
“阳光玫瑰”葡萄嫁接在不同砧木上的生长表现
2023-10-17 13:31魏灵珠程建徽
中国南方果树 2023年5期
郑 婷,魏灵珠,向 江,程建徽,吴 江
(浙江省农业科学院园艺研究所,杭州,310021)
葡萄是果树生产上的重要树种。我国葡萄产业发展快速,由于栽培技术和设施的完善,葡萄“上山下滩”,种植区域不断扩大,种植面积与产量迅速增加,成为我国6大水果之一[1]。为了提高葡萄的适应能力,人们通常使用砧木进行嫁接[2]。嫁接育苗不仅可以增强葡萄抗病和抗逆能力,还能改善接穗的生长结果习性,加快新品种的繁殖[3-5]。在国际葡萄品种目录数据库中,已有1 343个砧木品种进行了登记[6]。其中,国内常用的砧木主要有“SO4”“5BB”“3309C”“贝达”“1103P”“140R”和“520A”等[7]。
“阳光玫瑰”以“安芸津21号”与“白南”为亲本杂交而成,无论是外观和内在品质,还是品种抗性方面都表现优异[8]。近年来,随着栽培水平的提高,“阳光玫瑰”成为葡萄界的“爱马仕”,受到消费者的一致追捧。简小楠[9]测定了14种砧木对“阳光玫瑰”物候期、生长结果习性、果实品质的影响,发现“3309C”“SO4”“Gloire”是浙江地区最适合“阳光玫瑰”的砧木品种。而刘帅等的研究证实,“阳光玫瑰”自根苗的幼苗光合特性比砧穗组合的光合能力更佳[10]。由于南北方的气候差异,在进行嫁接育苗时,南方主要利用1年生的砧木小苗,北方主要利用多年生的砧木,或者采用高接。
目前,有关“阳光玫瑰”的砧穗组合研究较多,但多集中于其对果实品质的影响,而在葡萄所处生长阶段的转变以及分子水平的验证缺乏理论依据。因此,本研究以“阳光玫瑰”和3种生产常用的砧木“SO4”“5BB”和“3309C”作为试验材料,分别将“阳光玫瑰”嫁接到1年生和多年生的砧木上,结合植物学习性、细胞学观察和分子生物学,研究不同砧木对“阳光玫瑰”生长的影响,旨在明确嫁接对接穗生长状态转变的作用以及对花芽分化能力的影响,筛选“阳光玫瑰”适合的砧木,为生产中“阳光玫瑰”的嫁接栽培提供科学依据。
1 材料与方法
1.1 试材
供试砧木品种为“3309C”“SO4”和“5BB”(见表1),接穗为5年生“阳光玫瑰”葡萄上生长健壮、芽饱满、不带病毒的当年生木质化新梢。试验地点在浙江省农业科学院杨渡试验基地,常规管理。经过对葡萄新梢、叶片等指标的调查,确定了品种的真实性。

表1 3个葡萄砧木品种特性
1.2 嫁接处理
接穗采集:5—6月,当砧木和接穗的新梢均达到半木质化时,早晨采集半木质化的“阳光玫瑰”接穗,去除所有叶片,选择生长充实的新梢中部,留1~2个芽,保留叶柄1~2 cm,并用湿毛巾包裹或放入水中,以防失水。
砧木选择:1年生扦插苗及多年生“3309C”“SO4”“5BB”自根苗,选择生长健壮的新梢,粗度与接穗相近。
嫁接:采用单芽劈接法,将接穗嫁接到1年生扦插苗及多年生自根苗上,砧木新梢保留4~6片叶短截,嫁接后用塑料薄膜包扎。待嫁接芽长出8~10片叶时统计叶片及枝条形态学特征,枝条上、中、下部位的叶片取样,测定叶片长度、叶片宽度、节间长度、节间粗度、叶片数和卷须数。
1.3 石蜡切片制作
取材:取1年生和多年生砧木嫁接的“阳光玫瑰”的上部和下部叶片,用固定液固定24 h以上。将叶片从固定液取出,在通风橱内用手术刀将目的部位组织修平整,将修切好的组织和对应的标签放于脱水盒内。
脱水浸蜡:将脱水盒放进脱水机内依次梯度酒精进行脱水。75%酒精4 h,85%酒精2 h,90%酒精2 h,95%酒精1 h,无水乙醇I 30 min,无水乙醇II 30 min,醇苯5~10 min,二甲苯I 5~10 min,二甲苯II 5~10 min,65 ℃融化石蜡I 1 h,65 ℃融化石蜡II 1 h,65 ℃融化石蜡III 1 h。
包埋:将浸好蜡的叶片于包埋机内进行包埋。先将融化的蜡放入包埋框,待蜡凝固之前将叶片从脱水盒内取出,按照包埋面的要求放入包埋框并贴上对应的标签。于-20 ℃冻台冷却,蜡凝固后将蜡块从包埋框中取出并修整蜡块。
切片:将修整好的蜡块,放入-20 ℃冻台冷却,再将冷却的蜡块置于石蜡切片机切片,厚4 μm。切片漂浮于摊片机40 ℃温水上,将叶片展平,载玻片将叶片捞起,60 ℃烘箱内烤片。水烤干蜡烤化后取出常温保存备用,用于观察叶片细胞结构。
1.4 叶绿素含量测定
利用分光光度计法检测“阳光玫瑰”嫁接苗和自根苗的上部、下部叶片的叶绿素含量,称取0.5 g经液氮研磨过的叶片,加入预冷的95%乙醇5 mL,避光浸提24 h后,8 000 rpm离心20 min,提取上清液后分别测定663 nm、645 nm下的吸光值,Ca=12.72 A663-2.59 A645,Cb=22.88 A645-4.67 A663,C=Ca+Cb,Ca、Cb分别为叶绿素a和叶绿素b的浓度。
1.5 开花相关基因表达检测
利用改良CTAB法从葡萄叶片中提取总RNA,使用PrimeScriptTM RT试剂盒(Takara,Tokyo,Japan)合成cDNA。选择葡萄开花路径关键基因VvFLC、VvSOC、VvTFL、VvFT、VvAP1、VvAP2、VvLFY进行表达水平检测,以VvActin作为内参基因,基因表达水平的计算采用2-△Ct方法[11]。用Primer 5软件设计qRT-PCR引物(见表2)。

表2 引物列表
1.6 数据分析
采用Microsoft Excel 2016软件处理数据和制图,利用SPSS软件进行方差分析,显著性由邓肯氏新复极差法检验(p<0.05)。
2 结果与分析
2.1 不同砧木对“阳光玫瑰”葡萄枝条生长的影响
1年生和多年生砧木对“阳光玫瑰”枝条和叶片的生长有明显影响。嫁接在多年生砧木上的“阳光玫瑰”的叶片长、宽均比嫁接在1年生砧木上的大,而叶片形状没有明显差异(见图1)。嫁接在多年生砧木上的“阳光玫瑰”比1年生砧木的节间长0.38~2.97 cm,其中“5BB”和“3309C”的差异比“SO4”大,嫁接在多年生“5BB”砧木上的节间粗比1年生砧木大0.13 cm(见表3)。
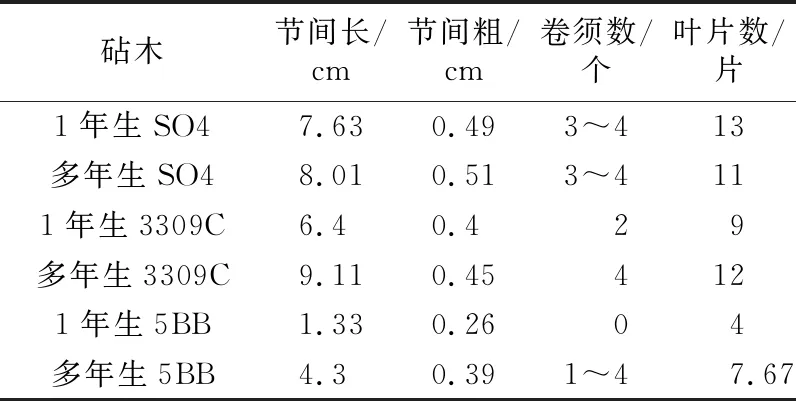
表3 “阳光玫瑰”嫁接在不同砧木上的枝梢生长指标
在3种砧木中,嫁接在“5BB”上的“阳光玫瑰”枝条节间长和节间粗均小于嫁接在“SO4”和“3309C”的“阳光玫瑰”,叶片数和卷须数也更少(见表3)。
2.2 不同砧木对“阳光玫瑰”叶绿素含量的影响
通过检测叶绿素含量发现,下部叶片的叶绿素含量均比上部叶片高,尤其在“SO4”砧木中,下部叶片叶绿素含量最高,“5BB”砧木最低。“阳光玫瑰”嫁接在“5BB”砧木上的长势最弱,整体叶绿素水平比嫁接在“SO4”和“3309C”上的都低(见图2)。
2.3 不同砧木对“阳光玫瑰”叶片细胞结构的影响
葡萄叶片的结构从上到下依次为上表皮、栅栏组织、海绵组织和下表皮。上下表皮由单层细胞构成,排列紧密,上表皮面积大于下表皮。栅栏组织为单层细胞,垂直上表皮分布,圆柱形细胞,排列紧密整齐。海绵组织为多层细胞,分布松散,排列疏松,细胞间隙大。
相比多年生砧木,嫁接在1年生砧木苗上的“阳光玫瑰”的叶片薄壁细胞少、气孔数少、主脉最外侧细胞活跃排列更紧密,内部细胞不规则,组织不发达,栅栏组织排列不紧密,海绵组织所占比例大。嫁接在多年生砧木上的“阳光玫瑰”的茎尖细胞活跃,排列紧密,分生能力强(见图3)。
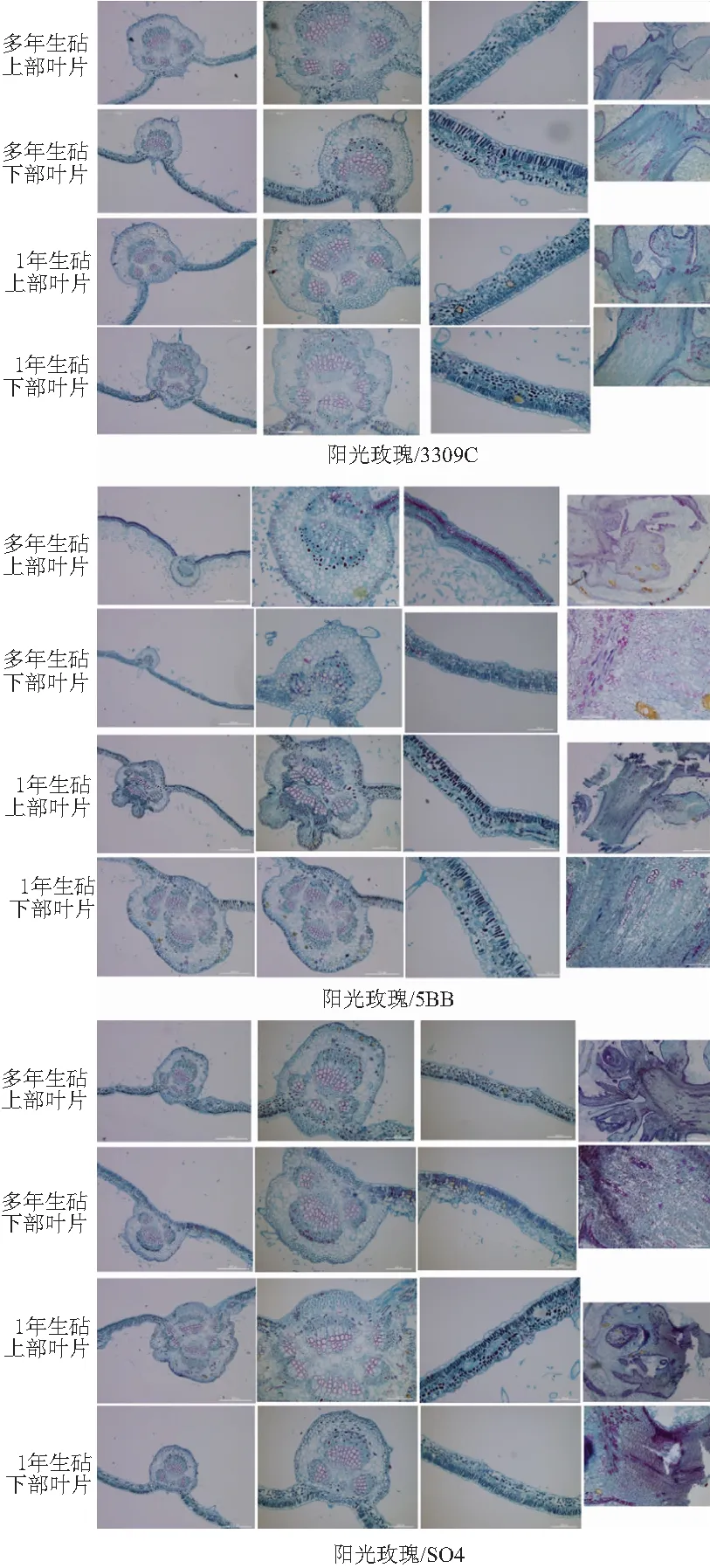
图3 嫁接在1年生和多年生砧木上的“阳光玫瑰”的细胞结构
2.4 不同砧木对“阳光玫瑰”开花相关基因表达的影响
由图4可知,在“阳光玫瑰”自根苗中,7个开花相关基因的表达量依次为VvFLC>VvSOC>VvAP2 >VvTFL>VvLFY>VvAP1 >VvFT。VvFT、VvTFL、VvLFY、VvAP1、VvFLC在砧木“3309C”“SO4”“5BB”中的表达量远高于嫁接“阳光玫瑰”之后。嫁接在1年生“SO4”和“3309C”砧木苗上的“阳光玫瑰”的VvFT、VvTFL、VvAP2、VvSOC、VvFLC的表达量高于嫁接在“SO4”和“3309C”多年生砧木苗上的表达量。“阳光玫瑰”嫁接在1年生的“SO4”和“3309C”砧木苗上,阶段转化特征基因VvFT的表达量高于“阳光玫瑰”自根苗,VvTFL的表达量低于“阳光玫瑰”自根苗。VvLFY、VvSOC、VvFLC在嫁接后的表达量也比在“阳光玫瑰”自根苗中低。在对“5BB”砧木的研究中,由于植株生长状况的原因,呈现出与其他砧木不同的规律,其中,嫁接在多年生砧木上的“阳光玫瑰”的VvFT、VvAP2、VvSOC、VvFLC基因表达量较高,其他基因较低。
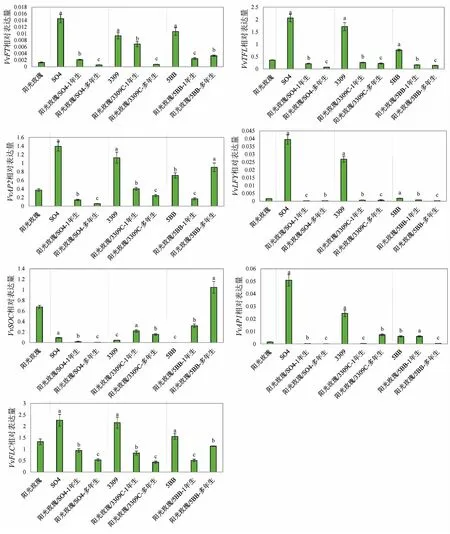
图4 嫁接在不同砧木苗上的“阳光玫瑰”的开花相关基因表达水平
3 讨论
近年来,“阳光玫瑰”以品质佳、抗性强、耐贮运的特点快速抢占了我国葡萄市场。为了提高“阳光玫瑰”的适应性和生长速度,砧木嫁接是生产中最常用的方法[12-14]。砧木能够提高葡萄的抗性,降低非生物胁迫对葡萄的伤害[15-17]。“SO4”“5BB”“3309C”是我国葡萄生产中常用的砧木,刘众杰等[7]研究表明,“SO4”和“3309C”对涝害、干旱、盐害有着较强的抗性,“5BB”对涝害也有一定的抗性,本研究中将“阳光玫瑰”嫁接在“SO4” “5BB”“3309C”3种砧木上,“5BB”的生长状况远不如其他两种砧木。
童期是实生葡萄生长必经的过程,通过无性繁殖虽然没有童期,但也有着营养生长为主的幼树期。在葡萄阶段转化过程中形态学和生理生化物质发生了变化,包括叶面积增大、厚度增加、颜色加深、芽由瘪小细长变饱满钝圆,出现卷须,叶组织发达、薄壁细胞增多、气孔数增多、木质部牢固性降低等[18]。本研究中,嫁接在多年生砧木上的“阳光玫瑰”叶片长、宽均比嫁接在1年生砧木上的大,嫁接在“5BB”砧木上的叶绿素水平比“SO4”和“3309C”低;1年生苗嫁接的“阳光玫瑰”叶片薄壁细胞少、气孔数少、主脉最外侧细胞活跃排列更紧密。多年生苗嫁接的“阳光玫瑰”茎尖细胞活跃,排列紧密,分生能力强,卷须数目更多。花芽分化能力的获得代表了葡萄从营养生长向生殖生长的转换,这一过程与FT、TFL、LFY、MADS、AP2、FLC、SOC、AP3等花芽分化相关基因有关[18-20]。其中FT为成花素基因,诱导开花;TFL1为花序分生组织特征基因,延长营养生长,抑制开花;LFY/AP1抑制TFL1,促进转化;FLC、AP2为抑制成花基因[21-24]。本研究中,由于砧木的品种差异造成了花芽分化相关基因的表达差异,嫁接在1年生“SO4”和“3309C”砧木上的“阳光玫瑰”的VvFT、VvTFL、VvAP2、VvSOC、VvFLC的表达量高于多年生上述砧木,阶段转化特征基因VvFT的表达量高于“阳光玫瑰”自根苗,VvTFL的表达量低于“阳光玫瑰”自根苗,表明嫁接促进了“阳光玫瑰”花芽分化能力的获得,加速了从营养生长向生殖生长的转换。
当然,生长阶段的转换过程受环境、栽培技术等外界条件以及植物生长调节剂、基因表达等内在引物的共同调控[25-27],从而控制新梢生长,改变花芽分化的能力。因此,在实际生产中,在有效利用嫁接技术的基础上,也要关注栽培技术的配套和葡萄园环境的有效调控。
4 结论
综合本研究结果与生产经验说明,嫁接技术可改变葡萄生长阶段的转换,促进花芽分化能力的获得。嫁接在多年生砧木上比嫁接在1年生砧木上的“阳光玫瑰”葡萄植株生长更加健壮,茎尖细胞更活跃,分生能力更强。从“阳光玫瑰”的嫁接结果来看,“3309C”和“SO4”比“5BB”更适合作为“阳光玫瑰”的嫁接砧木。
猜你喜欢
南方林业科学(2022年2期)2022-06-22
落叶果树(2021年6期)2021-02-12
落叶果树(2020年3期)2020-06-17
河北果树(2020年1期)2020-02-09
现代农村科技(2019年3期)2019-03-20
新疆农垦科技(2018年3期)2018-04-24
现代园艺(2017年21期)2018-01-03
湖南农业(2017年1期)2017-03-20
广东农业科学(2016年3期)2016-07-27
工会信息(2016年4期)2016-04-16
