this band was identified as HSP90 (90 kDa) using LC-MS.Western blotting also confirmed this result,and the stability of HSP90 was significantly improved by 1,8-cineole compared with the vector treatment(Fig.6B).CETSA and thermal shift results further verified that 1,8-cineole significantly enhanced the thermal stability of HSP90,suggesting that 1,8-cineole could bind to HSP90 (Figs.6C and D).Then,we examined the HSP90 ATPase activity and found that 1,8-cineole significantly reduced HSP90 ATPase activity in a dose-dependent manner.Thus,1,8-cineole may act as an inhibitor of HSP90 ATPase activity (Fig.6E).Next,we investigated the changes in the expression level of HSP90 and its client proteins in macrophages after 1,8-cineole treatment.As shown in Fig.6F,the mRNA expression levels ofHspa1bandHsp90aa1significantly increased in the DSS group.However,compared with the DSS group,there was no significant change in the expression levels ofHspa1a,Hsp90aa1,andHsp90ab1in the Cineole group.Only the expression level ofHspa1bwas significantly reversed by treatment with 1,8-cineole.In vitro,compared with LPS+IFNγ group,the levels ofHspa1ain BMDMs and RAW264.7 cells were increased after 1,8-cineole treatment (Fig.6G),which may be caused by a compensatory increase in transcription after HSP90 ATPase inhibition.The expression of HSP90 client proteins,such as phosphorylated signal transducer and activator of transcription 3 (p-STAT3),phosphorylated protein kinase B (p-AKT),and tumor protein p53(P53),was also downregulated after 1,8-cineole treatment(Fig.6H).Then,we used Autodock Vina to perform molecular docking between HSP90 and 1,8-cineole and found that the binding energy of 1,8-cineole and HSP90 was -4.7 Kal/mol (Fig.6I).Moreover,its key metabolite in the body,2-hydroxy-1,8-cineole[13],has a stronger binding capacity than that of 17-allylamino-17-demethoxygeldanamycin.To test whether HSP90 is the main target of 1,8-cineole for regulating macrophage M1 polarization,we investigated the activity of 1,8-cineole by overexpressing HSP90.The results showed that after overexpressing HSP90 in RAW264.7 cells,the inhibitory effect of 1,8-cineole on M1 polarization was significantly reduced(Figs.S9A-C).Thus,we suspected that 1,8-cineole may inhibit macrophage M1-type activation by suppressing HSP90 activity.
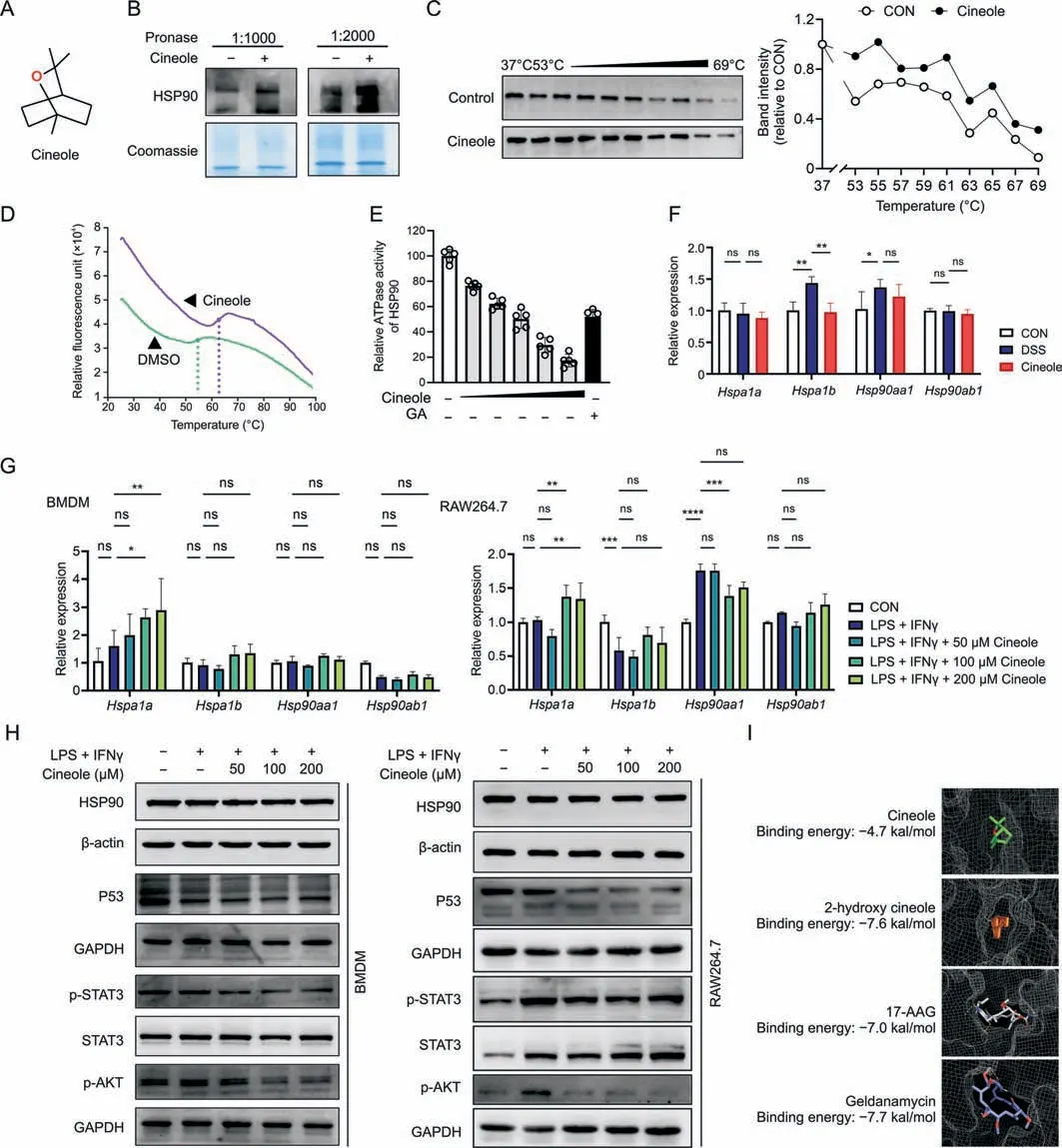
Fig.6. Heat shock protein 90 (HSP90) was identified as a potential target for 1,8-cineole (Cineole).(A) Chemical structure of Cineole.(B) Lysates from RAW264.7 cells were incubated with or without Cineole(100 μM)overnight and different proportions of pronase were added for 15 min,HSP90 levels were determined using Western blotting and was normalized with Coomassie brilliant blue staining.(C) Lysates from RAW264.7 cells were incubated with or without Cineole (100 μM) overnight and were subjected to cellular thermal shift assay.(D)The interaction between Cineole and HSP90 was determined via thermal shift assay.The purple lines represent incubation with Cineole and the green lines represent incubation with dimethyl sulfoxide (DMSO).(E) Effect of Cineole on the adenosinetriphosphatases activity of HSP90.(F) Expression of Hspa1a, Hspa1b, Hsp90aa1,and Hsp90ab1 in CD11b magnetic beads purified mesenteric lymph node (MLN) macrophages.(G) Expression of Hsp90aa1, Hsp90ab1, Hspa1a,and Hspa1b in RAW264.7 cells and bone marrow-derived macrophages(BMDMs)after treated with lipopolysaccharide(LPS)/interferon-γ(IFNγ)and different concentrations of Cineole(50,100,and 200 μM).(H)Effects of Cineole on the expressions of HSP90 and its client proteins.(I) Molecular docking of HSP90 binding of Cineole.All experiments were performed at least in triplicate,and the data represent mean ± standard deviation.ns: no statistical significance.*P <0.05,**P <0.01,***P <0.001,and ****P <0.0001.CON: control;ATPase: adenosine triphosphatases;GA:geldanamycin;DSS: dextran sulfate sodium;p53: tumor protein p53;GAPDH: glyceraldehyde-3-phosphate dehydrogenase;p-STAT3: phosphorylated signal transducer and activator of transcription 3;p-AKT: phosphorylated protein kinase B;17-AAG: 17-allylamino-17-demethoxygeldanamycin.
3.7.1,8-cineole attenuated DSS-induced colitis by regulating NODlike receptor pathway
Based on the aforementioned results,we found that 1,8-cineole could regulate HSP90 activity and inhibit its related downstream pathways.Next,we performed RNA-sequencing to explore its downstream targets in M1 polarization.As shown in Fig.7A,the gene expression of the 1,8-cineole group was more similar with that of the CON group than of the DSS group,and the expression levels of 2313 upregulated genes and 1247 downregulated genes in the DSS group were reverse regulated by 1,8-cineole,which indicated that 1,8-cineole significantly inhibited the genetic changes caused by DSS (Figs.7B and C).Gene ontology (GO) and Kyoto encyclopedia of genes and genomes (KEGG) analyses showed that 1,8-cineole mainly regulated immune-related pathways,such as the regulation of phagocytosis engulfment (Fig.7D) and hematopoietic cell lineage (Fig.7E).Then,we used weighted gene coexpression network analysis (WGCNA) to identify the 1,8-cineoleassociated coexpressive modules and hub genes.The results showed that the“MEbrown”modules were most closely related to the 1,8-cineole treatment groups (Figs.7F and S10).According to the KEGG analysis of hub genes in “MEbrown”modules,we found that these genes were highly enriched in the NOD-like receptor pathway(Fig.7G).The gene set enrichment analysis(GSEA)results indicated that cineole significantly downregulated the NOD-like receptor pathway (Fig.7H).As shown in the heatmap,1,8-cineole decreased the expression levels of most genes in the NOD-like receptor pathway,especially the expression level ofNlrp3,which is also the key client protein of HSP90 (Fig.7I).Therefore,we hypothesized that 1,8-cineole may inhibit DSS-induced colitis mainly by inhibiting the activation of NLRP3 inflammasome.
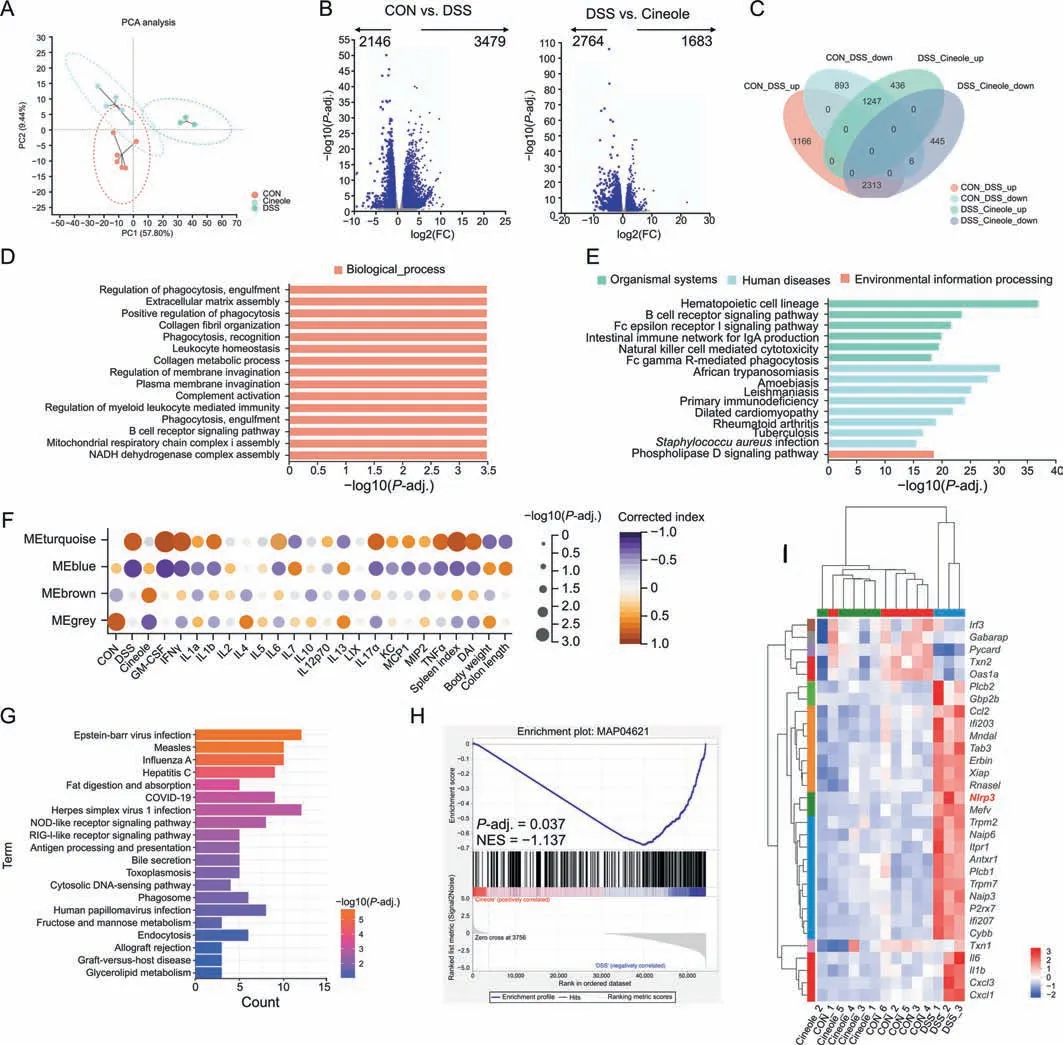
Fig.7. Bioinformatics analysis showed that 1,8-cineole (Cineole) alleviated dextran sulfate sodium (DSS)-induced colitis by inhibiting nucleotide-binding and oligomerization domain(NOD)-like receptor pathway.(A)Principal component analysis(PCA)plot of the messenger RNA(mRNA)expression profile.(B)Volcano analysis of differentially expressed genes(DEGs)of the control(CON)group(n=6)vs.DSS group(n=3)or DSS group vs.Cineole group(n=5).(C)Venn diagram showing the overlapping DEGs between CON group vs.DSS groups or DSS groups vs.Cineole group.(D,E)Gene ontology(GO)terms and Kyoto Encyclopedia of Genes and Genomes(KEGG)pathway enriched for overlapped DEGs.(F)Correlations between weighted gene co-expression network analysis modules and colitis mice phenotypes.(G)KEGG pathway analysis for brown module.(H)Gene set enrichment analysis(GSEA) showed that Cineole inhibited the NOD-like receptor signaling pathway(MAP04621).(I) Heat map showed the genes in the NOD-like receptor signaling pathway which was regulated by Cineole (fold change (FC) >2, P <0.05).NADH: nicotinamide adenine dinucleotide;GM-CSF: granulocyte-macrophage colony stimulating factor;IFNγ:human interferon-γ;LIX:C-X-C motif chemokine ligand 5;KC:C-X-C motif chemokine ligand 1;MCP1:monocyte chemotactic protein 1;MIP2:C-X-C motif chemokine ligand 2;TNFα: tumor necrosis factor α;DAI: disease activity index;COVID-19: coronavirus disease 2019;RIG1: retinoic acid-inducible gene I;NES: normalized enrichment score.
3.8.1,8-cineole inhibited the binding of HSP90 to NLRP3 and suppressed the activation of the NLRP3 pathway in macrophage polarization
To further explore the effect of 1,8-cineole on NLRP3 activation,we used Western blotting to determine the expression levels of the NLRP3,caspase 1,and other NLRP3 inflammasome components.The results suggested that the expression levels of NLRP3,apoptosisassociated speck-like protein containing a caspase recruitment domain (ASC),pro-caspase1,and pro-IL1β were significantly decreased after 1,8-cineole treatment in vivo and in vitro (Figs.8A-C).These data indicated that 1,8-cineole conspicuously inhibited NLRP3 inflammasome activation.It has been reported that HSP90 and its client-adaptor SGT1 can directly bind to NLRP3 to enhance NLRP3 stability and that HSP90 inhibitors significantly suppress the activation of NLRP3 inflammasome (Fig.8D) [14].Therefore,we further investigated how the interaction among HSP90,SGT1,and NLRP3 was disrupted by 1,8-cineole.The results suggested that 1,8-cineole may inhibit the formation of the HSP90,NLRP3,and SGT1 complex,leading to the degradation of NLRP3(Fig.8E).To confirmed that 1,8-cineole inhibits macrophage M1 polarization through the regulation of the NLRP3 inflammasome,we transfected RAW264.7 cells with small interfering RNAs to knock down NLRP3 and SGT1.Western blot results showed that compared with the those in the small interfering RNA negative control (Si-NC) group,the expression of SGT1 and NLRP3 were significantly decreased in the Si-SGT1 group and Si-NLRP3 group,respectively,after knockdown,but there was no obvious cross-interference (Fig.9A).After LPS-and IFNγ-induced M1 polarization of macrophages,there was no significant difference in the level of iNOS in the cells and in the level of IL1β in the supernatant after applying 1,8-cineole in the NLRP3 knockdown group and SGT1 knockdown group (Figs.9B and C).Additionally,the flow cytometry results showed that knocking down both SGT1 and NLRP3 significantly suppressed the inhibitory effect of 1,8-cineole on M1 polarization of macrophages(Fig.9D).Overall,these results illustrated that 1,8-cineole could inhibit the NLRP3 inflammasome by regulating the HSP90 activity.
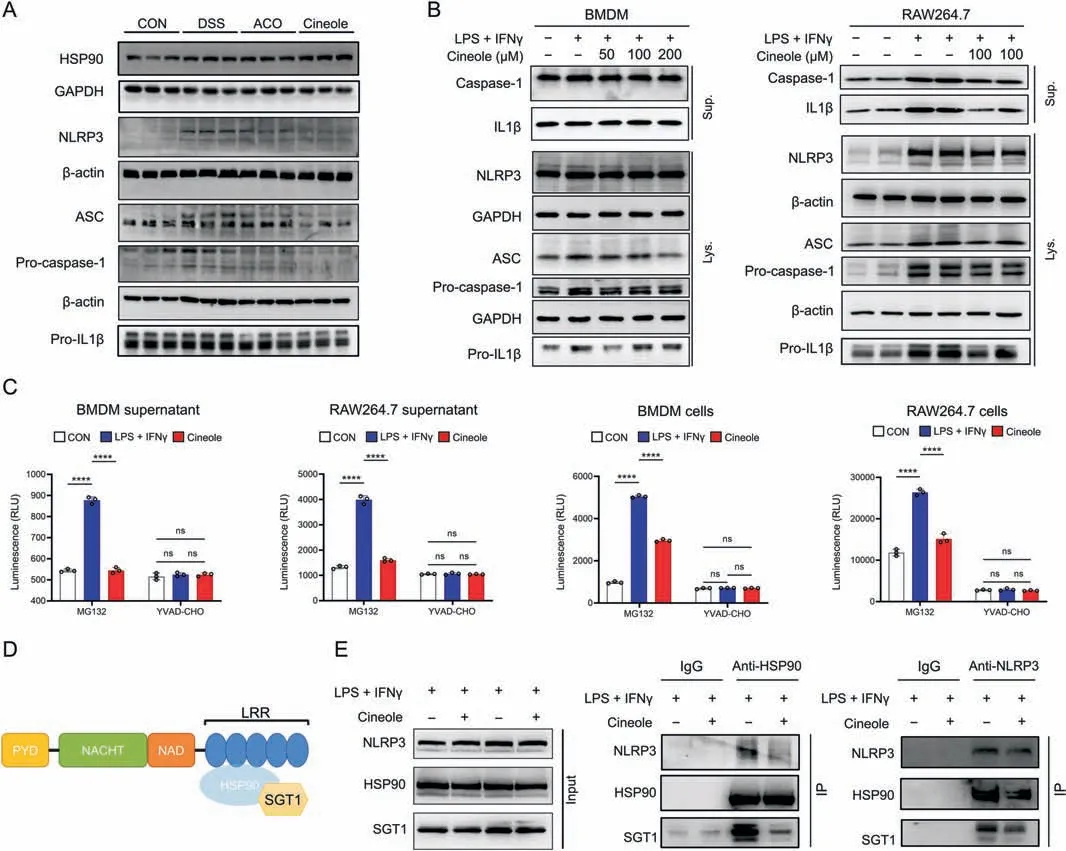
Fig.8. 1,8-cineole(Cineole)inhibited the binding of heat shock protein 90(HSP90)to nucleotide-binding and oligomerization domain(NOD)-,leucine rich repeat(LRR)-,and pyrin domain (PYD)-containing 3 (NLRP3) and the activation of NLRP3 inflammasome.(A,B) The expression level of proteins associated with NLRP3 inflammasome activation in colon tissues (A) and BMDMs and RAW264.7 cells (B).(C) Caspase-1 activity of bone marrow-derived macrophage (BMDM) supernatant,RAW264.7 supernatant,BMDMs,and RAW264.7 cells supernatant,which was assessed by Caspase1-Glo®assay.Mg-132 is a proteasome inhibitor and YYAD-CHO is a caspase-1 select inhibitor that enables to measure caspase-1 activity specificity.(D)Schematic representation of the domains in NLRP3 interacting with HSP90 and suppressor of G-two allele of SKP1(SGT1)[14].Reprinted from Ref.[14] with permission.(E) Co-immunoprecipitation (Co-IP) experiments to validate the effect of Cineole on the interaction of NLRP3 to HSP90 and SGT1.All experiments were performed at least in triplicate,and the data represent mean ± standard deviation.ns: no statistical significance.****P <0.0001.GAPDH: glyceraldehyde-3-phosphate dehydrogenase;ASC: apoptosis-associated speck-like protein containing a CARD;Pro-IL1β: pro-interleukin (IL)-1 beta;CON: control;DSS: dextran sulfate sodium;ACO: Amomum compactum Sol.ex Maton volatile oil;LPS:lipopolysaccharide;IFNγ:interferon-γ;Sup.:supernatant;Lys.:lysate;PYD:pyrin domain;NACHT:domain present in NAIP,CIITA,HET-E,and TP1;NAD: nicotinamide adenine dinucleotide.

Fig.9. Suppressor of G2 allele of skp1 (SGT1) and nucleotide-binding and oligomerization domain (NOD)-,leucine rich repeat (LRR)-,and pyrin domain-containing protein 3(NLRP3) knockdown weakened the inhibitory effect of 1,8-cineole (Cineole) on the polarization of M1 macrophages.(A) Western blotting results showing the expression of SGT1 and NLRP3.(B)The level of inducible nitric oxide(NO)synthase(iNOS)enzymatic activity in RAW264.7 cells was determined using flow cytometry.(C)Production of interleukin 1 beta (IL1β) was measured using enzyme-linked immunosorbent assay.(D) The percentages of CD86+ cells and CD206+ cells in RAW264.7 cells were detected by flow cytometry assay.All experiments were performed at least in triplicate,and the data represent mean±standard deviation.ns:no statistical significance.*P<0.05 and ****P<0.0001.GAPDH:glyceraldehyde 3-phosphate dehydrogenase;Si-NC:small interfering(si-)RNA negative control;LPS:lipopolysaccharide;IFNγ:interferon-γ;IL1β:interleukin-1 beta;CON:control.
4.Discussion
UC is a chronic inflammatory disease that gradually affects the rectum and part or the entire colon,seriously threatens patient quality of life.Although there are some clinical solutions for patients with mild and moderate UC,studies have shown that only approximately 40% of patients achieve clinical remission [15].Additionally,there are some limitations of medical treatment,such as severe side effects,a long treatment course,and unsatisfactory therapeutic effect.Therefore,a more effective treatment for UC is urgently needed.In previous studies,we found that ACO and its main compounds 1,8-cineole can significantly relieve intestinal barrier damage in pneumonia model mice (unpublished observations),but their underlying mechanism is still unclear.In this study,we demonstrated that ACO and 1,8-cineole similarly reduce inflammation in UC model mice.In addition,we found that 1,8-cineole could regulate the secretion of proinflammatory factors by inhibiting HSP90-NLRP3-SGT1 complex binding,which in turn inhibited M1 macrophage polarization,which reduced the damage to intestinal barrier function and alleviated colonic inflammation.Overall,these data indicated that 1,8-cineole could be a new candidate drug for UC treatment.
Intestinal mucosal barrier is one of the typical pathological features of UC[16].Decreased secretion of intestinal mucus and loss of intestinal tight junction proteins may result in increased intestinal permeability,which promotes exposure to luminal content and triggers an immunological response [17,18].Consistent with previous studies,DSS group mice showed significantly increased intestinal permeability,decreased goblet cell number,intestinal recess destruction,and downregulated tight junction protein.However,treatment with ACO and 1,8-cineole could significantly ameliorate the damage to intestinal epithelial cells,decrease intestinal permeability,increase goblet cell number,and increase the mucus secretion,which suggests that both ACO and 1,8-cineole can restore the intestinal barrier and protect the intestinal mucosa.However,this effect was not a result of increasing intestinal epithelial proliferation according to immunohistochemical result of Ki67.Therefore,we hypothesized these results may be related to their anti-inflammatory effect and investigated the levels of inflammatory factors in plasma and the proportion of immune cells in the colonic LP and MLNs.
Both the levels of plasma inflammatory cytokines and immune cells in UC model mice showed that ACO and 1,8-cineole had excellent anti-inflammatory effects.Flow cytometry and routine blood results showed that ACO and 1,8-cineole could significantly reduce the proportion of monocytes and neutrophils in the blood of UC mice,which play important roles in promoting the inflammatory response of UC [19].Further analysis of monocyte subpopulations in LP and MLN using flow cytometry showed that the proportion of macrophages in UC mice was significantly reduced after 1,8-cineole treatment,and 1,8-cineole also inhibited the M1-type macrophages polarization,which is considered as the proinflammatory state of macrophages.It has been reported that M1-type macrophage can promote the aggregation of neutrophils through the release of cytokines such as TNFα and IL1β,thus inducing inflammation in the colon mucosa and promoting the damage of colon barrier[20,21].Subsequently,we used Bio-Plex™analyses to evaluate the inflammatory factors related to macrophage polarization in serum,and the results also confirmed the above findings.After 1,8-cineole treatment,the serum levels of TNFα,MIP-2,MCP-1,KC,IL17A,IL6,IL1β,IFNγ,and GM-CSF in UC model mice were significantly decreased.GM-CSF is considered to be secreted by a variety of cells,mainly macrophages and T cells.Mast cells,natural killer cells,and others play an important role in promoting the M1 polarization of macrophages and the maturation and chemotaxis of neutrophils [22].TNFα,IL6,IL1β,and IFNγ are considered to be the key inflammatory factors released during the polarization of M1 macrophages[23].MCP-1 and MIP-2 are the key chemokines that regulate the migration and infiltration of monocytes/macrophages[24].The above results indicate that 1,8-cineole significantly inhibits the M1 polarization-related cytokines in macrophages and has a significant regulatory effect on macrophage migration and infiltration as well as M1 polarization.All these cytokines have been reported to impair intestinal barrier function by inducing intestinal epithelial cell injury and inhibiting the expression of tight junction proteins [25-27].Furthermore,macrophage polarization experiments and coculture experiments in vitro also verified the inhibitory effect of 1,8-cineole on macrophage M1 polarization and confirmed that reducing inflammatory factors released by M1 polarization can indeed significantly improve the damage to intestinal barrier function.
DARTS is a method to explore molecular binding targets based on the stability of protein enzymatic hydrolysis,which has been used to search for binding targets of various small molecules[9].To further verify the specific targets of 1,8-cineole inhibiting macrophage M1 polarization,we used LC-MS and Western blotting to determine the protein expression in the samples after the DARTS experiment,and we found that the protein stability of HSP90 was significantly increased after the treatment with 1,8-cineole.Furthermore,we verified the potential binding between 1,8-cineole and HSP90 by using CETSA,thermal shift assay,and ATPase activity assays and found that the binding between 1,8-cineole and HSP90 showed pharmacological activity.HSP90 is a key chaperone molecule that aids in the folding,stabilization,and remodeling of more than 400 proteins,referred to as HSP90“clients”,many of which are involved in the polarization of macrophages and mainly include transcription factors (such as nuclear factor kappa-B and STAT3)and kinases (such as AKT).These HSP90 clients are critical for the sensing,transmission,and amplification of inflammatory signals and the transcription of inflammatory factors during macrophage polarization.Thus,we measured the expression of inflammatoryrelated client proteins and determined that 1,8-cineole could inhibit the levels of inflammatory-related client proteins of HSP90,including P53,p-STAT3,and p-AKT.P53 is one of the classical client proteins of HSP90 and plays an important role in tumorigenesis.Recent studies have also shown that P53 may be involved in the process of macrophage M1 polarization induced by ROS overproduction.The increase of ROS levels in macrophages can significantly induce the expression of P53 and promote its acetylation[28].Moreover,another HSP90 client protein,STAT3,which binds to the N-terminal region of HSP90 protein to maintain its phosphorylation,is considered as a key player in regulating macrophage phenotype.It has been reported that after LPS stimulation,the phosphorylation and nuclear transcription of STAT3 in macrophages can be significantly increased,which leads to the M1-polarized phenotype of macrophages and the release of IL1β [29].AKT is one of the first identified client proteins of HSP90 and plays an important role in regulating macrophage polarization and promoting inflammation [30].High levels of ROS during colitis can activate the phosphoinositide 3-kinase/AKT signaling cascade,thereby promoting AKT phosphorylation and subsequent NFκB activation,thus regulating the ratio of M1 to M2 macrophages[31,32].In this study,we found that the levels of several HSP90 clients involved in macrophage polarization were significantly inhibited after 1,8-cineole treatment,suggesting that 1,8-cineole may have a suitable inhibitory effect on HSP90.Furthermore,we overexpressed HSP90 in RAW264.7 cells,and the results also showed that the anti-M1 polarization effect of 1,8-cineole was significantly weakened after overexpression,indicating that HSP90 may be the main target of 1,8-cineole in the anti-M1 polarization of macrophages.However,the interaction between 1,8-cineole and HSP90 is still unclear and will be further explored in the future.
NLRP3 is an essential component of the innate immune system and acts as a pattern recognition receptor in macrophages and promotes M1 polarization of macrophages[33].It has been proven to be an important client protein of HSP90 and plays an important role in thepathogenesis of inflammatory bowel disease.Studies have shown that inhibition of HSP90 can inhibit the activation of NLRP3 in the intestine,thereby reducing the release of proinflammatory factors and alleviating the related symptoms of colitis[11].In our study,we first found that the NOD-like receptor pathway was greatly altered after 1,8-cineole treatment by transcriptome sequencing,and found that the expression of NLRP3 and activation-related proteins such as ASC,pro-caspase 1,and pro-IL1 was significantly downregulated after 1,8-cineole treatment,and the release of IL1β and Caspase1 was also significantly reduced in the supernatant of BMDMs and RAW264.7 medium.These results indicated that NLRP3 activation was inhibited by 1,8-cineole.Recent studies have shown that inhibiting of HSP90 can significantly reduce NLRP3-related inflammatory response mainly because NLRP3 is more easily degraded in the absence of HSP90 protection[14].Further studies showed that SGT1,an HSP90 client aptamer,dissociated from the HSP90/NLRP3 complex upon HSP90 inhibitor treatment,thereby partially affecting inflammasome activation [14].Thus,we used co-IP and RNA interference methods to investigate the effect of 1,8-cineole on the binding of HSP90,SGT1,and NLRP3.The results showed that the 1,8-cineole may affect the formation of the HSP90-SGT1-NLRP3 complex to inhibit the M1 polariton of macrophage.
5.Conclusion
In summary,our data demonstrated that 1,8-cineole remarkably inhibited the intestinal barrier damage and increased intestinal permeability caused by DSS and showed great therapeutic potential for the treatment of UC.We also found that 1,8-cineole significantly inhibited the polarization of macrophage of M1 macrophages and the release of proinflammatory factors in the intestinal LP and MLNs,which also promoted the recovery of intestinal epithelial barrier function.Through further verification,we found that 1,8-cineole could inhibit the binding between HSP90 and its key client protein NLPR3,thus inhibiting the M1 polarization of macrophages.These findings provide convincing evidence that 1,8-cineole is a promising candidate in the search for a novel therapeutic strategy for UC treatment.
CRediT author statement
Shengsuo Ma:Conceptualization,Methodology,Software,Formal analysis,Data curation,Writing-Original draft preparation,Reviewing and Editing;Bing Yang:Methodology,Writing-Original draft preparation;Yang Du:Resources,Investigation,Formal analysis,Writing -Original draft preparation;Yiwen Lv:Methodology,Investigation,Data curation,Writing -Original draft preparation;Jiarong Liu:Methodology;Yucong Shi:Investigation;Ting Huang:Resources;Huachong Xu:Supervision,Project administration,Funding acquisition;Li Deng:Supervision,Project administration,Funding acquisition;Xiaoyin Chen:Conceptualization,Supervision,Project administration,Funding acquisition.
Declaration of competing interest
The authors declare that there are no conflicts of interest.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant Nos.: 81830114,82004232,82174253,and 82104707),Guangdong Basic and Applied Basic Research Foundation,China (Grant Nos.: 2021A1515011215 and 2022A1515110827),Guangzhou Basic and Applied Basic Research Foundation,China (Grant No.: 2023A1515011149),China Postdoctoral Science Foundation (Grant Nos.: 2020M683206 and 2021M701443),the Key Area Research and Development Program of Guangdong Province,China (Grant No.: 2020B1111100010),Guangzhou Key Laboratory of Formula-Pattern of Traditional Chinese Medicine,China (Grant No.: 202102010014),the Cross-disciplinary Special Project of Jinan University,China (Grant No.:21621115),the State Key Laboratory of Dampness Syndrome of Chinese Medicine,China (Grant No.: SZ2021KF13),and the Outstanding Innovative Talents Cultivation Funded Programs for Doctoral Students of Jinan University,China (Grant No.:2021CXB024).
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.jpha.2023.07.001.
 Journal of Pharmaceutical Analysis2023年9期
Journal of Pharmaceutical Analysis2023年9期
- Journal of Pharmaceutical Analysis的其它文章
- Comment on: “Development of a CLDN18.2-targeting immuno-PET probe for non-invasive imaging in gastrointestinal tumors”
- JOURNAL OF PHARMACEUTICAL ANALYSIS BEST PAPERS 2022
- Targeted bile acids metabolomics in cholesterol gallbladder polyps and gallstones: From analytical method development towards application to clinical samples
- Characterization of natural peptides in Pheretima by integrating proteogenomics and label-free peptidomics
- Eight Zhes Decoction ameliorates the lipid dysfunction of nonalcoholic fatty liver disease using integrated lipidomics,network pharmacology and pharmacokinetics
- Rapid metabolic fingerprinting with the aid of chemometric models to identify authenticity of natural medicines: Turmeric, Ocimum,and Withania somnifera study