
不同加工工艺饲料对彭泽鲫生长性能、抗氧化、免疫和消化能力的影响
2023-10-17 07:17巫伟华陈文静龚海波张平平丁立云
中国饲料 2023年19期
姚 远, 肖 俊, 巫伟华, 陈文静, 龚海波, 张平平, 丁立云
(1.江西省水产科学研究所,江西南昌 330039;2.江西省水生生物保护救助中心,江西南昌 330096;3.南昌湘大骆驼饲料有限公司,江西南昌 330052)
膨化饲料是通过配方粉料充分混合, 在膨化机内经高温高压作用制备得到的蓬松多孔的饲料(李军国等,2022)。 得益于漂浮稳定性好,消化效率高,溶失比率低,污染程度低的特点,膨化饲料在水产养殖的应用前景极其广泛。 在高温高压的制备环境中,原料中的淀粉糊化、蛋白质变性过程更加充分,相比于颗粒饲料,膨化饲料中蛋白酶抑制因子、 淀粉酶抑制因子显著降低(Barrows 等,2007;Francis 等,2001), 鱼类对于淀粉(Gaylord等,2008)、蛋白质(马飞等,2014)的消化利用率显著提升; 饲料体积的增加使得饲料与鱼类肠道接触面积更大,提升了鱼类对饲料的消化效率(Venou 等,2009);同时,膨化饲料提高了鱼类氮、磷利用率,减少了养殖水体污染(刘永士,2018)。
彭泽鲫(Carassius auratus var.Pengze)隶属鲤形目、鲤科、鲤亚科、鲫属,是国内推广程度较高的鲫鱼养殖品种之一,具有生长快、耐低氧、抗病力强、不易脱鳞、肉味鲜嫩、营养价值高、适应地区广、耐长途运输等优良特点(丁立云等,2017)。 目前市场上彭泽鲫养殖使用膨化饲料和颗粒饲料情况均存在, 但关于两种饲料对彭泽鲫养殖进行对比研究鲜有报道。因此,本试验以彭泽鲫为试验对象,探索投喂膨化饲料和颗粒饲料对其生长性能、抗氧化、免疫、消化能力产生的影响,以期能为彭泽鲫养殖饲料的选择及投喂方式提供参考。
1 材料与方法
1.1 试验饲料 试验饲料委托南昌湘大骆驼饲料有限公司加工制作而成。如表1 所示,试验用膨化饲料和颗粒饲料使用相同配方及原料制作。
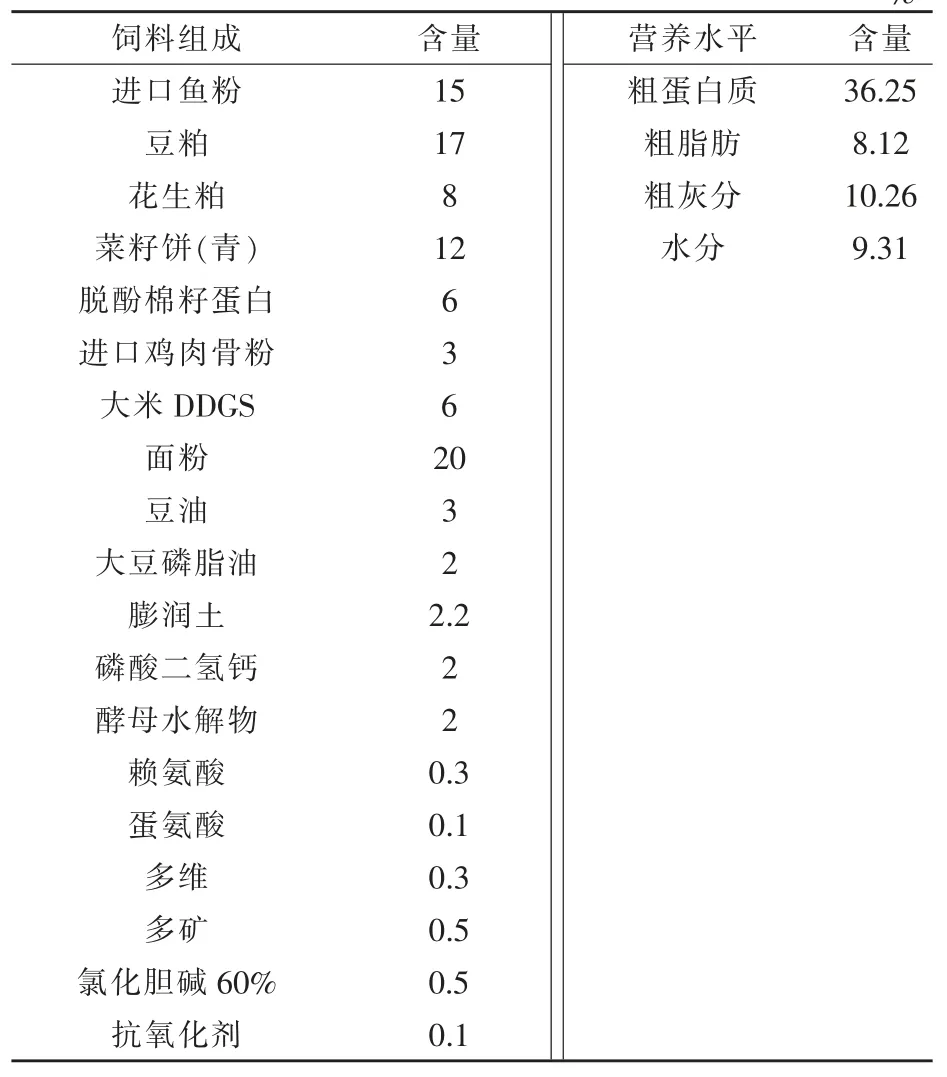
表1 试验饲料组成及营养水平(风干基础) %
1.2 试验鱼与饲养管理 试验用彭泽鲫由江西省水产科学研究所黄马养殖基地提供,试验开始前暂养于三个直径0.8 m, 深0.65 m 圆形养殖桶中,试验前驯化养殖两周,驯化期间投喂商品饲料。 驯化完成后选取活力强、体质优良的个体180 尾,平均初始重量(39.79±0.46)g,试验开始前禁食24 h,随机放置于9 个圆形养殖桶中(规格为直径0.8 m,深0.65 m),每个养殖桶20 尾,养殖周期56 d。 试验分为3 组,每组3 个重复,膨化组全程投喂膨化饲料;交替组采取膨化料和颗粒料交替投喂:第1、3、5、7周投喂膨化饲料, 第2、4、6、8 周投喂颗粒饲料;颗粒组全程投喂颗粒饲料。试验期间使用自动投料机对所有组别进行每日四次精确投喂,投喂时间分别为7:30、10:30、13:30、16:30,日投喂量保持在试验鱼总重的2%左右。 试验期间环境监测情况为: 水温(26.5±1.5)℃,溶氧不低于7 mg/L,pH 为(7.26±0.1),氨氮和亚硝酸浓度不高于0.2 mg/L,光周期利用灯带自动控制模拟日光照射周期。
1.3 取样与分析 试验结束前24 h 开始禁食,称取各个养殖桶内试验鱼总重, 计算增重率及特定生长率。 每个试验养殖桶随机取三尾, 测量体长、体重,之后解剖获取内脏团、肝脏、肠道,测量内脏团及肝脏质量,获取试验鱼的肥满度、脏体比值、肝体比值。 相关公式如下:
增重率=(试验末总重-试验初总重) /试验初总重;
特定生长率 (SGR) =(ln 试验末总重- ln 试验初总重)/试验养殖时长(d);
饲料系数(FCR) = 饲料摄入总量/(试验末总重-试验初总重);
肥满度(CF) = 体重/体长3;
脏体比值(VSI) = 内脏总质量/体重;
肝体比值(HSI) = 肝脏总质量/体重。
鱼体水分测定采用105 ℃烘箱干燥恒质量法;粗蛋白质的测定采用凯氏定氮法;粗脂肪的测定采用乙醚抽提法;粗灰分采用马弗炉550 ℃灼烧法。消化酶、非特异性免疫酶活性采用南京建成生物工程研究所生产的试剂盒测定。
1.4 数据统计与分析 试验数据使用Excel 2022 进行初步统计, 使用SPSS 26.0 进行单因素ANOVA 检验,采取Tukey 检验法对组间差异进行多重比较,差异显著性为P <0.05,试验结果使用“平均值±标准误”表示。
2 结果
2.1 饲料制造方法差异对彭泽鲫生长性能的影响 如表2 所示, 饲料制造方法差异对彭泽鲫的增重率、特定生长率、饲料系数、肥满度、内脏指数、肝脏指数产生了显著性影响。 膨化组增重率显著高于交替组和颗粒组, 分别增加9.01%和32.34%(P <0.05),交替组增重率显著高于颗粒组,增加21.40%(P <0.05)。 膨化组特定生长率显著高于交替组、颗粒组,分别增长6.86%和23.86%(P <0.05), 交替组特定生长率显著高于颗粒组, 增加15.91%(P <0.05)。 膨化组饲料系数显著低于交替组和颗粒组,分别降低9.93%和17.58%,交替组饲料系数较颗粒组降低8.48%(P <0.05)。 膨化组肥满度与交替组无显著性差异(P >0.05),膨化组、交替组肥满度显著高于颗粒组, 分别增加5.34%和3.05%(P <0.05)。膨化组内脏指数与交替组无显著性差异(P >0.05),膨化组、交替组内脏指数显著高于颗粒组, 分别增加10.96%和14.07%(P <0.05)。膨化组肝脏指数与交替组、 颗粒组无显著性差异(P >0.05),交替组肝脏指数显著高于颗粒组,增加25.6%(P <0.05)。
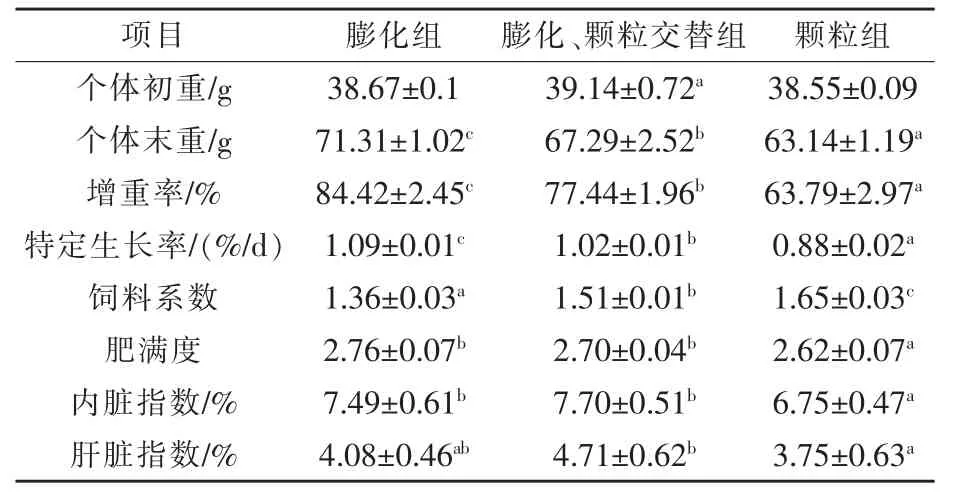
表2 彭泽鲫摄食制造方法差异的饲料对生长性能影响情况
2.2 饲料制造方法差异对彭泽鲫体成分的影响如表3 所示,饲料制造方法差异对彭泽鲫全鱼水分和粗蛋白质产生了显著性影响。 膨化组、交替组的水分显著低于颗粒组,分别降低了4.38%和4.06%(P <0.05)。膨化组、交替组的粗蛋白质显著高于颗粒组,分别提高了8.49%和9.54%(P <0.05)。 膨化组、交替组的粗脂肪显著高于颗粒组,分别提高了11.51%和14.04%(P <0.05)。 饲料制造方法差异对彭泽鲫全鱼灰分没有产生显著性影响(P >0.05)。
2.3 饲料制造方法差异对彭泽鲫抗氧化、 免疫能力的影响 如表4 所示, 膨化组超氧化物歧化酶(SOD) 活性显著高于交替组和颗粒组, 分别提升10.40%和30.77%(P <0.05),交替组超氧化物歧化酶(SOD)活性显著高于颗粒组,提升了18.45%(P<0.05)。 饲料制造方法差异对彭泽鲫肝脏内丙二醛(MDA)含量、总抗氧化能力(T-AOC)没有显著性影响(P >0.05)。 膨化组溶菌酶活性显著高于交替组和颗粒组,分别提升了31.30%和84.15%(P <0.05),交替组溶菌酶活性显著高于颗粒组,增加了40.26%(P <0.05)。 饲料制造方法差异对酸性磷酸酶活性没有显著性影响(P >0.05)。
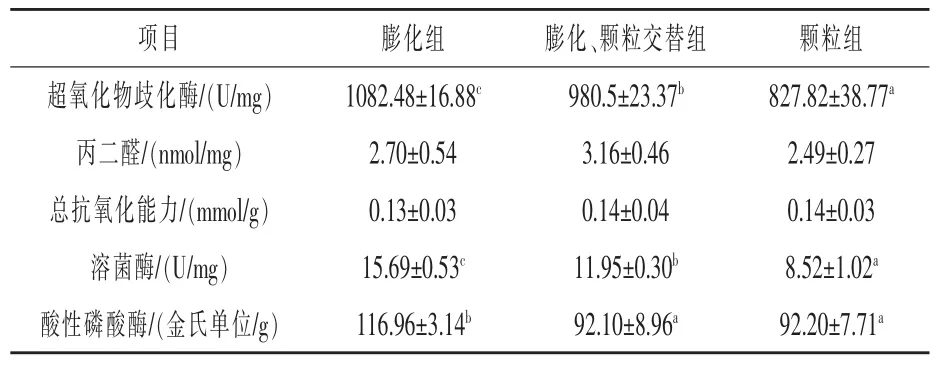
表4 彭泽鲫摄食制造方法差异的饲料对抗氧化、免疫能力影响情况
2.4 饲料制造方法差异对彭泽鲫消化能力的影响 如表5 所示,膨化组、交替组肝脏胰蛋白酶活性显著低于颗粒组, 分别降低67.34%和58.81%(P <0.05)。 膨化组、交替组肝脏淀粉酶活性显著低于颗粒组, 分别降低48.45%和47.26%(P <0.05)。 膨化组肠道淀粉酶活性显著低于颗粒组,降低28.61%(P <0.05), 交替组肠道淀粉酶活性与膨化组、颗粒组没有显著性差异(P >0.05)。 饲料制造方法差异对肝脏脂肪酶、肠道蛋白酶、肠道脂肪酶活性没有显著性影响(P >0.05)。
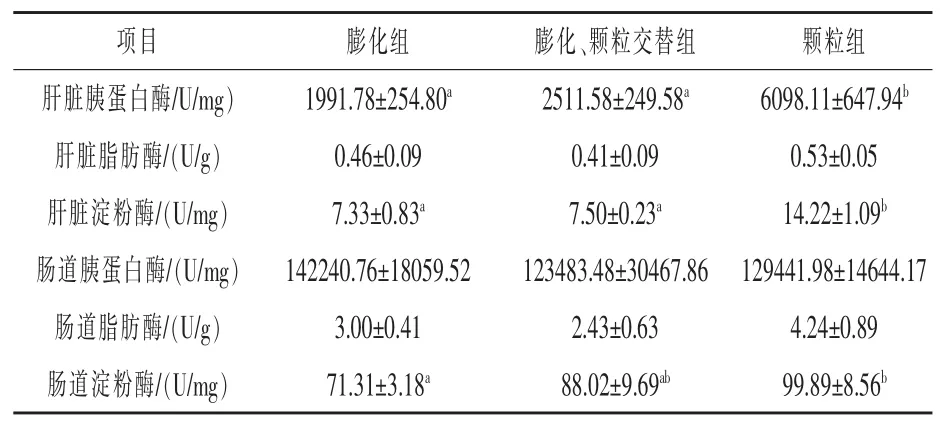
表5 彭泽鲫摄食制造方法差异的饲料对消化能力影响情况
3 讨论
3.1 饲料制造方法差异对彭泽鲫生长性能的影响 大多研究认为, 水产品养殖使用膨化饲料进行投喂综合效果更佳。 对比试验表明, 相同配方下,金头鲷(Venou 等,2003),虹鳟(何川,2004;闫仲双等,2002; 吴秀峰等,2000), 鲟鱼(任华等,2014),草鱼(陈团等,2018)使用膨化饲料投喂生长性能优于颗粒饲料,增重率显著提升,饲料系数显著下降,经济效益提升显著。哲罗鲑(王常安等,2008),泥鳅(李玲丽等,2020)使用膨化饲料投喂增重率、 特定生长率与使用颗粒饲料投喂没有显著性差异,但饲料系数显著下降。也有部分研究发现, 膨化饲料养殖效果相对于颗粒饲料不具有优势,使用膨化饲料投喂鲤鱼,饲料系数没有显著降低,且部分个体出现出血症状(冷永智等,2001);膨化饲料养殖大菱鲆,增重率、特定生长率均显著低于非膨化饲料,饲料系数显著增加,且试验个体出现肝脏病变情况(魏旭光,2015)。本试验结果表明,使用膨化饲料投喂的彭泽鲫增重率、特定生长率、内脏指数显著提升,饲料系数显著下降(P <0.05),使用膨化饲料投喂提升了彭泽鲫的生长性能。 主要原因可能为膨化饲料通过高温膨化工序后,纤维结构被破坏和软化,被获取的可消化物质增加, 淀粉充分糊化且部分淀粉大分子分解为小分子, 相比颗粒饲料能量利用效率更高且更有利于彭泽鲫消化吸收(吴立敏等,2006);膨化饲料可长时间漂浮于水面,更易观察试验鱼进食情况,便于精准投喂,有助于降低饲料系数,同时膨化饲料溶失率更低,高温膨化杀灭了大部分有害微生物,减少了水体污染和病害的发生, 有利于鱼体生长(刘凡等和李艳芳,2016)。
3.2 饲料制造方法差异对彭泽鲫体成分的影响膨化饲料相比于颗粒饲料能否提升水产品营养品质值得进一步研究, 现有关于体成分研究结果不尽相同。 使用膨化饲料投喂哲罗鲑,鱼体水分、灰分显著低于颗粒饲料投喂(P <0.05),粗蛋白质、粗脂肪含量显著升高 (P <0.05)(王常安等,2008);对鲤鱼研究发现,膨化饲料投喂试验鱼水分、 灰分、 粗蛋白质均显著低于颗粒饲料投喂组(P <0.05), 膨化饲料组粗脂肪显著提升 (P <0.05)(罗琳等,2011); 罗非鱼使用膨化饲料投喂水分、灰分、粗脂肪与颗粒饲料投喂无显著性差异(P >0.05),粗蛋白质显著增加(P <0.05)(Ma 等,2016);使用膨化饲料投喂的草鱼水分、灰分、粗蛋白质与颗粒饲料投喂没有显著性差异(P >0.05),鱼体内粗脂肪含量显著增加(P <0.05)(张正洲等,2019)。 赣昌鲤鲫使用膨化饲料投喂水分、灰分、粗蛋白质均无显著性差异,粗脂肪显著低于颗粒饲料投喂组(姚学良等,2020)。本研究表明,相较于全程投喂颗粒饲料,全程使用膨化饲料和交替投喂膨化饲料、颗粒饲料的鱼体水分显著降低(P <0.05),粗蛋白质和粗脂肪含量显著提升(P <0.05),灰分无显著性差异(P >0.05),投喂膨化饲料有助于鱼体营养品质的提升。原因可能是摄食膨化饲料后彭泽鲫拥有更高的蛋白质效率、蛋白质沉积率、脂肪沉积率、能量沉积率,这与银鲈(Booth 等,2002)和罗非鱼(马飞等,2013)的研究结果一致。
3.3 饲料制造方法差异对彭泽鲫抗氧化、 免疫能力的影响 超氧化物歧化酶(SOD)通过清除自由基维持生物体内自由基处于平衡, 丙二醛(MDA)自由基与脂质发生过氧化反应的主要产物, 超氧化物歧化酶和丙二醛含量间接反映了鱼体抗氧化能力的强弱(吴康等,2015)。 溶菌酶(LZM)、酸性磷酸酶(ACP)、碱性磷酸酶(AKP)是鱼体内参与代谢及免疫的非特异性免疫酶,其活性是衡量机体免疫能力的重要标准 (陈剑杰等,2019)。 本研究结果表明, 相较于颗粒饲料, 膨化饲料投喂可显著提升彭泽鲫超氧化物歧化酶 (SOD)、 溶菌酶(LZM)、 酸性磷酸酶(ACP)活性(P <0.05),对丙二醛(MDA)和总抗氧化能力的影响没有显著性影响(P >0.05),证明使用膨化饲料投喂一定程度促进了彭泽鲫抗氧化和免疫能力的提升。 对褐菖鲉 (蒋飞等,2019)和草鱼(陈团等,2018)的研究发现,膨化饲料可显著提升鱼体内超氧化物歧化酶(SOD)活性,本研究结果与其结果一致。
3.4 饲料制造方法差异对彭泽鲫消化能力影响消化酶活性是揭示鱼类消化生理特征的重要指标, 鱼类对于营养物质的吸收能力高低取决于消化酶活性(Fernandez 等,2001),投喂饵料包含的营养成分与含量是鱼体内消化酶活性的重要影响因素(李晨露等,2022)。本试验结果表明,使用膨化饲料饲料投喂的彭泽鲫肝脏胰蛋白酶、肝脏淀粉酶、肠道淀粉酶活性显著低于颗粒饲料组(P<0.05), 投喂膨化饲料有助于彭泽鲫消化能力提升。 原因可能是淀粉在高温环境充分糊化,高温膨化使原料中部分蛋白质性状发生改变,同时破坏了豆粕中的抗胰蛋白酶等抗营养因子,使鱼体运用更少的消化酶即可完成消化(刘凡等和李艳芳,2016;罗琳等,2011;林仕梅,2001)。
4 结论
本研究结果表明, 投喂膨化饲料可提升彭泽鲫生长性能和营养品质, 膨化饲料对彭泽鲫抗氧化、免疫、消化能力有促进作用,单独投喂膨化饲料或使用膨化饲料、 颗粒饲料交替投喂彭泽鲫经济效益更佳。
猜你喜欢
当代水产(2022年8期)2022-09-20
丝绸(2022年5期)2022-05-25
动物营养学报(2022年4期)2022-05-12
Chinese Physics B(2022年3期)2022-03-12
Chinese Physics B(2021年11期)2021-11-23
饲料工业(2017年15期)2017-01-08
渔业致富指南(2016年12期)2016-11-11
兽医导刊(2016年6期)2016-05-17
蚕桑通报(2015年1期)2015-12-23
中国卫生(2014年10期)2014-11-12
