
不同土壤和微量元素对车桑子幼苗生长的影响
2023-10-17 06:14王雪梅闫帮国王梓丞史亮涛刘刚才
热带亚热带植物学报 2023年5期
王雪梅, 闫帮国, 王梓丞, 史亮涛, 刘刚才
不同土壤和微量元素对车桑子幼苗生长的影响
王雪梅1,2, 闫帮国3, 王梓丞1, 史亮涛3, 刘刚才2*
(1. 绵阳师范学院资源环境工程学院, 四川 绵阳 621000; 2. 中国科学院成都山地灾害与环境研究所, 成都 610041; 3. 云南省农业科学院热区生态农业研究所, 云南 元谋 651300)
为了解微量元素对车桑子()生长的作用,研究添加微量元素(硼B、铁Fe、锰Mn、锌Zn)对车桑子生长和叶绿素荧光特性的影响。结果表明,除Mn外,B、Zn和Fe均对车桑子的生长和叶绿素荧光参数有显著促进作用(<0.05);且添加B的车桑子具有更高的生物量积累,比对照显著提高了133.61%。微量元素与土壤类型对叶片磷(P)含量和叶片氮磷比(N/P)具有显著的交互作用(<0.05),紫色土添加Zn、黄棕壤添加Fe均显著降低了叶片N/P。燥红土和黄棕壤上车桑子的株高、叶面积和生物量积累均高于紫色土,但紫色土和黄棕壤上车桑子的根冠比和叶片N/P显著高于燥红土(<0.001)。这表明微量元素对干热河谷车桑子生长具有重要作用,在植被恢复过程中可通过添加B、Fe、Zn尤其是B来促进植物生长。
土壤类型;微量元素;车桑子;干热河谷;生物量
车桑子()又称为明油子、坡柳,是我国西南干热河谷区的优势灌木,优势度达到33.67%[1]。干热河谷生态环境具有水分与养分双重胁迫的特点,植被恢复一直是该区生态和经济建设的重点和难点工作[2–3]。车桑子作为一种乡土种,经过了长时期的进化和适应,由于其耐干旱、耐贫瘠、适应性强等特性,被广泛应用于干热河谷植被恢复中[4]。目前,对车桑子的研究主要集中于种子休眠[5–6]、药用价值[7]、对重金属污染地区的修复效应[8]、抗旱[9–10]、耐贫瘠[11–12]等方面。幼苗阶段是木本植物定植的瓶颈期,也是对环境变化较为敏感的阶段[13],环境因子对车桑子幼苗期的过滤效应十分显著,在植被恢复过程中应加强车桑子苗期管护[4]。
微量元素是植物生长过程中必不可少的营养元素,微量元素的缺乏会产生相应的缺素症,并严重影响植物生长[14–15]。目前公认的植物必需微量元素主要有铁(Fe)、锰(Mn)、锌(Zn)、铜(Cu)、硼(B)、钼(Mo)和氯(Cl)等。微量元素添加在农业和林业生产上的研究较多,土壤有机质含量及速效氮、速效磷和速效钾含量对少刺苏铁()的生长影响不明显,而土壤中微量元素Fe、Zn和 Mn的含量与其生长有密切关系[16]。添加Zn、B、Mn对兰州百合(var.)[17]、玉米()[18]、秃杉()[19]等均具有显著促进作用。施微量元素后,高寒沼泽湿地的植被盖度、地上生物量及主要物种的株高也明显提高[20]。早在1923年就证明B是植物必需的营养元素[21],虽然目前对此有所争议,认为B的作用可能表现为一种间接作用[22],但大量研究表明[23–26], B对维持细胞质膜的完整性、叶片光合作用、植物抗逆性等都具有重要作用,而缺B会影响根伸长、吲哚乙酸氧化酶活性、糖转运、碳水化合物代谢、核酸合成和花粉管生长等,从而影响植物生长发育。此外,从干热河谷植物叶片中各生源要素的异速增长关系来看,Fe的增长速率大于N、P、K[27],表明Fe在干热河谷植物生长过程中极为重要。干旱是干热河谷植物生长面临的最主要的环境问题。Zn能提高植物的抗旱能力,而缺Zn会降低水分用于生物量生产的效率,并且会降低植物应对水分胁迫的渗透调节能力[28–29]。闫帮国等[30–31]报道元谋燥红土的有效Fe、有效Zn和有效Mn的含量分别为0.49、0.20和7.58 mg/kg,说明元谋燥红土缺Fe、缺Zn,但不缺Mn。Mn对凋落物的周转具有重要作用,在干热河谷生态系统功能中同样具有不容忽视的地位[32]。由于不同土壤会影响植物对微量元素的吸收[33], 微量元素缺乏会导致叶片皱缩[31]。生长于黄棕壤和紫色土的车桑子出现叶片皱缩、卷曲现象, 可能是不同土壤中的微量元素差异导致的。然而, 对西南干热河谷区土壤微量元素与植物生长关系的研究还鲜有报道,不同土壤上添加微量元素对车桑子生长效应还不清楚。
因此,本研究以车桑子幼苗为研究对象,从云南元谋县采集不同的土壤进行4种微量元素(B、Fe、Mn、Zn)添加试验,研究其对车桑子幼苗生长与生物量积累、氮磷养分吸收和叶绿素荧光特性的影响,探究车桑子对不同微量元素的响应,并探明不同土壤上车桑子的生长差异和微量元素的限制作用,为车桑子在干热河谷植被恢复中的应用提供理论参考。
1 材料和方法
1.1 研究区概况
试验在中国科学院成都山地灾害与环境研究所元谋干热河谷沟蚀崩塌观测研究站进行,观测站位于云南省楚雄彝族自治州元谋县(25°23′~26°06′ N,101°35′~102°06′ E)。元谋县位于云贵高原北缘的金沙江一级支流龙川江下游的河谷地带,海拔898~2 836 m,相对高差约2 000 m,生态环境垂直分异明显,从河谷到山顶依次为河谷南亚热带(898~ 1 350 m)、山地中亚热带(1 351~1 700 m)、山地北亚热带(1 701~2 000 m)及山地温暖带-中温带(2 001~2 835 m)等4个垂直气候带[34]。河谷区是典型的干热河谷生态脆弱带,区域内年均温21.5 ℃,年蒸发量大于3 500 mm,年均降雨量小于700 mm。其中6月— 10月为雨季,期间降雨量占全年的90%以上,而干季较长,达6~7个月(11月至翌年5月)。低海拔区的植被类型主要为车桑子、扭黄茅()灌草群落,中海拔区主要为栎树()、车桑子、栌菊木()、云南松()等乔灌植物,高海拔区主要为杜鹃()、云南松、栎树等乔木群落。从低海拔到高海拔区土壤类型分别为燥红土、紫色土和黄棕壤,其中,燥红土和紫色土区都有车桑子生长,而山顶黄棕壤区没有车桑子生长[11–12]。
1.2 材料
在元谋干热河谷车桑子种子成熟季节,选择性状优良的母树采集成熟饱满、大小基本一致的种子装于布袋,带回实验室自然风干保存,种子百粒重为(1.327±0.032) g。采样点位于元谋坝区苴林乡, 受人为干扰较小,植被类型主要为车桑子灌木林,还有扭黄茅、孔颖草()、柱花草()等当地优势草本植物,土壤类型为燥红土。
1.3 方法
本研究在元谋县典型土壤上进行微量元素添加处理,土壤从不同的海拔梯度进行采集。虽然高海拔区黄棕壤上没有车桑子分布,但在全球气候变暖的背景下,植物有往更高处迁移的趋势[35]。因此,为探讨气候变化条件下车桑子对潜在土壤环境的适应性,采集了山顶黄棕壤。试验所用的土壤类型包括河谷区燥红土、中海拔区紫色土和高海拔区黄棕壤。除全P和有效P含量外,3种土壤类型的其余理化指标均具有显著差异,土壤有机质、全N、有效P含量为黄棕壤>紫色土>燥红土[11–12]。
在采样点采集0~20 cm的表层土壤,每种土壤类型采集3个土样,将土壤运回观测站,经风干、挑拣、碾碎、过筛后混匀装入盆钵,每个盆钵大约装土1 400 mL。所有盆钵放入25 ℃/35 ℃ (8 h/16 h)的气候培养箱中,一次性浇透水后播种经过休眠破除的车桑子种子。待种子发芽1个月,长出2~3片真叶后定植,5株/钵,开始进行微量元素添加处理。微量元素处理设置对照(CK)、加硼(+B)、加铁(+Fe)、加锰(+Mn)、加锌(+Zn)共5种,3种土壤类型共15个处理,每个处理4个重复。微量元素以水溶液进行添加,参照霍格兰营养液成分[36],分别以H3BO3、FeSO4·7H2O、MnCl2·4H2O和ZnSO4·7H2O形式添加。为保证车桑子对微量元素的有效吸收,微量元素营养液每周添加1次,每盆添加50 mL,对照则添加等量的蒸馏水。其余时间每天添加等量的蒸馏水,以保证植株正常生长。试验处理3个月后,收获车桑子样品进行相关指标的测定。
1.4 指标测定
试验末期,使用叶绿素荧光仪Handy PEA 1024测定车桑子叶片的叶绿素荧光特征。每盆随机选择植株中部、充分展开的叶片,暗适应30 min后进行叶绿素荧光参数的测定,获得最大荧光(Fm)、最小荧光(Fo)、可变荧光(Fv)、PS II的最大光化学效率(Fv/Fm)和潜在活性(Fv/Fo)等参数[37]。用直尺测量植株的株高;每盆选取植株中部、充分展开的5片叶片用扫描仪扫描,用Image J数字化计算叶面积。
试验结束时,将植株的地上和地下部分分开,冲洗干净后放入65 ℃烘箱中烘干,用电子天平称量各部分质量,然后将叶片粉碎后,测定N、P含量。全N含量采用凯氏定氮法测定,全P含量采用钼锑抗比色法测定[38]。
1.5 数据分析
根冠比=根生物量/地上生物量。用双因素方差分析法(Two-Way ANOVA)分析土壤类型和微量元素添加对车桑子形态(株高、叶面积)、生物量(根生物量、地上生物量、根冠比)、叶片养分和叶绿素荧光特征(Fo、Fv/Fo、Fv/Fm)的影响,并用最小显著性差异法(least significant difference, LSD)进行多重比较。同时,对这些指标进行主成分分析。此外,将数据进行对数转化处理,采用响应指数反映各指标对微量元素添加的响应,响应指数(response ratio)= ln[ADD/CK],式中,ADD为养分添加处理(B、Fe、Mn、Zn)响应变量的测量值,CK为对照处理响应变量的测量值。响应指数小于0,表示养分添加具有负效应,大于0表示具有正效应[39–40],用Student’s检验处理的响应指数与0的差异显著性。所有数据处理在SPSS 19.0中进行。
2 结果和分析
2.1 土壤类型和微量元素对形态特征的影响
土壤类型和添加微量元素均对车桑子株高和叶面积具有显著影响,但两者的交互作用不显著(表1)。车桑子在燥红土和黄棕壤上的株高分别比紫色土上的显著提高了24.04%和21.89%;燥红土和黄棕壤上的叶面积也分别比紫色土显著增加了24.22%和19.17%;而燥红土和黄棕壤间的株高和叶面积无显著差异。从图1可见,+B和+Fe的株高分别比对照显著增加了12.65%和12.30%;不同微量元素处理间的叶面积也具有极显著差异(<0.001),其中,+B、+Zn和+Fe的叶面积显著增大,分别比对照增加81.82%、56.84%和47.49%。响应指数的变化趋势一致(表2)。
2.2 土壤类型和微量元素对生物量的影响
土壤类型和添加微量元素显著影响了车桑子的生物量积累,但两者不存在显著的交互作用(表1)。黄棕壤的根生物量显著高于燥红土和紫色土,同时,黄棕壤和燥红土的地上生物量也显著高于紫色土,黄棕壤和燥红土的车桑子总生物量分别比紫色土显著提高了39.03%和34.59%。此外,不同土壤类型的车桑子根冠比具有显著差异(<0.001),燥红土的根冠比显著低于紫色土和黄棕壤,紫色土和黄棕壤无显著差异。与对照相比,+B、+Zn和+Fe显著促进了车桑子的生物量积累,其总生物量分别比对照显著提高了133.61%、65.31%和59.33%,但添加微量元素没有改变车桑子的根冠比(图2)。响应指数表明+Mn显著降低了车桑子根生物量(表2)。

表1 土壤类型和微量元素对车桑子生长特性的双因素方差分析

图1 不同土壤添加微量元素对车桑子形态特征的影响。CK: 对照。柱子上不同字母表示差异显著(P<0.05)。下同
2.3 土壤类型和微量元素对N、P养分的影响
从图3可见,叶片的N和P含量均以燥红土>紫色土>黄棕壤,且差异达显著水平;而叶片N/P以黄棕壤>紫色土>燥红土,黄棕壤和紫色土均大于20,显著高于燥红土。此外,微量元素与土壤类型对叶片P含量和N/P有显著的交互作用(表1)。燥红土上,+Mn显著增加了叶片P含量,+Zn显著降低了叶片P含量;而紫色土上,+Zn显著增加了叶片P含量,+Zn和+Fe均显著降低了叶片N/P;黄棕壤上,+Fe显著降低了叶片N/P。
2.4 土壤类型和微量元素对叶绿素荧光特性的影响
从图4、表1和2可见,土壤类型和添加微量元素显著影响了车桑子叶片的叶绿素荧光特征。紫色土的Fo最低,而Fv/Fo和Fv/Fm最高,均与黄棕壤和燥红土的差异达显著水平。+B和+Zn处理的Fv/Fo分别比对照显著增加了9.97%和9.62%,且Fv/Fm也显著增加。添加微量元素处理的车桑子叶片Fv/Fm为0.786~0.836,且Fv/Fo和Fv/Fm有极显著的正相关关系(=0.996,<0.001)。
2.5 主成分分析
主成分分析表明(表3),前3个主成分特征值均大于1,累积贡献率达87.136%,其中,第1主成分与Fv/Fm、Fv/Fo、Fo、P含量紧密相关,以Fv/Fm、Fv/Fo、Fo的载荷量最大,表现为叶绿素荧光参数;第2主成分与N/P、P和N含量紧密相关,表现为N、P养分特征;第3主成分与株高、总生物量、叶面积紧密相关,反映幼苗的生长特征,生物量积累与株高(=0.474,<0.01)、叶面积(=0.915,<0.001)呈显著正相关。

表2 添加不同微量元素处理的响应指数
*:<0.05; **:<0.01
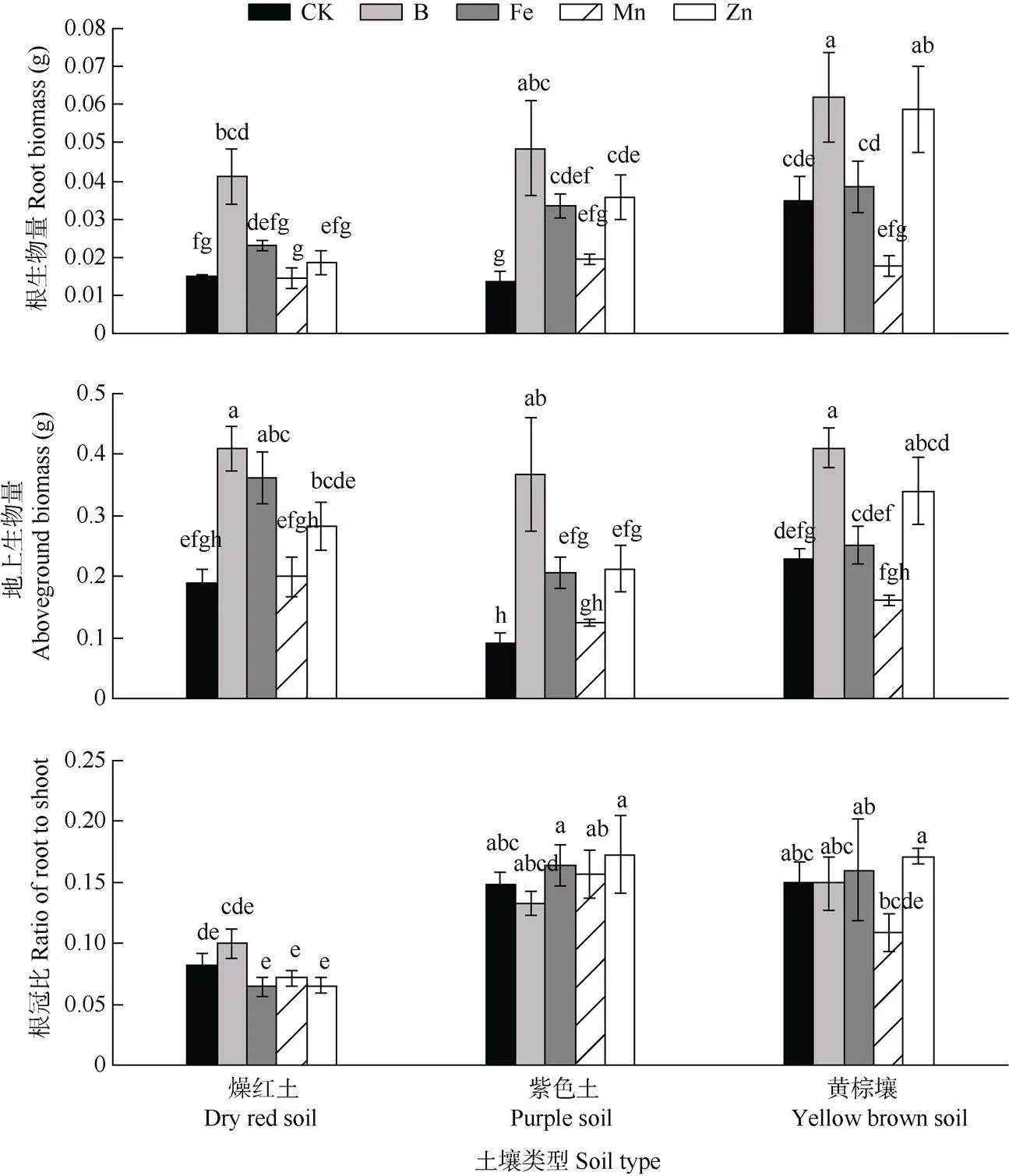
图2 不同土壤上添加微量元素对车桑子生物量特征的影响
3 结论和讨论
3.1 微量元素对生长和叶绿素荧光特性的影响
本研究结果表明,不同土壤上+B、+Zn和+Fe 对车桑子幼苗生长(株高、叶面积和生物量)有促进作用,尤以+B更为突出,说明车桑子对B的需求较大。前人[14,41]研究表明,缺B使甘薯()和柑桔生物量显著降低,严重限制植物的生长。添加不同微量元素均可促进车桑子的生长, 表明植物的微量元素限制作用不符合利比希最小因子定律。养分资源的共同限制是生物量限制的普遍形式,并可分为同时发生的共同限制、独立的共同限制和序列限制[39]。本研究中多种微量元素对车桑子幼苗可能表现为独立的共同限制,即单独添加各种微量元素时都有显著的促进作用。
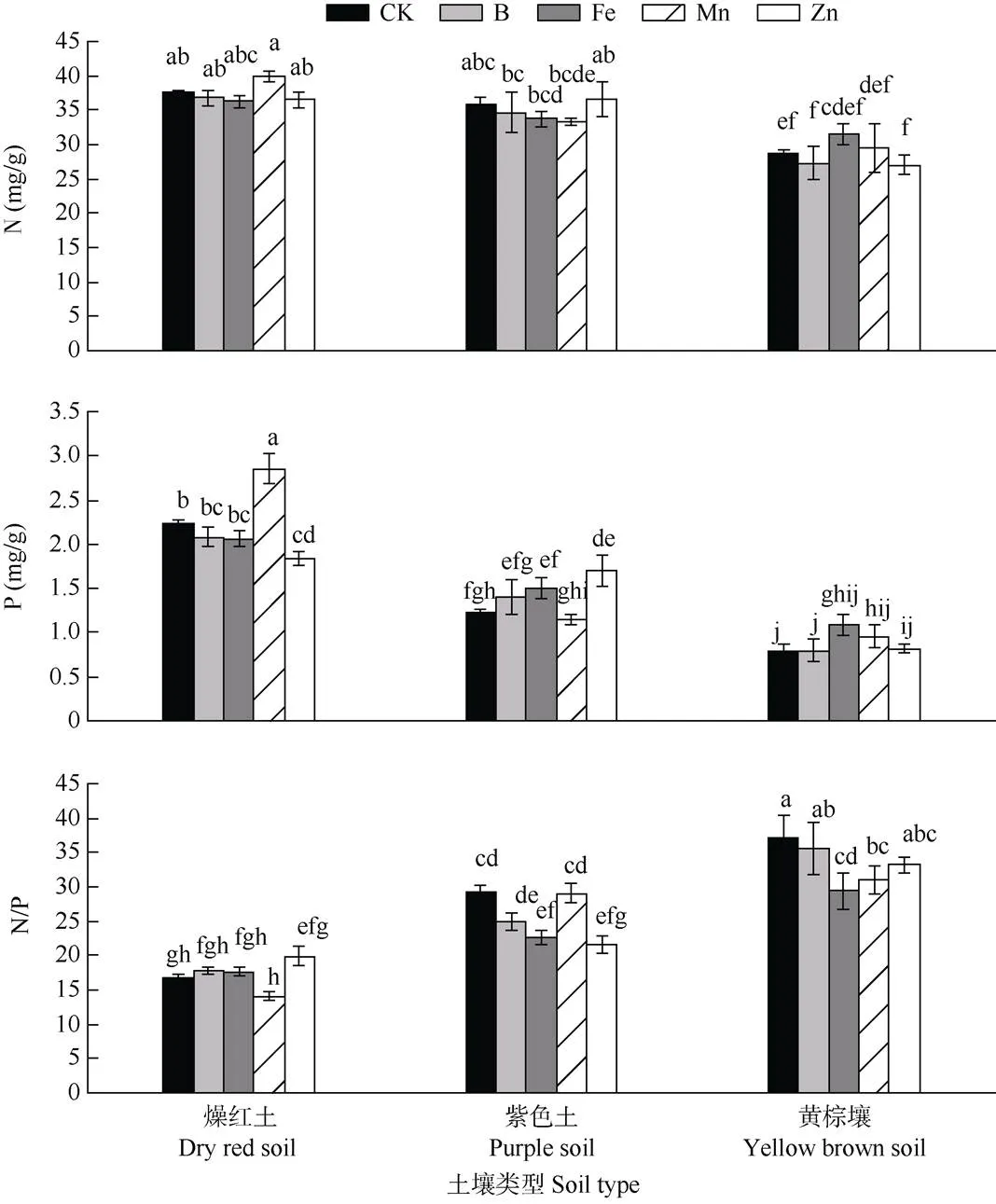
图3 不同土壤上添加微量元素对车桑子叶片N、P养分的影响

图4 不同土壤上添加微量元素对车桑子叶绿素荧光特征的影响
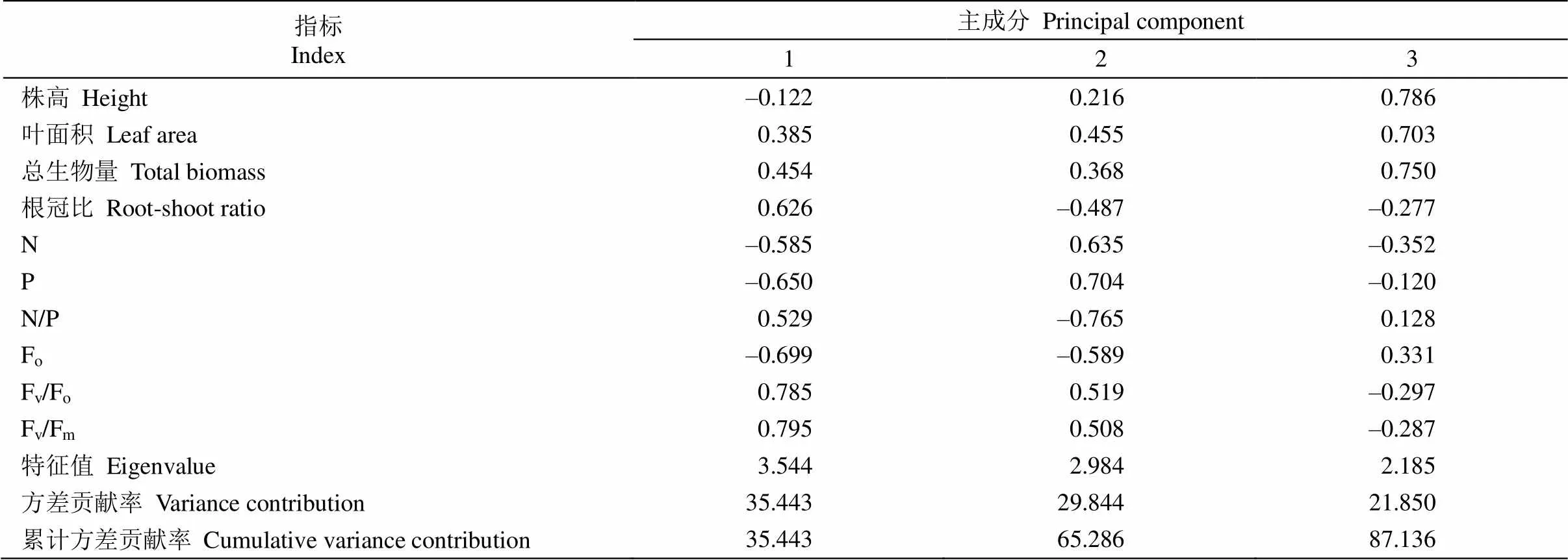
表3 各主成分特征向量、特征值及贡献率
叶绿素荧光参数也可作为诊断植物微量元素营养状况的指标之一[42]。本研究中B和Zn能提高PSII的Fv/Fo和Fv/Fm。Fv/Fm是叶绿体PSII光化学效率的一种度量,一般为0.80~0.85,在胁迫条件下会显著降低[43]。本研究中添加微量元素处理的Fv/Fm基本都大于0.80,说明车桑子幼苗没有受到明显的胁迫作用,没有发生光抑制,一定程度上说明车桑子的适应性较广[44]。植物的Fv/Fm愈高,表明其对光能的转化利用率和对环境的适应能力愈强[42],因此,微量元素B和Zn的添加提高了车桑子幼苗对光能的吸收利用效率。从施肥的角度来说,当一种生命元素以生物有效形态添加时,如果造成了生物量的增加或生化过程速率的增加,那么就存在着这种养分的限制[45]。综合来看,微量元素B、Zn和Fe的添加对车桑子幼苗生长有显著的促进作用,但添加Mn未表现出显著的促进作用。因此,土壤中的有效微量元素B、Zn和Fe对车桑子幼苗生长存在限制,而Mn元素不存在限制作用。前期研究表明[30–31], 元谋燥红土的有效Fe和有效Zn含量均较为缺乏,而有效Mn含量相对较高,这和本研究结果一致。虽然Mn元素对干热河谷凋落物分解具有重要作用[32],但对车桑子幼苗的生长和叶绿素荧光特性并没有促进作用。
3.2 微量元素和土壤类型对叶片氮磷养分的交互作用
不同土壤上车桑子叶片P含量和N/P对微量元素添加的响应不同,微量元素对植物叶片N、P含量的影响表明元素之间存在复杂的相互作用。矿质元素N、P对植物的生长发育和产量起着重要的作用,是植物生长过程中最关键的两种营养元素,微量元素的缺乏会导致植物对N、P吸收的减少[38]。燥红土上+Zn显著降低叶片P含量,可能一方面是微量元素的添加减少了对P的吸收,另一方面与叶片增长对养分浓度的稀释作用有关。然而,燥红土上+Mn显著增加了叶片P含量,这可能与植物体内的生态化学计量平衡有关,或者与Mn对土壤养分循环的影响有关。Mn是多种土壤氧化酶的重要辅助因子[46],土壤中Mn含量增加可促进有机质的分解,释放N、P养分,从而促进植物N、P养分的增加。紫色土+Zn不仅显著促进了车桑子叶面积和地上生物量的增加,也显著增加叶片P含量,降低叶片N/P,表明紫色土+Zn促进车桑子对土壤中P的吸收。同样,黄棕壤+Fe促进了叶片对P的吸收,显著降低叶片N/P,说明紫色土和黄棕壤上车桑子的生长存在磷限制[11–12],表明某些微量元素添加在一定程度上可提高车桑子对限制性养分的吸收。
3.3 土壤类型对车桑子生长的影响
从形态和生物量特征来看,3种土壤类型中以燥红土和黄棕壤的车桑子生长最好,紫色土较差。紫色土和黄棕壤上车桑子具有更高的根冠比,表明车桑子生长存在养分限制,需要增加根系生长以促进对土壤养分的吸收[47]。当叶片N/P大于20时, 表明存在磷的限制[48],紫色土和黄棕壤上叶片N/P显著高于燥红土,说明微量元素的添加并没有整体上解除紫色土和黄棕壤上的磷限制。虽然黄棕壤上土壤养分状况尤其N显著高于紫色土,但黄棕壤上叶片的N、P含量均显著低于紫色土,N/P更大,磷限制作用更强。然而,黄棕壤上车桑子生物量又高于紫色土,说明土壤N、P养分绝对含量并不是车桑子生长的决定因素,相比于N、P,微量元素对紫色土上植物生长的限制作用可能更大,这种限制作用导致了紫色土上植物吸收较高的N、P养分后也无法正常生长,导致生物量较低和叶片N、P含量较高,紫色土上车桑子生长对微量元素添加的响应指数最大,而黄棕壤较小,即黄棕壤上微量元素的限制作用较小。另外,本试验的土壤从野外采集回来后,在观测站堆放了半年才开始进行试验,黄棕壤中带有的乡土微生物对车桑子生长也具有显著的促进作用[49],从而使黄棕壤上车桑子的生长优于紫色土。
综上,车桑子在3种土壤类型中以燥红土上的生长较好,表明在干热河谷燥红土区用车桑子进行植被恢复是比较合适的。同时,添加微量元素对干热河谷车桑子幼苗生长有重要作用,在恢复过程中通过添加微量元素B、Fe、Zn来促进车桑子幼苗生长是值得尝试的措施,这对于干热河谷植被恢复工作具有重要的借鉴意义。然而,本研究没有测定土壤和植株的微量元素含量,这导致了一些适应机理无法解释清楚,如微量元素添加如何影响N、P养分的吸收,是否与植物体内生命元素的化学计量平衡有关?各微量元素在植物抗旱性方面具有什么作用?还有待于进一步研究和探讨。
[1] YANG Z Y. Research on landscape dynamics and restoration of vegetation in Hot-and-dry Valley of Yuanmou County, Yunnan Province [D]. Beijing: Chinese Academy of Forestry, 2007. [杨振寅. 元谋干热河谷植被景观动态与植被恢复研究 [D]. 北京: 中国林业科学研究院, 2007.]
[2] YANG Z Y, SU J R, LUO D, et al. Progress and perspectives on vegetation restoration in the Dry-hot Valley [J]. For Res, 2007, 20(4): 563–568. [杨振寅, 苏建荣, 罗栋, 等. 干热河谷植被恢复研究进展与展望 [J]. 林业科学研究, 2007, 20(4): 563–568. doi: 10.3321/j.issn: 1001-1498.2007.04.024.]
[3] ZHANG J Y, XU Y, SU C J, et al. Research progress on vegetation restoration in the Dry-hot Valleys of the Jinsha River [J]. Res Soil Water Conserv, 2005, 12(6): 101–104. [张金盈, 徐云, 苏春江, 等. 金沙江干热河谷植被恢复研究进展 [J]. 水土保持研究, 2005, 12(6): 101– 104.
[4] ZHANG Q, LIU L W, LI J Q, et al. Characteristics ofpopulation regenerated in abandoned cropland of Yunnan Dry-hot Valleys [J]. SW China J Agric Sci, 2016, 29(9): 2234–2238. [张琴, 刘利文, 李俊清, 等. 云南干热河谷区裸地植被恢复过程中车桑子的种群特征 [J]. 西南农业学报, 2016, 29(9): 2234–2238. doi: 10.16213/j.cnki.scjas.2016.09.039.]
[5] BASKIN J M, DAVIS B H, BASKIN C C, et al. Physical dormancy in seeds of(Sapindales, Sapindaceae) from Hawaii [J]. Seed Sci Res, 2004, 14(1): 81–90. doi: 10.1079/SSR2003157.
[6] PHARTYAL S S, BASKIN J M, BASKIN C C, et al. Physical dormancy in seeds of(Sapindaceae) from India [J]. Seed Sci Res, 2005, 15(1): 59–61. doi: 10.1079/SSR2004194.
[7] RANI M S, PIPPALLA R S, MOHAN K.Linn.: An overview [J]. J Pharm Res Health Care, 2009, 1: 97–112.
[8] ROJAS-LORIA C C, FAVELA-TORRES E, GONZÁLEZ-MÁRQUEZ H, et al. Role of glutathione and glutathione-transferase in lead tole- rance and bioaccumulation by(L.) Jacq [J]. Acta Physiol Plant, 2014, 36(9): 2501–2510. doi: 10.1007/s11738-014-1623-8.
[9] CHEN S Y, LANG N J, JIA L Q, et al. Effects of drought stress on lipid peroxidation and activity of defense enzymes of,andseedlings [J]. Bull Bot Res, 2006, 26(1): 88–92. [陈少瑜, 郎南军, 贾利强, 等. 干旱胁迫对坡柳等抗旱树种幼苗膜脂过氧化及保护酶活性的影响 [J]. 植物研究, 2006, 26(1): 88–92. doi: 10.3969/j.issn.1673-5102.2006.01.019.]
[10] LI J Y, JIA L Q, LANG N J, et al. Characteristics of photosynthesis ofin the Dry-hot Valley of Jinsha River [J]. J Beijing For Univ, 2003, 25(5): 20–24. [李吉跃, 贾利强, 郎南军, 等. 金沙江干热河谷坡柳的光合特性 [J]. 北京林业大学学报, 2003, 25(5): 20–24. doi: 10.3321/j.issn:1000-1522.2003.05.005.]
[11] WANG X M, YAN B G, ZHAO G, et al. Responses ofgrowth and soil biological properties to nitrogen and pho- sphorus additions in Yuanmou Dry-hot Valley [J]. J Mount Sci, 2018, 15(6): 1283–1298. doi: 10.1007/s11629-017-4544-3.
[12] WANG X M, LIU Q, YAN B G, et al. Responses of photosynthetic characteristics ofto nitrogen and phosphorus addition in a Dry-hot Valley [J]. Acta Ecol Sin, 2019, 39(22): 8615– 8629. [王雪梅, 刘泉, 闫帮国, 等. 干热河谷车桑子光合生理特性对氮磷添加的响应 [J]. 生态学报, 2019, 39(22): 8615–8629. doi: 10. 5846/stxb201810102197.]
[13] LECK M A, PARKER V T, SIMPSON R L. Seedling Ecology and Evolution [M]. Cambridge: Cambridge University Press, 2008: 189–214.
[14] O’SULLIVAN J N, ERNEST J. Nutrient deficiencies in lesser yam () characterized using constant-water table sand culture [J]. J Plant Nutr Soil Sci, 2007, 170(2): 273–282. doi: 10.1002/ jpln.200625049.
[15] AFROUSHEH M, ARDALAN M, HOKMABADI H, et al. Nutrient deficiency disorders inseedling rootstock in relation to eco-physiological, biochemical characteristics and uptake pattern of nutrients [J]. Sci Hort, 2010, 124(2): 141–148. doi: 10.1016/j.scienta. 2009.12.001.
[16] LI N, CHEN K K, GU W L, et al. Effects of physical-chemical charac- ters and microelement contents in soil on growth of[J]. J Plant Resour Environ, 2006, 15(3): 43–46. [李楠, 陈考科, 顾蔚蓝, 等. 土壤理化性状及微量元素含量对少刺苏铁生长的影响 [J]. 植物资源与环境学报, 2006, 15(3): 43–46. doi: 10.3969/j.issn.1674- 7895.2006.03.010.]
[17] LU Z, HUANG P, WANG Y Z. Effects of spraying boron, zinc, manganese fertilizer on accumulation and distribution of dry matter, yield and N P asorption ofvar.[J]. Soil Fertil Sci China, 2011(1): 39–43. [路喆, 黄鹏, 王玉忠. 喷施锌、硼、锰肥对兰州百合干物质积累分配、产量和氮磷吸收的影响 [J]. 中国土壤与肥料, 2011(1): 39–43. doi: 10.3969/j.issn.1673-6257.2011.01.009.]
[18] ZHANG X Y. Effect of trace element on growth and development and mineral nutrients absorption of high-yielding summer maize (L.) [D]. Baoding: Hebei Agricultural University, 2019. [张晓园. 微量元素对夏玉米生长发育及矿质养分吸收的影响研究 [D]. 保定: 河北农业大学, 2019.]
[19] SUN Z G, LIU Y C, CHEN Q, et al. Effect of trace elements spraying onseedlings in Tengchong County [J]. J W China For Sci, 2016, 45(1): 137–141. [孙志刚, 刘云彩, 陈强, 等. 腾冲秃杉苗木喷施微量元素的效应 [J]. 西部林业科学, 2016, 45(1): 137–141. doi: 10.16473/j.cnki.xblykx1972.2016.01.023.]
[20] LUO Q Y, MA Y G, XIE H G, et al. Effect of micro-fertilizer appli- cation on the soil dynamic and the status of vegetation in Qilian Mountain alpine-cold swamp ecosystem [J]. J Qinghai Norm Univ (Nat Sci), 2015, 31(3): 27–33. [罗巧玉, 马永贵, 谢慧春, 等. 施微肥对祁连山高寒沼泽湿地生态系统土壤及植被生长状况的影响 [J]. 青海师范大学学报(自然科学版), 2015, 31(3): 27–33. doi: 10.3969/j. issn.1001-7542.2015.03.006.]
[21] WARINGTON K. The effect of boric acid and borax on the broad bean and certain other plants [J]. Ann Bot, 1923, 37(148): 629–672.
[22] MCGRATH S P. Arguments surrounding the essentiality of boron to vascular plants [J]. New Phytol, 2020, 226(5): 1225–1227. doi: 10. 1111/nph.16575.
[23] GOLDBACH H E, WIMMER M A. Boron in plants and animals: Is there a role beyond cell-wall structure? [J]. J Plant Nutr Soil Sci, 2007, 170(1): 39–48. doi: 10.1002/jpln.200625161.
[24] LI Z L, ZHANG J L, LI Z G, et al. The physiological mechanism of the regulation of boron on the growth and development and resistance of potato plantlets under drought stress [J]. Agric Res Arid Areas, 2014, 32(5): 67–72. [李自龙, 张俊莲, 李宗国, 等. 硼调控干旱胁迫下马铃薯生长发育及抗性的生理机制 [J]. 干旱地区农业研究, 2014, 32 (5): 67–72.]
[25] CAKMAK I, RÖMHELD V. Boron deficiency-induced impairments of cellular functions in plants [J]. Plant Soil, 1997, 193: 71–83. doi: 10. 1023/A:1004259808322.
[26] CAMACHO-CRISTÓBAL J J, REXACH J, GONZÁLEZ-FONTES A. Boron in plants: Deficiency and toxicity [J]. J Integr Plant Biol, 2008, 50(10): 1247–1255. doi: 10.1111/j.1744-7909.2008.00742.x.
[27] YAN B G, HE G X, LI J C, et al. Scaling relationships and stoichio- metry of plant leaf biogenic elements from the arid-hot valley of Jinsha River, China [J]. Chin J Plant Ecol, 2012, 36(11): 1136–1144. [闫帮国, 何光熊, 李纪潮, 等. 金沙江干热河谷地区植物叶片中各生源要素的化学计量特征以及异速增长关系 [J]. 植物生态学报, 2012, 36 (11): 1136–1144. doi: 10.3724/SP.J.1258.2012.01136.]
[28] KHAN H R, MCDONALD G K, RENGEL Z. Zinc fertilization and water stress affects plant water relations, stomatal conductance and osmotic adjustment in chickpea (L.) [J]. Plant Soil, 2004, 267(1/2): 271–284. doi: 10.1007/s11104-005-0120-7.
[29] MOTALEBIFARD R, NAJAFI N, OUSTAN S, et al. The combined effects of phosphorus and zinc on evapotranspiration, leaf water poten- tial, water use efficiency and tuber attributes of potato under water deficit conditions [J]. Sci Hort, 2013, 162: 31–38. doi: 10.1016/j.scienta. 2013.07.043
[30] YAN B G, HE G X, SHI L T, et al. Element concentration in leaves and nutrient resorption efficiency on dry-red soil and vertisols in Dry and Hot Valley in Yuanmou, China [J]. Chin J Appl Ecol, 2016, 27(4): 1039–1045. [闫帮国, 何光熊, 史亮涛, 等. 元谋干热河谷燥红土和变性土上植物叶片的元素含量及其重吸收效率 [J]. 应用生态学报, 2016, 27(4): 1039–1045. doi: 10.13287/j.1001-9332.201604.002.]
[31] SHEN H G. Influence of soil microelement on plant [J]. For Sci Technol Inform, 2010, 42(4): 12–14. [沈惠国. 土壤微量元素对植物的影响 [J]. 林业科技情报, 2010, 42(4): 12–14. doi: 10.3969/j.issn. 1009-3303.2010.04.006.]
[32] YAN B G, JI Z H, HE G X, et al. The effects of leaf stoichiochemistric characters on litter turnover in an Arid-hot Valley of Jinsha River, China [J]. Acta Ecol Sin, 2013, 33(18): 5668–5674. [闫帮国, 纪中华, 何光熊, 等. 金沙江干热河谷植物叶片元素含量在地表凋落物周转中的作用 [J]. 生态学报, 2013, 33(18): 5668–5674. doi: 10.5846/stxb 201304180739.]
[33] LU L, JI Y M, LI L R, et al. Analysis of Fe, Zn and Se contents in different wheat cultivars (lines) planted in different areas [J]. Chin J Appl Environ Biol, 2010, 16(5): 646–649. [鲁璐, 季英苗, 李莉蓉, 等. 不同地区、不同品种(系)小麦锌、铁和硒含量分析 [J]. 应用与环境生物学报, 2010, 16(5): 646–649. doi: 10.3724/SP.J.1145.2010.00646.]
[34] The Compilation Committee of Yuanmou County Annals. Yuanmou County Annals [M]. Kunming: The Peoples Press of Yunnan, Kunming, 1993: 37–46. [云南省元谋县志编纂委员会. 元谋县志[M]. 昆明: 云南人民出版社, 1993: 37–46.]
[35] WALTHER G R. Plants in a warmer world [J]. Perspect Plant Ecol Evol Syst, 2003, 6(3): 169–185. doi: 10.1078/1433-8319-00076.
[36] HOAGLAND D R, ARNON D I. The water-culture method for growing plants without soil [J]. California Agric Exper Stat Circul, 1950, 347: 32–39.
[37] TANG G L, LI X Y, LIN L S, et al. Effect of short-term girdling on stomatal conductance and chlorophyll fluorescence in[J]. Acta Ecol Sin, 2014, 34(23): 6817–6827. [唐钢梁, 李向义, 林丽莎, 等. 短期环割对骆驼刺气孔导度及叶绿素荧光的影响 [J]. 生态学报, 2014, 34(23): 6817–6827. doi: 10.5846/stxb201303050352.]
[38] LIU Y Q, LIU Y, FAN H M. Review of boron and plant cells and its relationship with other elements [J]. Soil Fertil Sci China, 2009(5): 1– 4. [刘宇庆, 刘燕, 范红梅. 硼对植物细胞的影响及与其它元素关系的研究进展 [J]. 中国土壤与肥料, 2009(5): 1–4.]
[39] HARPOLE W S, NGAI J T, CLELAND E E, et al. Nutrient co- limitation of primary producer communities [J]. Ecol Lett, 2011, 14(9): 852–862. doi: 10.1111/j.1461-0248.2011.01651.x.
[40] YANG K, ZHU J J. The effects of N and P additions on soil microbial properties in paired stands of temperate secondary forests and adjacent larch plantations in northeast China [J]. Soil Biol Biochem, 2015, 90: 80–86. doi: 10.1016/j.soilbio.2015.08.002.
[41] MEI L, SHENG O, PENG S A, et al. Growth, root morphology and boron uptake byrootstock seedlings differing in boron- deficiency responses [J]. Sci Hort, 2011, 129(3): 426–432. doi: 10. 1016/j.scienta.2011.04.012.
[42] WU Z H, LI T H, ZHOU X D. Effects of trace elements on chlorophyll fluorescence parameters ofseedling [J]. Guihaia, 2013, 33(6): 832–838. [吴志华, 李天会, 周旭东. 微量元素营养对桉树无性系苗木叶绿素及荧光参数影响 [J]. 广西植物, 2013, 33(6): 832– 838. doi: 10.3969/j.issn.1000-3142.2013.06.021.]
[43] LI H, TIAN K, LIU G D, et al. Impacts of change in altitude on chloro- phyll fluorescence characteristics of dominant plants in plateau wetland [J]. Acta Ecol Sin, 2018, 38(20): 7421–7434. [李晖, 田昆, 刘国栋, 等. 海拔变化对高原湿地优势植物叶绿素荧光特性的影响 [J]. 生态学报, 2018, 38(20): 7421–7434. doi: 10.5846/stxb201710111820.]
[44] XIANG F, ZHOU Q, TIAN X R, et al. Leaf morphology and PSII chlorophyll fluorescence parameters in leaves ofin different habitats [J]. Acta Ecol Sin, 2014, 34(2): 337–344. [向芬, 周强, 田向荣, 等. 不同生境吉首蒲儿根叶片形态和叶绿素荧光特征的比较 [J]. 生态学报, 2014, 34(2): 337–344. doi: 10.5846/ stxb201210291501.]
[45] VITOUSEK P M, PORDER S, HOULTON B Z, et al. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-pho- sphorus interactions [J]. Ecol Appl, 2010, 20(1): 5–15. doi: 10.1890/ 08-0127.1.
[46] SINSABAUGH R L. Phenol oxidase, peroxidase and organic matter dynamics of soil [J]. Soil Biol Biochem, 2010, 42(3): 391–404. doi: 10.1016/j.soilbio.2009.10.014.
[47] BONIFAS K D, WALTERS D T, CASSMAN K G, et al. Nitrogen supply affects root : shoot ratio in corn and velvetleaf () [J]. Weed Sci, 2005, 53(5): 670–675. doi: 10.1614/WS-05- 002R.1.
[48] GÜSEWELL S. N:P ratios in terrestrial plants: variation and functional significance [J]. New Phytol, 2004, 164(2): 243–266. doi: 10.1111/j. 1469-8137.2004.01192.x.
[49] WANG X M, YAN B G, FAN B, et al. Temperature and soil micro- organisms interact to affectrowth on mountain- sides [J]. Plant Ecol, 2018, 219(7): 759–774. doi: 10.1007/s11258- 018-0832-4.
Effects of Soil Types and Microelements on Growth and Physiological Characteristics ofSeedlings
WANGXuemei1,2, YANBangguo3, WANG Zicheng1, SHI Liangtao3,LIUGangcai2*
(1. School of Resource and Environmental Engineering, Mianyang Normal University, Mianyang 621000, Sichuan, China; 2. Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610041, China; 3. Institute of Tropical Eco-Agriculture, Yunnan Academy of Agricultural Sciences, Yuanmou 651300, Yunnan, China)
To understand the function of microelements ongrowth, the effects of micro- elements (B, Fe, Mn, Zn) on the growth and chlorophyll fluorescence characteristics were studied. The results showed that all the microelements except Mn significantly promoted the growth and chlorophyll fluorescence parameters of(<0.05). Biomass accumulation ofsupplemented with B wassignificantly higher than that of control by 133.61%. Microelement and soil types had significant interactions on leaf P concentration and N/P (<0.05). The addition of Zn in purple soil and Fe in yellow brown soil significantly reduced the leaf N/P. The plant height, leaf area and biomass accumulation ofin dryland red soil and yellow brown soil were higher than those in purple soil, but the root-shoot ratio and leaf N/P ofin purple soil and yellow brown soil were significantly higher than those in dryland red soil (<0.001). Therefore, it was indicated that microelements played an important role of in the growth ofin dry-hot valley, and the plant growth could be promoted by adding B, Fe, Zn, especially B during the vegetation restoration process.
Soil type; Microelement;; Dry-hot valley; Biomass
10.11926/jtsb.4651
2022-04-08
2022-06-03
国家自然科学基金项目(32101363); 绵阳师范学院生态安全与保护四川省重点实验室开放基金项目(ESP1901); 绵阳师范学院科研启动项目(QD2019A04)资助
This work was supported by the National Natural Science Foundation of China (Grant No. 32101363), the Open Project of Ecological Security and Protection Key Laboratory of Sichuan Province, Mianyang Normal University (Grant No. ESP1901), and the Project for Scientific Research of Mianyang Normal University (Grant No. QD2019A04).
. E-mail: liugc@imde.ac.cn
猜你喜欢
中国科学探险(2022年1期)2022-04-16
老友(2020年2期)2020-03-08
中国水土保持科学(2019年6期)2019-04-26
中国环境科学(2017年12期)2018-01-09
诗潮(2017年4期)2017-12-05
红土地(2016年6期)2017-01-15
中国资源综合利用(2016年10期)2016-01-22
云南师范大学学报(自然科学版)(2015年5期)2015-12-26
中国烟草学报(2012年5期)2012-04-12
植物营养与肥料学报(2011年5期)2011-11-06
