
外源褪黑素对低温胁迫下番茄幼苗光合荧光及生化特性的影响
2023-09-13 02:47于郑娇李书昆褚鹏飞房雨婷王德颖李亚楠吴冰洁
山东农业科学 2023年8期
于郑娇李书昆褚鹏飞房雨婷王德颖李亚楠吴冰洁
(聊城大学农学与农业工程学院,山东聊城 252000)
温度是一个影响植物生长发育的重要因素,低温引起的冷害是植物栽培中经常出现的问题,对植物造成一定的生理危害,减产甚至绝收。 番茄(Lycopersicon esculentum)作为一种喜温蔬菜在15~35 ℃温度范围内均可实现种子萌发和幼苗生长,其同化作用的最适温度为20 ~30 ℃,10 ℃以下生长缓慢,零下1~2 ℃会被冻死。 番茄生长的最适地温为20~23 ℃,降到6 ℃时根系停止生长。 番茄如遇连续阴冷天气,其萌发期和幼果期都会受到低温胁迫影响,会因光照不足而徒长,引起落花落果、坐果率低,产量下降;遇到低温,轻者会生长缓慢,苗弱,叶片皱缩,茎和叶均会变紫,或者出现黄白色花斑或呈黄化状态,严重时叶片呈水浸状,最终会导致全株死亡。 因此,通过提高抗冷性实现番茄增产一直是重要的研究课题。
褪黑素最早在动物体内发现,其研究也是在动物体内进行。 研究发现,褪黑素是一种亲脂性的抗氧化剂,具有较强的抗氧化功能,其与自由基间的反应可以消除一定数量的活性氧,当褪黑素大量增加时则可以加速活性氧的消除速度,减少活性氧的产生[1-2]。 目前,关于番茄低温胁迫的研究主要集中在形态和越冬成活率以及低温胁迫下抗寒蛋白和信号转导等各方面,表明低温下采用ABA(脱落酸)、SA(水杨酸)、CaCl2(氯化钙溶液)进行预处理,能够抑制番茄幼苗体内ROS(活性氧)的产生,提高抗氧化酶活性,减少电解质外渗,降低膜脂过氧化水平,保持细胞质膜的完整性,从而降低低温对番茄幼苗的不利影响,提高幼苗对低温胁迫的耐受性[3-8]。 目前低温对番茄生长发育和各项生理指标的影响已有大量研究:陈胜萍等[9]研究表明,低温胁迫下不同品种番茄各生理指标会有不同程度的变化,而且不同种类番茄在低温胁迫下生理指标的变化有所不同,主要表现在POD、SOD、CAT 和丙二醛上[10]。 较低的温度使番茄幼苗产生大量的自由基和活性氧,导致膜脂过氧化,生物膜透性增强,积累大量过氧化产物即丙二醛,而褪黑素有清除自由基和活性氧的作用,能提高抗氧化酶活性,调节植物幼苗体内氧化反应,同时还具有亲水和亲脂能力,可以在植物细胞内自由活动,以保持番茄幼苗细胞质膜的结构和功能完整性,从而减轻番茄幼苗在逆境中受到的伤害。
1 材料与方法
1.1 试验材料
供试微型番茄品种为Micro-Tom。
外源褪黑素溶液:褪黑素粉剂(纯度98%,购于上海麦克林生化科技股份有限公司)用乙醇溶解后再用无菌水配制成所需浓度,分别为50、100 μmol/L 和150 μmol/L。
1.2 试验设计及处理
1.2.1 番茄幼苗培育选取健康饱满的Micro-Tom 野生型番茄种子,用75%乙醇浸泡2 min,后用无菌水冲洗3 ~4 次,随即用3%次氯酸钠溶液消毒处理,再将种子用无菌水浸泡15 min,用无菌水多次冲洗,彻底除净种子表面附着的残留药液。之后,将种子放进湿润培养皿中,用湿润纱布覆盖后放到光照培养箱中避光室温条件下催芽生根,待根长至1 ~2 cm 时种入穴盘(32 孔穴,上口径6.0 cm,底径2.6 cm,高5.3 cm)中。 穴盘土壤以育苗土和蛭石按1 ∶1混合,湿润程度以手握成团不滴水为宜,约为田间持水量的50%。 用保鲜膜将穴盘包裹住,保持穴盘中水分不易散失,短期内不用浇水。 三叶一心时将番茄幼苗移入小花盆(上口径14.8 cm,底径7.2 cm,高9.8 cm)中,土壤改为腐殖土与蛭石按1 ∶1混合。 整个育苗过程在适宜温度(20~25 ℃)下的光照培养室(聊城大学农学院植物种质资源创新与利用实验中心)中进行。
1.2.2 褪黑素以及低温处理待幼苗长到五叶一心时,于当天下午5 ∶30—6 ∶00 喷施1 次褪黑素,以叶面凝结水滴为准。 每隔24 h 喷施1 次,共喷施3 次。 之后,常温(25 ℃)环境下放置48 h再移至光照培养箱中进行低温处理,条件为温度4 ℃、光强60 μmol/(m2·s)和湿度70%。 试验共设置5个处理:CK,常温(25 ℃)+叶面喷施清水,简称常温0;低温+叶面喷施清水(0 μmol/L 褪黑素溶液),简称低温0;低温+叶面喷施50 μmol/L褪黑素溶液,简称低温50;低温+叶面喷施100 μmol/L 褪黑素溶液,简称低温100;低温+叶面喷施150 μmol/L 褪黑素溶液,简称低温150。 每处理15 盆,每盆1 株,重复5 次。 每处理选6 株,于低温处理后第1、2、3、4 天分别进行光合参数和叶绿素荧光参数以及SPAD 值测定。
1.3 测定项目及方法
1.3.1 光合参数测定于低温处理后第1、2、3、4天,将番茄幼苗移至光照下处理30 min,之后选择完全展开功能叶,利用LCPro-T 便携式光合作用仪,设定光强600 μmol/(m2·s),测定叶片净光合速率(Pn)、胞间二氧化碳浓度(Ci)、气孔导度(Gs)。
1.3.2 叶绿素荧光参数测定于低温处理后第1、2、3、4 天,将番茄幼苗暗处理30 min,随后使用Handy PEA 荧光仪在弱光下测定叶片初始荧光(Fo),再用3 000 μmol/(m2·s)红光强饱和脉冲光激发,使QA 全部处于还原状态,测得最大荧光值(Fm)[16]。 测出番茄叶片PSⅡ的最大光化学效率Fv/Fm、反应中心吸收的光量子将电子传递到电子传递链中Q-A下游的其它电子受体的概率(φEo)和PSⅠ受体侧末端电子受体还原的量子效率(φRo)。
1.3.3 SPAD 值测定于低温处理后第1、2、3、4天用便携式SPAD 叶绿素仪测定。 每重复随机选取4 片叶测定SPAD 值,每片叶至少采15个点,避开叶脉,取其平均值,即为叶片叶绿素相对含量。
适用于不对称情况的线换相换流器动态相量模型//曾淑云,江全元,陆韶琦,赵敏,徐希望//(11):129
1.3.4 生化指标测定过氧化物酶(POD)活性测定采用愈创木酚法[17],过氧化氢酶(CAT)活性测定采用紫外分光光度法[17],超氧化物歧化酶(SOD)活性测定用NBT(氮蓝四唑)光化学还原法[18],丙二醛(MDA)含量测定用硫代巴比妥酸法[19]。
1.4 数据处理与分析
采用SigmaPlot 14.0 软件处理数据和作图,采用SPSS 16.0 软件,通过单因素方差分析(oneway ANOVA)和Duncan’s 法进行多重差异显著性分析(α =0.05)。
2 结果与分析
2.1 不同浓度褪黑素对低温胁迫下番茄叶片光合指标的影响
由图1 看出,与常温0(CK)相比,低温处理组番茄叶片的Pn、Gs 显著下降,Ci 在低温处理2 d 后显著上升;连续4 天低温处理番茄幼苗受冻损伤明显,Pn、Gs 都呈显著下降趋势,Ci 则呈显著上升趋势。
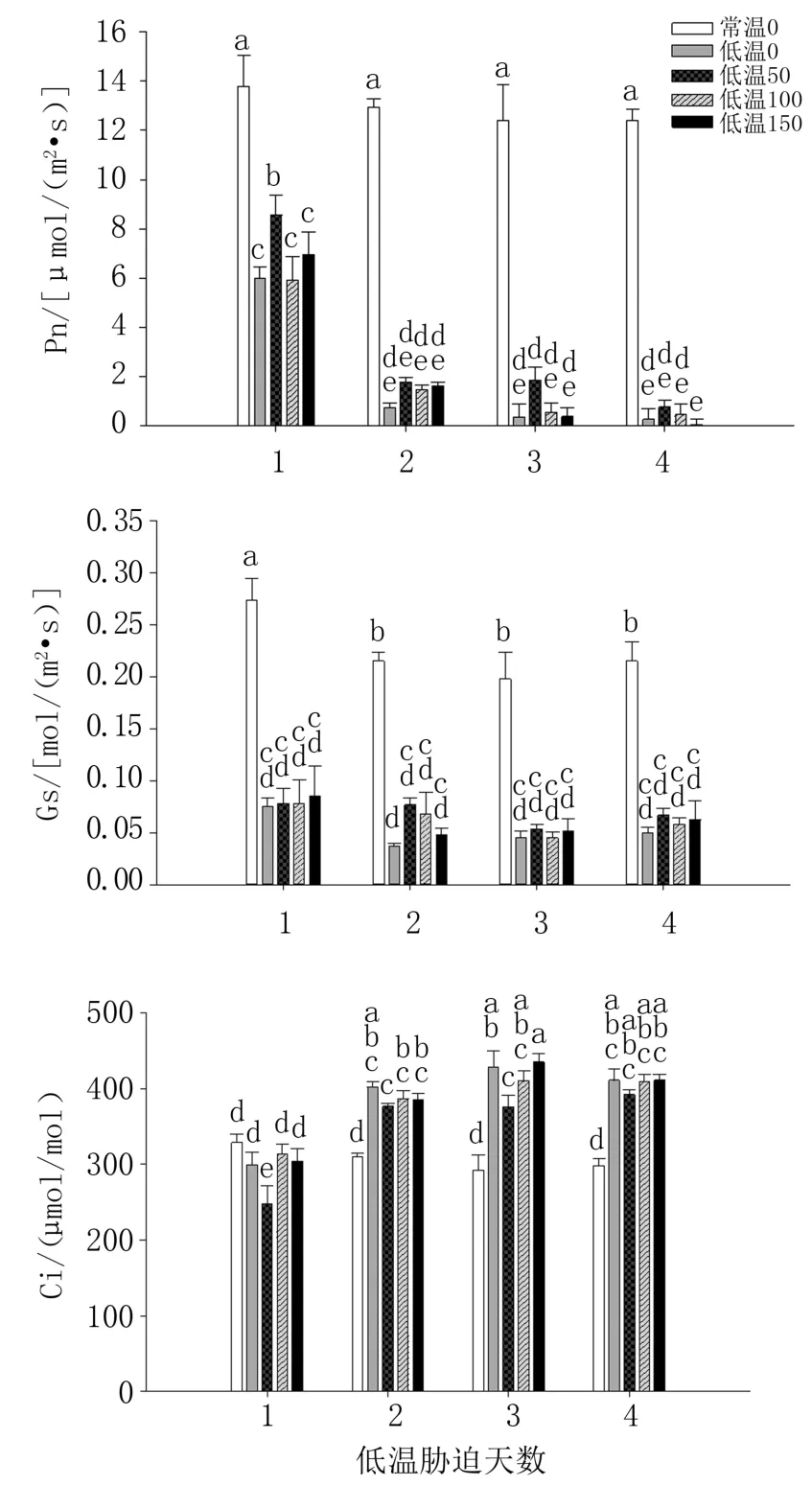
图1 外源褪黑素对低温胁迫下番茄叶片Pn、Gs、Ci 的影响
低温胁迫下不同浓度褪黑素处理对番茄生长影响不同,与低温0 处理相比,施用褪黑素可以有效缓解低温胁迫造成的光合抑制。 低温胁迫第1天,低温50 处理叶片Pn 比低温0、低温100、低温150 分别高43.25%、44.74%和23.18%(P<0.05);而第2 天至第4 天,低温处理间Pn 差异不显著。第1 天,低温50 处理Ci 比低温0、低温100、低温150 分别低17.10%、20.93%和18.42%(P<0.05)。低温处理间叶片Gs 差异不显著。
综上,低温处理组中,50 μmol/L 外源褪黑素处理番茄叶片Pn、Gs 高于其它处理,Ci 最低。 因此,该处理可通过保持较高的Gs 和降低Ci 而提高Pn,从而缓解低温逆境伤害。
2.2 不同浓度褪黑素对低温胁迫下番茄叶片叶绿素荧光指标的影响
由图2A 看出,与常温0(CK)相比,低温处理组番茄叶片Fv/Fm 均降低。 低温处理第1 天,4个低温处理番茄叶片的最大光化学速率(Fv/Fm)相近,低温50 处理稍高;第2 天,低温处理组变幅较大,低温50 高于低温0 处理22.50%(P<0.05);第3 天、第4 天低温处理组Fv/Fm 整体大幅下降。
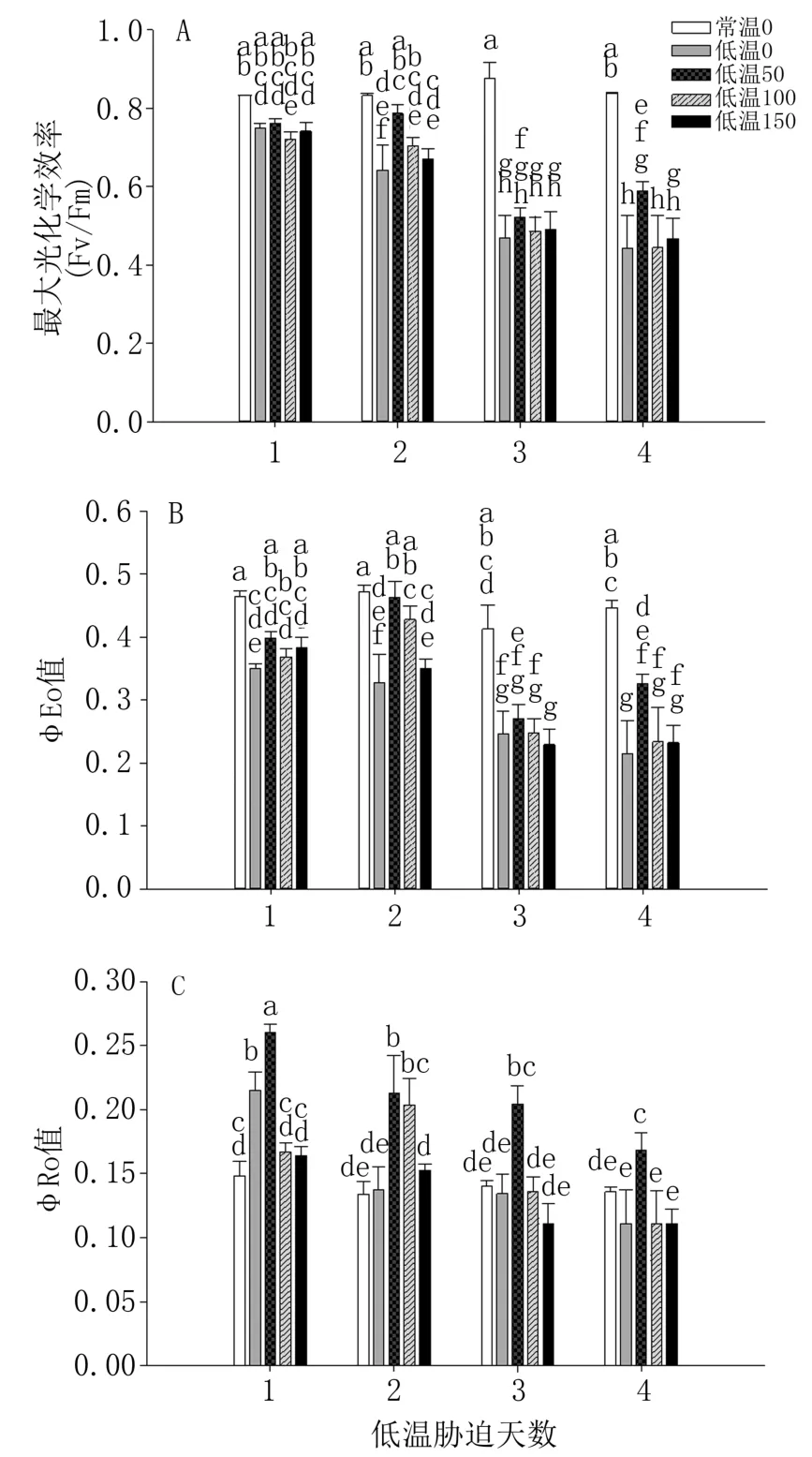
图2 外源褪黑素对低温胁迫下番茄叶片叶绿素荧光指标的影响
图2B、C 显示,番茄叶片φEo 值低温组与CK相比降低,而φRo 值在低温处理的前两天升高,第3 天开始除低温50 处理外也均降低。 低温处理组番茄叶片φEo 值和φRo 值,整体上以喷施50 μmol/L 褪黑素处理效果最优。 低温50 处理叶片φEo 值在第2 天分别高于低温0、低温150 处理41.50%和31.94%(P<0.05)。 低温处理第1 天、第2 天叶片φRo 值,低温50 处理高于常温及其余低温处理组,且在第1 天高于低温0、低温100处理21.11%和37.63%(P<0.05),而第3 天、第4天低温50 处理仍然最高,而其它低温处理则明显下降,均低于CK。
低温下外源褪黑素4个浓度处理相比,50 μmol/L 处理番茄叶片的Fv/Fm、φEo 和φRo 均高于其它3个浓度处理。
2.3 不同浓度褪黑素对低温胁迫下番茄叶片SPAD 值的影响
由图3 可知,低温处理前3 天,50 μmol/L 褪黑素处理的番茄叶片SPAD 值明显高于常温0 处理(CK),但差异不显著;第4 天,与CK 相近。 低温处理组间相比较,50 μmol/L 褪黑素处理番茄叶片的SPAD 值要高于其它3个处理,对番茄叶片叶绿素含量的促进效果最优。
2.4 外源褪黑素对低温胁迫下番茄幼苗抗氧化酶活性的影响
2.4.1 对POD 活性的影响如图4A 所示,常温对照番茄幼苗POD 活性基本保持稳定。 前3 天,整体上低温胁迫组幼苗POD 活性上升,第4 天番茄幼苗受到较大伤害,清除活性氧能力降低,POD活性有所下降。 其中,50 μmol/L 褪黑素处理幼苗POD 活性最高,效果最好;100、150 μmol/L 高浓度褪黑素处理POD 活性降低,但仍显著高于对照,对提高幼苗低温胁迫耐受性仍有一定效果。
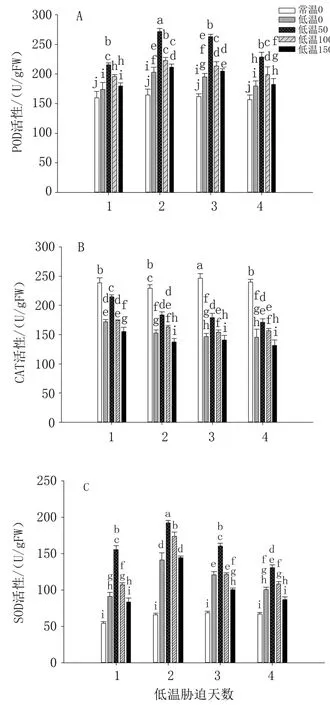
图4 外源褪黑素对低温胁迫下番茄叶片POD、CAT、SOD 活性的影响
2.4.2 对CAT 活性的影响由图4B 可知,常温对照番茄幼苗CAT 活性最高且基本平稳,低温胁迫下幼苗CAT 活性呈下降趋势,即低温处理第1天CAT 活性最高,长时间低温胁迫使其清除活性氧能力减弱,CAT 活性降低。 各褪黑素浓度处理中,低温胁迫第1 天至第4 天,50 μmol/L 处理幼苗CAT 活性均最高,提高幼苗低温胁迫耐受性效果最强,即提高幼苗抗逆性最显著;褪黑素浓度为100、150 μmol/L 时,CAT 活性逐渐降低,说明过高浓度的褪黑素会抑制CAT 活性。
2.4.3 对SOD 活性的影响由图4C 可知,随着低温胁迫时间延长,番茄幼苗SOD 活性呈先升高后降低趋势,第2 天时最高,之后降低,清除活性氧能力下降。 4个浓度褪黑素处理幼苗SOD 活性均高于对照,可见喷施褪黑素可以提高低温胁迫下幼苗SOD 活性。 其中,褪黑素浓度为50 μmol/L时SOD 活性最高,幼苗低温胁迫耐受性最强,即提高幼苗抗逆性的效果最显著;褪黑素浓度为100、150 μmol/L 时,其SOD 活性逐渐降低,幼苗抗逆性效果下降。
2.5 外源褪黑素对低温胁迫下番茄丙二醛含量的影响
由图5 可知,低温处理1 天时,各处理番茄幼苗的MDA 含量最低,随着低温胁迫时间延长而逐渐升高。 第2 天至第4 天(低温0)处理幼苗的MDA 含量均较高,各时间点50 μmol/L 褪黑素处理的MDA 含量均最低,浓度增至100、150 μmol/L时幼苗受到高浓度褪黑素的抑制,MDA 含量升高。 由此可见,50 μmol/L 褪黑素处理提高番茄幼苗低温胁迫耐受性的效果最好,能够有效减轻低温对幼苗的伤害。 表明喷施适当浓度的外源褪黑素可有效减轻番茄幼苗低温胁迫后的膜脂过氧化作用,清除活性氧,维持膜系统结构和功能的完整性,减轻低温对细胞膜系统的伤害。
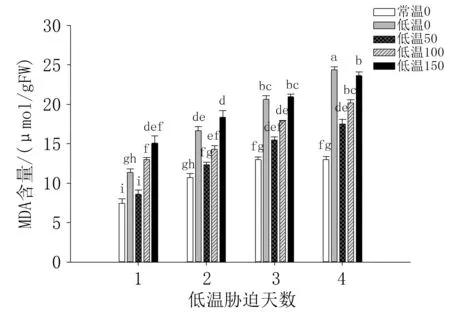
图5 外源褪黑素对低温胁迫下番茄叶片MDA 含量的影响
3 讨论
褪黑素的首次提取是在牛的松果体中,并且随即证实它是一种类激素物质。 褪黑素的出现不断成为学者研究的热点,无论是在动物还是植物上的研究都被广泛关注,不断出现成功应用型案例。 前人研究显示,50 μmol/L 外源褪黑素处理大豆植株,能显著促进其生长发育且增产[20]。 向葡萄幼果喷施100 mg/L 褪黑素溶液,能够促进果实生长和膨大[21]。 喷施适宜浓度的外源褪黑素可以明显缓解高温逆境胁迫对黄瓜光合器官造成的损伤[15]。 园艺作物在生物胁迫及非生物胁迫下喷施褪黑素都具有一定程度的抗性调控作用[22]。
温度是影响园艺作物生长发育最重要的环境因素,尤其是反季节栽培蔬菜的关键环境因子。一定浓度的外源褪黑素可以降低光合系统在低温胁迫下的受损程度[23]。 低温胁迫对园艺植物尤其是冷敏性植物会产生较大影响,严重影响其生长状态和产量。 冯建灿等[24]研究发现,喜树叶片在低温条件下其SPAD 值呈现下降趋势;马德华等[25]也发现,低温条件下黄瓜幼苗叶片SPAD 值也明显下降;黄伟等[26]认为,低温处理后,番茄叶片的SPAD 值明显下降。 低温不但影响叶绿素的合成,还会造成叶绿素的分解强度加大,光合速率下降[27]。 已有研究表明,低温胁迫主要抑制PSⅡ的活性,而对PSⅠ的影响相对较小,降低电子传递效率[28-29]。 本试验中,低温处理组与常温对照相比,其Pn 和Gs 降低,Ci 值升高,说明非气孔限制影响番茄叶片的光合作用;低温胁迫下喷施褪黑素处理的Pn 和Gs 值呈现升高趋势,而Ci 值则下降,说明褪黑素可缓解低温对番茄光合作用的影响。 本试验中,低温与常温处理相比,胁迫前两天番茄叶片SPAD 值差异不显著,第4 天低温处理的SPAD 值有所降低;低温处理组中,50 μmol/L 褪黑素处理的SPAD 值最高,这说明喷施褪黑素可以缓解叶绿素的分解,提高光合速率。
本试验中,连续低温导致番茄叶片的最大光化学效率(Fv/Fm)明显下降,说明低温引起了番茄叶片的光抑制,而低温50 处理与低温0 相比,Fv/Fm 值明显增加,说明喷施褪黑素可以缓解低温对番茄叶片的光抑制。 φRo 是指用于还原PSⅠ受体侧末端电子受体的量子产额[30],φEo 则反映了反应中心吸收的光能将电子传递到电子传递链中Q-A下游的其它电子受体的概率[31]。 本试验中,低温胁迫第1 天,低温0 处理φRo 值比常温对照升高44.83%,后3 天降低,第4 天低温0 比常温0 下降18.31%;而φEo 值低温0 与常温0 相比,4 天分别降低24.62%、30.76%、40.53%和51.83%(P<0.05)。这表明低温对PSⅠ和PSⅡ的电子传递产生不同影响,连续低温抑制PSⅡ的活性,对于PSⅠ呈现后期抑制趋势,连续低温对于PSⅡ受体侧的伤害更大些。 有研究表明,裉黑素上调PSⅠ反应中心亚基(PsaD、PsaE、PsaF、PsaH 和PsaN)、PSⅡ相关蛋白PsbA(D1)和核酮糖-1,5-二磷酸羧化酶或加氧酶大亚基(RBCL) 和Rubisco 激活酶(RCA)的蛋白[32]。 外源褪黑素通过调控叶黄素循环基因CsVDE与CsVDE的表达来加强叶黄素循环组分的脱环化程度,从而提高调节性能量耗散的占比,减少过多激发能对光系统造成的伤害,进而维护光系统Ⅱ光能利用的最大潜能。 低温处理2 天后,低温50 处理比低温0 的叶绿素荧光指标值均明显增高,表明适宜浓度褪黑素处理可缓解低温逆境对番茄幼苗光系统的影响。 褪黑素可提高植物的抗寒性,其在酶、蛋白质、生物小分子水平的作用机理尚不明确。 在遭遇低温胁迫后,褪黑素还可通过上调冷响应途径的关键转录因子CBFs/DREBs 和低温应答基因COR15a的表达水平,诱导转录激活因子CAMTA1、ZAT10 和ZAT12的表达,从而提高拟南芥的冷冻耐受性[33]。 褪黑素可以作为信号分子在逆境胁迫中起重要作用,然而对于植物如何感受褪黑素受体和信号转导途径仍未明确,这也将是褪黑素研究的一个重要方向。
包宇等[34]研究表明,对番茄幼苗(大红番茄“合作903”)进行100 μmol/L 褪黑素预处理之后,其相对电导率和MDA 含量比对照显著降低,抗氧化酶活性最高,说明该浓度褪黑素对提高番茄幼苗低温胁迫耐受性的效果最好。 这与本试验结果不同的原因之一可能为所用番茄品种不同:就番茄特性而言,大红番茄“合作903”比Micro-Tom 野生型番茄形体大,茎粗叶大,抗氧化活性高,低温胁迫耐受性较好。 唐鸿吕等[35]的研究表明,甘蓝在低温胁迫24 h 后,各组幼苗表型有明显变化,主要表现为叶片失水萎蔫;处理48 h 后,喷施外源褪黑素处理可提高抗氧化酶活性和SPAD 值,减少电解质外渗,提高甘蓝受冷害后的恢复能力,降低低温胁迫对甘蓝的伤害,幼苗均有所恢复;72 h 低温胁迫则受到严重伤害。 相比较而言,甘蓝是耐冷性植物,而番茄为一种冷敏性植物,当温度低于5℃就会停止生长,长时间低温胁迫使番茄幼苗受到严重伤害,本试验低温处理2天时番茄的抗氧化酶活性和丙二醛含量最高。
综上所述,喷施外源褪黑素能够有效清除番茄植株细胞内的自由基和活性氧,维持细胞内水分和膜系统功能结构的完整性,从而保持细胞膨压,增强番茄幼苗低温胁迫的耐受性,尤以50 μmol/L 褪黑素处理提高幼苗低温胁迫耐受性的效果最好。
4 结论
本试验中,3个褪黑素处理相比,低温胁迫下喷施50 μmol/L 褪黑素处理番茄叶片的净光合速率(Pn)最高,气孔导度(Gs)除胁迫第1 天较高外其它时间点也均最高,胞间二氧化碳浓度(Ci)均最低;叶绿素荧光指标中,其最大光化学效率(Fv/Fm)、反应中心吸收的光能将电子传递到电子传递链中Q-A下游的其它电子受体的概率(φEo)和PSⅠ受体侧末端电子受体还原的量子效率(φRo) 均最高,SPAD 值也均最高;SOD、POD、CAT 活性也均最高,即能通过提高抗氧化酶活性来调节番茄幼苗体内的氧化反应,提高番茄低温胁迫的耐受性。 同时,50 μmol/L 褪黑素处理在低温胁迫2 天时提高幼苗抗逆性的效果最明显。低温胁迫下,褪黑素浓度增至100、150 μmol/L时幼苗受到高浓度褪黑素的抑制,提高幼苗抗逆性的效果降低。 随着低温胁迫时间延长,过氧化产物丙二醛(MDA)大量积累,喷施50 μmol/L 褪黑素处理番茄幼苗的MDA 含量最低,从而减轻低温对细胞膜系统的伤害。 以上表明,低温逆境下喷施50 μmol/L 褪黑素提高番茄抗逆能力的效果最好。
猜你喜欢
钻井液与完井液(2022年4期)2022-10-26
哈尔滨轴承(2021年1期)2021-07-21
园林科技(2020年2期)2020-01-18
电镀与环保(2017年3期)2017-06-23
天津科技大学学报(2016年1期)2016-02-28
山东医药(2015年38期)2015-12-07
中国医学科学院学报(2015年5期)2015-03-01
中国医药科学(2015年15期)2015-02-27
金属矿山(2014年7期)2014-03-20
食品工业科技(2014年13期)2014-03-11
