
南昌市城市废弃地再野化时空特征研究
2023-10-18 08:38刘青
中国园林 2023年9期
刘 青
杨坪赫
章昊天
古新仁
刘苑秋*
全球城市化快速扩张、土地利用及人口膨胀所导致的全球环境改变,被普遍认为是物种灭绝、栖息地丧失和气候变化的根本原因[1],特别是近几十年来,人类极大地加速了世界范围内生物多样性的改变和丧失[2]。再野化被认为是解决这类问题的一个可能的方法[3-4],并已从大量的实践和研究中得到印证[5]。在再野化实践及研究过程中,由于实践和研究的逐步深入及其立足点的差异,再野化的概念缺乏一个统一明确的定义,不同学者提出了10余种不同的再野化概念[6],主要包括更新世野化(Pleistocene rewilding)[7]、营养野化(Trophic rewilding)[8]、生态野化(Ecological rewilding)[9]和被动野化(Passive rewilding)[10]。因再野化定义的模糊,导致再野化概念和实践本身也存在较大争议[11],其核心的思辨在于“野化”和“恢复”之间相似的管理活动所引发的边界的模糊性,以及人与自然生态系统间的“内”与“外”关系[12-15]。当前的再野化研究和实践主要基于3种路径:一是对缺失的生态链关键种进行再引入的恢复,特别是顶级肉食动物的再引入[16];二是在现有基础上减少或限制人为干扰及介入的自我恢复;三是“3C”模式,包括核心区(core)、生态廊道(corridor)和食肉动物(carnivore)所构建的保护和恢复更大范围的景观及其生态过程[17-18]。结果表明,3种路径对于场地内的物种丰富度及生态系统恢复均表现出积极的正向效应[19-22]。然而,由于缺乏监测,在试点地点以外再野化的应用和扩大规模的实践研究仍然有限,对于再野化的长期结果仍然知之甚少[23]。城市作为人为干扰及物种丰富度丧失最为严重的场所,其内部场地再野化的时空梯度关系及场地外围环境对再野化的影响,特别是在废弃裸地基础上的再野化时空演变特征缺乏研究。裸地的再野化可以认为是从原有生态系统丧失的起点开始自然恢复的生态过程,对于裸地再野化生态过程的研究,有助于理解再野化过程中人类介入的必要性和价值,以及再野化进程中人与自然生态系统的复杂关系。
出于对以上问题的思考与构想,本研究以南昌市具有城乡梯度关系的东西向8处再野化样地为例,探讨城市环境再野化过程的时空特征及其内部演化机理。
1 研究方法
1.1 研究区概况
南昌市(115°27′~116°35′E,28°10′~29°11′N)位于长江中下游平原,全市面积7402km2,其中水域面积2204km2。全市地势总体西北高、东南低,依次发育低山丘陵、岗地、平原,呈现层状地貌特征,以赣江为界,赣江西北部为构造剥蚀低山丘陵、岗地,赣江以东为河流侵蚀堆积平原。该地区属于亚热带季风气候,气候温暖湿润,四季分明,多年平均气温为17.78℃,多年平均降雨量为1582.81mm。南昌市作为江西省省会城市,为江西省政治、经济、文化中心,据《南昌市统计年鉴》统计数据显示,2021年全市GDP为6650.53亿元,南昌市常住人口625.50万人,其中城镇人口为488.38万人,全市城镇化率达到78.08%。
本次再野化调查样地如图1所示,横跨南昌市东西轴向,由省博物馆为中心,向东延伸为鄱阳湖水域的平原地形,向西延伸为梅岭国家森林公园的山地地形,所选样地能够较好地体现南昌市的地形地貌及城市发展的环境特征。所选样地基本情况如表1所示。
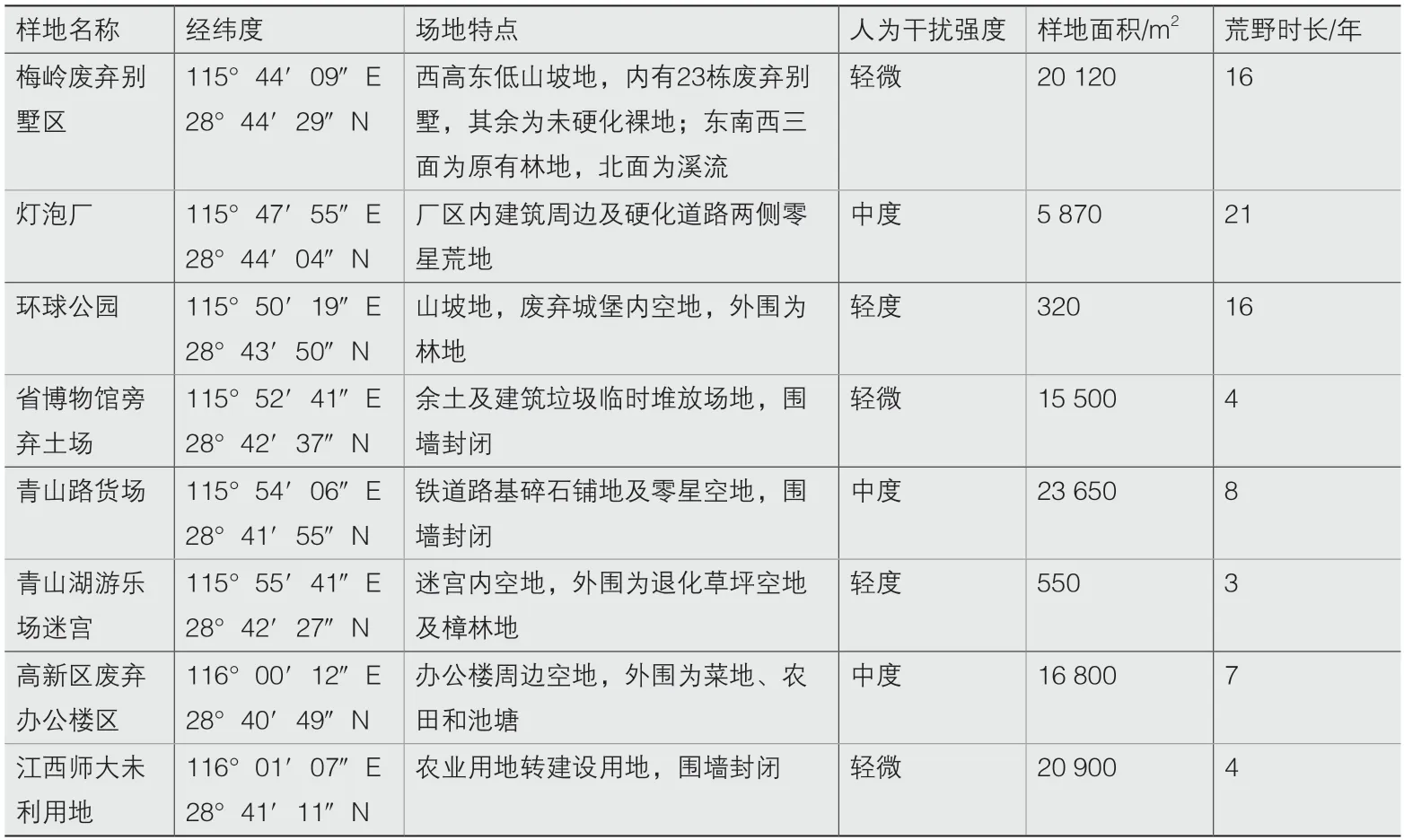
表1 样地基本情况
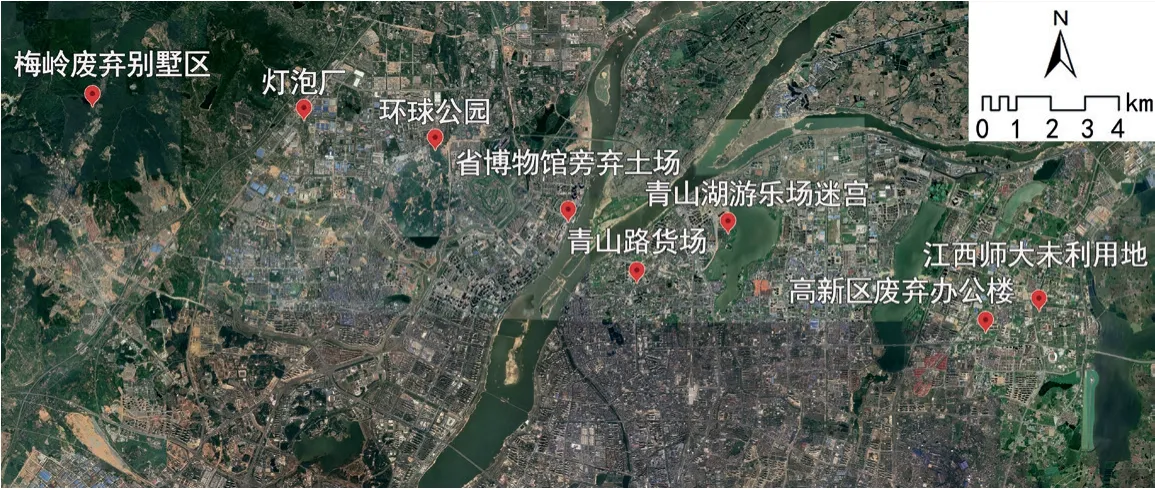
图1 南昌市再野化调查样地分布(底图引自图新地球)
1.2 研究方法与数据获取
采用大疆无人机(大疆精灵4PRO)进行航测,结合大疆智图对所选样地绘制二维正测投影图及三维模型图,以获取样地空间附着物的精确空间数据(如建筑的精确边界,植被的点位、冠幅等信息);依据航测图像及提取信息,人工现场核对修正,并现场对植被加以辨认、实测及统计;利用ArcGIS 10.5对所有乔木树种与场地矗立物做邻近距离提取;用ArcGIS 10.5及SPSS 26对获取数据进行空间分析和统计分析。植物种类的调查方法为:乔木采用全样地调查;灌木、藤本植物采用2m×2m样方抽样调查,草本植物采用1m×1m样方抽样调查,每块样地随机抽取3个样方,植物种类仅调查非人为种植的、再野化过程中自然繁衍的植物。以2和5月的调查数据作为统计分析数据。
2 结果与分析
2.1 基于时空梯度的再野化特征
8处样地的调查结果表明(图2),在城乡梯度上总体表现为物种丰度由城市中心向城郊逐渐递增的趋势,相关性分析也表明(表2),城乡梯度与植物物种总数存在显著相关关系。然而,通过对不同类型植物与城乡梯度进行相关分析可以看出,各类型植物与城乡梯度均呈正相关,但与乔木、灌木和藤本植物的相关性并不显著,仅与草本植物呈极显著相关。
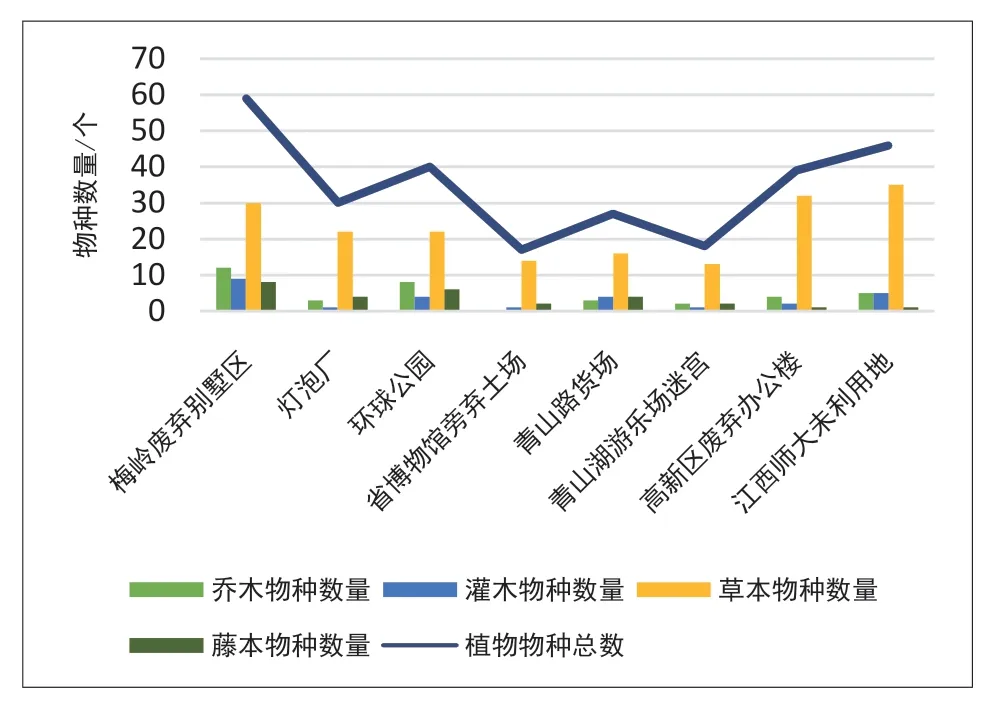
图2 城乡梯度上的物种差异
时间梯度分析表明,荒野时长与植物物种数量有正相关性,但除与藤本植物呈显著相关外,与其他3类植物相关性不显著,与物种总数相关性也不显著。
基于以上分析结果,在时空梯度上,城市再野化表现出以下特征。1)再野化存在城乡梯度效应。在城乡梯度上,由于人为干扰对物种丰富度的负面影响,导致物种丰富度由城郊的近自然环境向城市人工环境呈逐步递减趋势,这种梯度关系影响、制约着再野化进程。2)在完全人为破坏原有生态环境基础上的再野化,其物种丰富度的恢复与时间正相关但并不存在显著的相关性,这说明生态系统的再生恢复是一个缓慢的过程,特别是木本植物的恢复更为缓慢,而草本植物作为传播繁殖能力强的物种,其物种数量前期恢复的速度较快,但后期增长缓慢。3)再野化物种的丰富度受外围环境影响,外围物种丰富将促进再野化场地的物种多样性。4)不同样地的植物种类存在较大差异,这与样地被扰动、改造前的原始性状,以及外围环境和物种传播途径密切相关。
2.2 基于场地特点的再野化特征
在再野化过程中,不同的立地条件及外围环境对场地的再野化产生重要影响(表3)。1)梅岭废弃别墅区与环球公园均为山地,土质为山地原土红壤土,外围均为原有林地,物种丰富,因而其自然再生的乔木、灌木和藤本植物种类较其他样地多。但二者间的物种也存在较大差异,梅岭废弃别墅区蔷薇科植物物种较多,环球公园菊科植物物种较多,原因主要是二者与外部环境的边界分隔方式不同,且由于环球公园内部尚有烧烤营地经营,人为扰动也相对较强。2)灯泡厂和青山路货场大多为建筑、硬化道路或铁道碎石路基,零星未硬化空地土质多为碎石和沙土,土壤贫瘠,乔木树种主要为构树(Broussonetia papyrifera)和白花泡桐(Paulownia fortunei),菊科植物物种也较为丰富。青山路货场的灌木物种多于灯泡厂,主要是因为货场原有绿化灌木,如海桐(Pittosporum tobira)和小叶女贞(Ligustrum quihoui)等的传播。3)省博物馆旁弃土场主要用于堆放工程余土及建筑垃圾,灌木仅有苎麻(Boehmeria nivea)(也是所有样地唯一均有的灌木植物),草本植物主要为加拿大一枝黄花(Solida canadensis)和狗尾草(Setaria viridis),分别占该样地植物总数的56.6%和20.4%,表明入侵植物在城市内这种类型的土地中具有较强的繁衍优势。4)青山湖游乐场迷宫的草本植物主要是耐阴植物井栏边草(Pteris multifda),乔木树种主要是构树和香樟(Cinnamomum camphora)。5)高新区废弃办公楼及江西师大未利用地均由农用地转变而来,地形平坦,土质较为肥沃,乔木树种主要为构树、白花泡桐、香樟和乌桕(Triadica sebifera)等,草本植物较为丰富。2处样地的植物种类也基本相同,主要为菊科、禾本科、大戟科和蓼科植物。
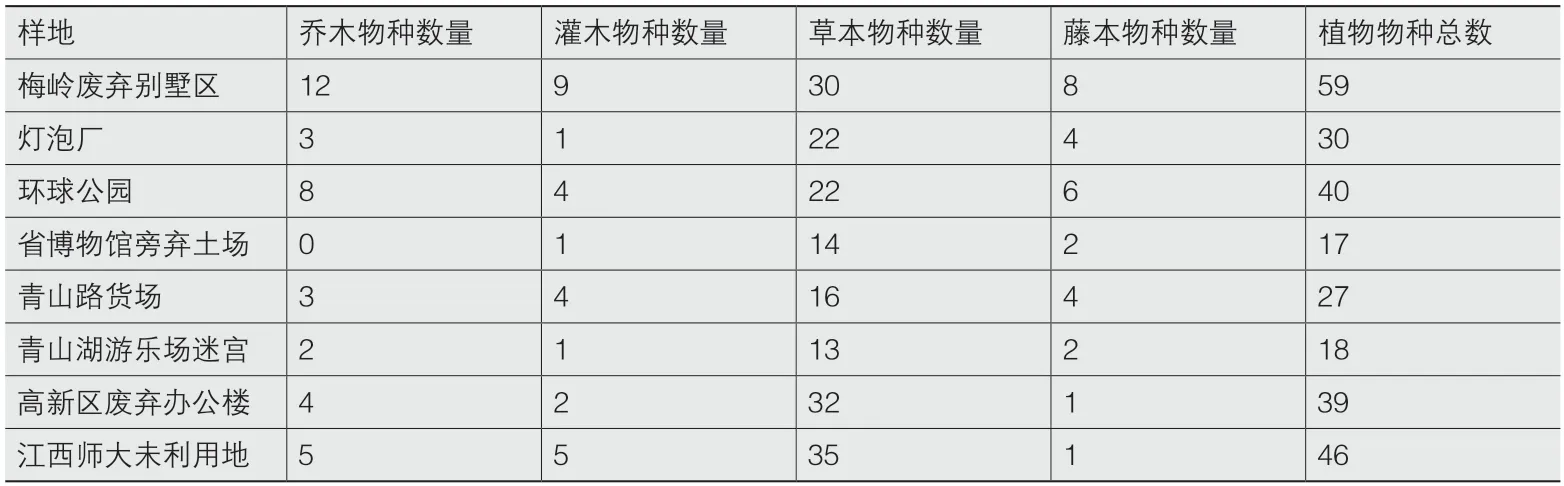
表3 不同样地植物物种数量(单位:个)
2.3 基于乔木恢复的演替特征
再野化过程是一个物种缓慢恢复的过程,但目前还未有研究深入探究其恢复演替机制。调查发现,灌木在同一再野化场地中分散分布,且植物形态差异大,灌龄难以测定;草本植物经长时间演替在较大空间尺度上整体分布趋于均质化(局部尺度由于物种繁衍方式存在优势物种斑块化生长现象),且多为一年生植物,其随时间梯度发展的特征较难量化分析,因此,本研究仅从乔木的种类变化特征角度探讨群落演替机制。基于不同场地的巨大差异,选择人为扰动小、有一定面积,且荒废之前为人为扰动后的裸地的梅岭废弃别墅区进行植被恢复演替特征研究。
2.3.1 乔木恢复的组成特征
通过实地调查,南昌市梅岭废弃别墅区自生乔木及乔木状植物共12种,包括白花泡桐、构树、乌桕、苦楝(Melia azedarach)、加杨(Populus×canadensis)、盐麸木(Rhus chinensis)、枫香(Liquidambar formosana)、朴树(Celtis sinensis)、香樟、女贞(Ligustrum lucidum)、马尾松(Pinus massoniana)和毛竹(Phyllostachys edulis)。由于毛竹的自然繁衍完全依靠场地内原有毛竹的分布(图3东北和东南2处原有林地均为香樟和毛竹林,其旁的密集常绿植物为毛竹),因此其分布特征在再野化过程中不存在典型性和代表性,故本文不对其做分析。对其余乔木进行分析可以发现以下特征。1)荒野样地再野化过程中,乔木的出现及发展由场地中的矗立物为核心,逐渐向外围扩散。从梅岭废弃别墅区再野化乔木核密度分析图(图3)及落叶与常绿乔木分布图(图4)可以看出,乔木的分布以别墅建筑为核心,建筑外墙周边乔木较为密集,并向外随距离增加而逐渐稀疏。这个现象在所有调查样地中均有显现,产生的原因可能是荒地乔木主要靠鸟类传播,场地中的突出矗立物是鸟类停息的立足点,因而矗立物周边就成了乔木恢复的起始点。在场地中没有矗立物的大块空地中(如博物馆旁弃土场、江西师大未利用地等)极少发现有乔木树种,也从另一方面证明了这个推断。2)再野化乔木树种以地带性的落叶先锋树种为主。梅岭废弃别墅区及其他样地乔木调查表明,裸地再野化过程中,自生乔木树种主要为构树、白花泡桐、乌桕、苦楝、枫香、盐麸木和朴树等落叶树种,其中占绝对优势的是构树和白花泡桐,且在除省博物馆旁弃土场样地外的其他样地均有这2种树种。3)在再野化过程中,落叶乔木和常绿乔木的冠幅及二者到建筑的距离存在较大差异。通过测量场地中所有乔木的冠幅及其到建筑外墙的邻近距离,并进行单因素方差分析(表4),结果表明:落叶与常绿乔木间冠幅的大小,以及与建筑的距离均存在极显著差异,箱线图(图5)说明场地中落叶乔木的冠幅比常绿乔木的冠幅大,也说明落叶乔木在植被恢复过程中先于常绿乔木生长于场地中,且落叶乔木离场地建筑物比常绿乔木近。乔木冠幅与建筑距离不存在显著相关性,原因可能是:虽然乔木会由于鸟类传播的原因而从建筑附近开始繁殖生长,但随着鸟类传播的持续进行,以及乔木自身的开花结果并自播繁殖,会使大龄树与小龄树交错存在。此外,邻近建筑的树木树冠也在一定程度上受到建筑外立面的限制与影响[25],而空旷地没有这种限制,所以整体上未形成明显的梯度特征。

表4 常绿乔木和落叶乔木再野化过程演变特征差异单因素方差分析
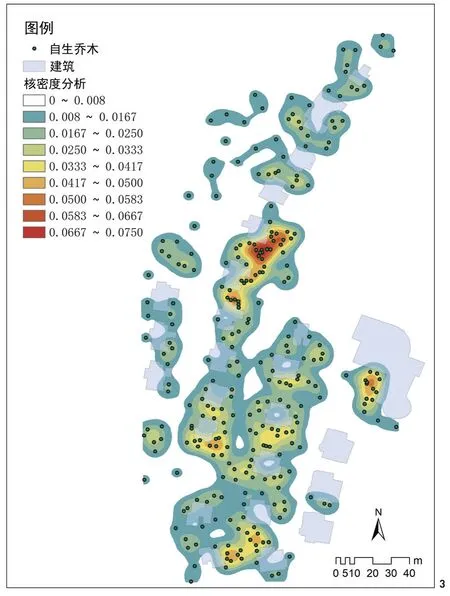
图3 梅岭废弃别墅区再野化乔木核密度分析图

图4 梅岭废弃别墅区再野化落叶与常绿乔木分布图
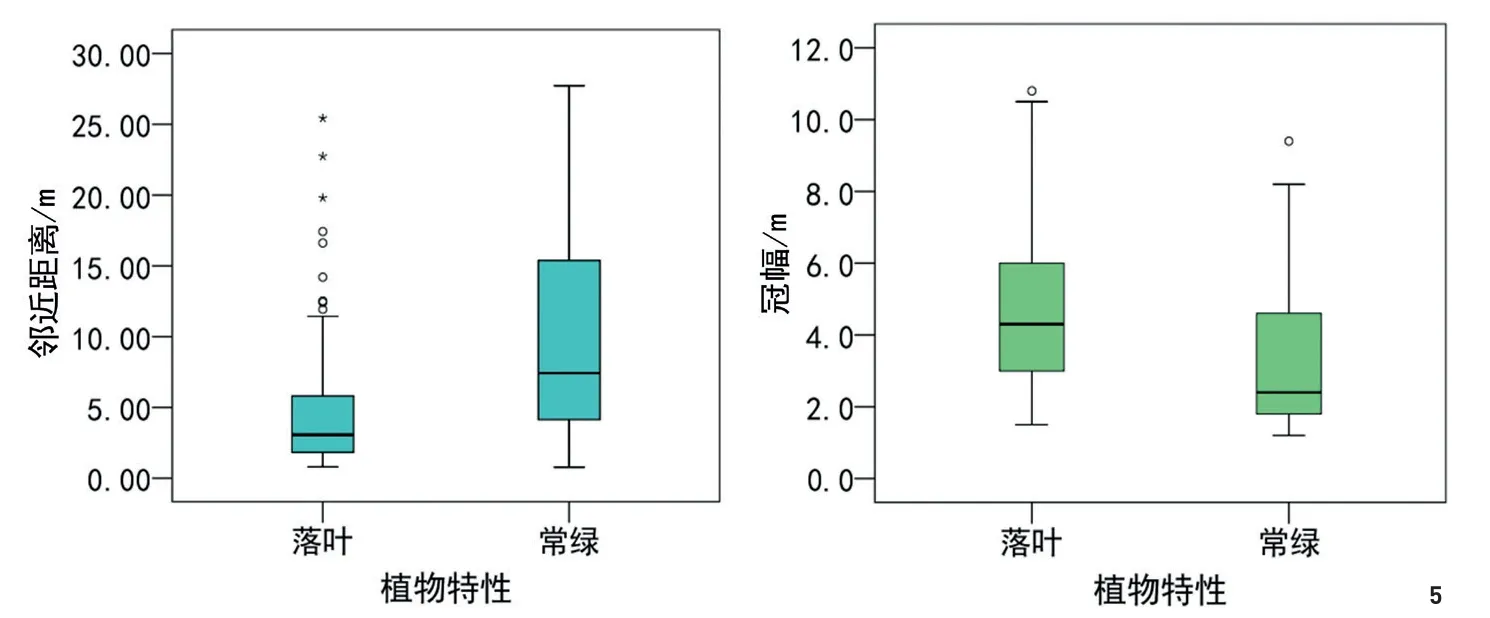
图5 常绿乔木和落叶乔木间再野化过程演变特征
2.3.2 不同乔木树种的再野化特征
在生态过程中,乔木树种的空间分布通常在较大尺度上表现出一定的趋势性,在中等或局部尺度上呈斑块分布,在小尺度上则表现为随机性。但在某些特定场地中,不同种类的植物也会反映相似的生态过程特征[26]。因此,本研究仅将场地中数量达到10株以上的树种分别进行空间分布核密度分析(图6),结果表明:1)各树种在种内均表现出一定的聚集特征,株数相对少的树种的斑块性聚集更加明显,表明再野化过程中,偶发性的母树对树种分布有重要影响;2)场地中存在明显的优势树种,但不同树种间存在空间占位现象,如图6所示,一种树种比较密集的区域,其他树种相对较少,说明在再野化早期,不同树种间竞争较小,自体传播方式容易形成同一树种在局部聚集生长。
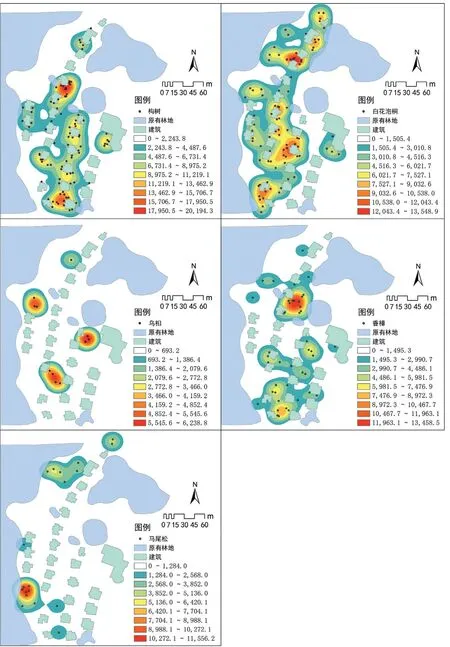
图6 梅岭废弃别墅区不同乔木树种再野化特征
对不同树种与场地矗立物(建筑)的关系进一步量化分析(图6、7),并结合图4,可以发现在与建筑的邻近距离上:构树>白花泡桐>香樟>乌桕>马尾松。这种分布现象体现了树种本身的生态特性差异。1)构树浆果是鸟类非常喜爱的食物,有助于构树的传播,且构树是一种耐瘠薄干旱的广适性植物,适应能力极强[27]。调查发现,构树在建筑屋顶、窗台、墙脚都有生长,也证明其传播方式及适应能力使其具有占据建筑最邻近空间的条件和能力。2)白花泡桐是喜阳植物,适应能力强,但更偏好土层深厚、湿润肥沃,pH值6~8的沙壤土,对于建筑附近残留或填埋的建筑废弃物及所形成的碱性环境的适应能力相比构树差[28],在调查中也极少发现紧贴建筑生长的白花泡桐。3)香樟适宜生长于pH值5.5~7.0的沙质壤土,忌碱性、瘠薄土壤,因而对建筑邻近空间的适应能力也较差[29]。4)乌桕喜光,适合生长于阳坡、旷地和林缘,阴坡、林内较少,特别是林地郁闭度达到0.8以上时,基本没有乌桕生长[30]。5)马尾松具有喜光、耐干旱、耐贫瘠的生长特性,在遮阴环境中其适应能力较差[31]。研究场地中的马尾松主要集中分布在原有林地下坡方向的空旷地中,表明是其松果从原有林地母树滚落到别墅区萌发生长,以及后期成熟马尾松自播繁殖的结果。
3 讨论
3.1 城市废弃地再野化时空维度的差异与协同
在空间上,城乡梯度空间的城市生态系统研究具有重要意义,相关研究表明,生态系统结构、功能及物种多样性具有显著的城乡梯度关系[32-33],且城市野化的自生植物物种丰富度是斑块特性及城乡梯度相互作用的结果[34],这与本研究结果相一致。在时间梯度上,已有研究表明,在缓慢的生态恢复过程中,物种丰富度和物种多样性总体呈增长趋势[35-36],本研究支持该观点。相关研究表明,物种组成和多样性模式更受区域尺度中微环境变量(土壤、水分、光照等)的影响[37],本研究对裸地再野化乔木树种空间分布特征的分析也表明,场地中的立地条件和地物地貌,特别是矗立物会对乔木树种再野化过程产生显著影响。城乡梯度与物种总数显著相关,但与不同类型物种的相关性存在较大差异,这可能是受到了城乡梯度上物种种库差异的影响;时间与物种相关性的不显著,则说明了再野化演替进程的复杂性,因此需要关注更长时间尺度上的再野化恢复机制,例如从裸地到稳定生态群落形成的时间尺度。本研究虽然探讨了时空梯度与样地差异对再野化植物多样性的影响,但受限于样本差异与数量,未能量化出时间梯度、空间梯度和场地特征三者对再野化进程中物种丰富度的影响。
3.2 再野化物种丰富度特征
调查表明,南昌市废弃地再野化的主要物种为草本植物,共计38科67属76种,与同属废弃地的伊敏露天煤矿排土场(再野化25年,自生植物21科64种)再野化结果相近[36],而与北京废弃采石场(再野化15年,共出现植物37种)有较大差异[35],并低于南京市公园绿地(56科186属241种自生草本植物)及北京奥林匹克森林公园(28科79属102种)[38-39],说明废弃地相较于城市绿地自生植物少,植物恢复缓慢,且同为废弃地时,场地初始立地条件对后期植物恢复有重要影响。从物种来源看,废弃地草本植物中的非入侵植物有42种,占比57.9%,入侵植物32种,占比42.1%;从传播途径看,动物传播8种,占比10.5%,风力传播20种,占比26.3%,自体传播44种,占比57.9%,水力传播4种,占比5.3%,说明在废弃地中自体传播为物种的主要传播途径,这与相关研究认为城市生境中土壤种子库是城市地上自发植物的时空丰度和多样性的决定因素的结果相一致[40]。对研究数据的相关分析也表明,不同样地非入侵、入侵物种数量及不同传播途径植物种数与时空梯度间不存在显著相关关系,这也说明城市的复杂环境会导致物种来源的复杂化。基于易鉴别性原因,现有对城市自生植物的研究主要是针对草本植物和木本幼苗[41],而少有对已经进入生长期及成熟期树木的研究,因此以再野化树木为研究对象探讨再野化演替机制可以拓展对再野化的系统认知。裸地再野化乔、灌、草的自然恢复进程中,乔、灌的恢复难度要大于草本,因此适当的人工恢复措施对于废弃地的生态恢复具有重要意义。再野化是一个复杂、综合的地上地下生态过程,对再野化裸地植物、动物、微生物、环境因子等的协同演替过程与效应进行研究,能更好地探索再野化复杂生态过程的机制和机理[39]。从“3C”视角而言,城市绿地的斑块大小、分维数和连通性等对再野化有重要影响[17-18]。从城市中自生乔木植物传播路径来看,乔木主要靠自体传播和动物传播,其中动物传播为乔木传播的主要方式,而飞行动物具有更大的传播空间和距离,因此对城市物种丰富度保护具有重要意义。研究城市环境再野化实践中的物种丰富度恢复过程、机制和路径,测度物种丰富度并预估其峰值,可以更加科学合理地指导城市再野化实践。
3.3 城市语境下的再野化
城市形态由空间构造和基础设施背景组成,构成了城市的物质维度;城市环境由人及其活动组成,构成了城市的主体维度[42]。无论是城市的物质维度还是主体维度,人的主导作用毋庸置疑。因而,城市语境中的再野化必然是从减少或避免人为介入与干扰的自然自主恢复演替的进程[12-15]。但以人为核心的环境塑造,必然体现社会的普遍意识和认知的合理性与正当性,这又为人为介入与干扰提供了强大的社会人文基础,并表现为对自生植物的选择性清除及再野化环境的干预(改造或围封)[43-44]。因此,城市再野化研究与实践迫切需要从人文视角和生态视角引导公众认知,促进公众对再野化的认同与参与[45-46]。
4 结论
通过对南昌市具有时空差异的8处废弃场地的再野化研究,对南昌市废弃地再野化时空特征得出如下结论。
1)再野化存在显著的城乡空间梯度关系,但时间梯度关系不明显。废弃场地的再野化是场地内因与环境外因、空间横向与时间纵向上共同作用的过程与结果。内因的生态本底与立地条件构成了再野化的基础,外因的环境条件发挥了促进物种迁移渗透及生态系统复杂性的重要作用。内因与外因实质上构成了空间横向维度的再野化基底,在空间基底上叠加时间的自然选择与修正,推动了再野化的演替进程。本研究表明,时间梯度上植被的自然恢复是一个缓慢的生态过程,因而在原有生态系统被重度破坏的场所,适宜的人工生态修复是快速恢复和重建新生态系统的必要措施。
2)不同立地条件及场地内外环境对植被再野化演进特征有显著影响。再野化的生态过程是一个双向影响的演化系统,既有内部的物种演进及物质能量流动的内推影响,又受内部与外部相互渗透的交互影响。生态系统所包含的动物、植物和微生物及其所处环境是一个综合的复杂系统,因此对于再野化的研究需要更深入地探讨各生态要素间相互作用的生态逻辑和机理。
3)南昌市再野化的先锋树种主要是落叶树种构树和白花泡桐,乔木的恢复与场地内矗立物的分布有密切关系,其生态占位与其传播途径和生理习性相关。再野化植被未形成稳定的植物群落前,不同类型乔木间呈现斑块状聚集性恢复特征,但其生态占位与植被自身生理特性及场地环境特征相关,表现出相应的空间依赖关系。人工生态修复需要考虑所选植被的种间竞争关系及环境适应能力,以促进荒野地植物群落的快速恢复。
注:文中图片除注明外,均由作者绘制。
猜你喜欢
建筑与预算(2022年6期)2022-07-08
绿色中国(2019年16期)2019-11-26
中国林业产业(2019年3期)2019-03-25
生态文明新时代(2018年2期)2018-03-21
中国扶贫(2017年16期)2017-08-16
西南学林(2016年2期)2016-11-08
建筑与预算(2014年5期)2014-04-11
河南科技(2014年16期)2014-02-27
学与玩(2009年2期)2009-03-09
