
玉米雄穗主轴长和分枝数QTL定位
2023-09-11 03:21吴昊王瑞泽何永辉梁爽刘欢欢
江苏农业科学 2023年15期
关键词:玉米
吴昊 王瑞泽 何永辉 梁爽 刘欢欢
摘要:合理的雄穗结构在协调玉米株型、增加生物产量方面具有重要的意义,而雄穗结构的2个重要指标分别是雄穗主轴长和分枝数。本研究利用玉米雄穗主轴长和分枝数具有显著差异的自交系Y915和郑58构建重组自交系(RIL)群体,在连续自交6代后,利用高密度分子遗传图谱在3个环境(2020年春扬州、2021年春扬州和2021年夏镇江)对玉米雄穗主轴长(TL)和分枝数(TBN)进行QTL分析。结果共检测到6个雄穗主轴长QTLs,分布于1号、8号和10号染色体上,贡献了5.10%~14.63%的表型变异;4个雄穗分枝数相关性状QTLs,分布于1号、4号和6号染色体,贡献了5.69%~10.56%的表型变异。其中在多个环境下检测到qTBN1-1位点,并且此区间上还有1个控制雄穗主轴长的位点qTL1-1。本研究将为挖掘玉米雄穗主轴长和分枝数基因和分子标记辅助品种改良提供参考。
关键词:玉米;雄穗主轴长;雄穗分枝数;QTL定位
中图分类号:S513.032 文献标志码:A
文章编号:1002-1302(2023)15-0016-05
基金项目:江苏省种业揭榜挂帅项目(编号:JBGS[2021]055);江苏省种业振兴揭榜挂帅项目(编号:JBGS[2021]002);江苏高校优势学科建设工程资助项目。
作者简介:吴 昊(1996—),男,江苏扬州人,博士研究生,主要从事玉米遗传研究。E-mail:874856414@qq.com。
通信作者:刘欢欢,博士,讲师,主要从事玉米遗传研究。E-mail:liuhh@yzu.edu.cn。
玉米是世界上极为重要的粮饲作物,在美国、中国、巴西等国家广泛种植[1]。玉米雄穗性状对产量有较大的影响,其中雄穗分枝数一般对产量有负作用。李建波研究发现,去除雄穗后玉米产量提高[2]。Lambert等研究发现,雄穗分枝数对玉米产量有负面影响[3-4],这可能是由于过多的雄穗分枝数消耗大量养分,加剧与雌穗的竞争;同时由于产生的花粉过多,玉米叶片表面接受阳光面积变小,削弱了叶片的光合作用。较长的雄穗往往伴随着更长的散粉时长,在高温、干旱等逆境下有较好的籽粒结实率[5]。因此,选育大小合适的雄穗玉米品种,是玉米育种的目标之一。
玉米雄穗主轴长和分枝数是由多个位点协同控制的数量遗传性状。汤华等利用杂交种豫玉22号连续自交得到266份材料的F2:3群体,在武汉和襄樊2个环境下检测到9个雄穗分枝数的数量性状基因座(QTL),其中有5个在2个环境下均能被检测到[6];杨钊钊等在2个以黄早四为共同亲本的重组自交系(RIL)群体中都检测到一个主效雄穗一级分枝数QTL,贡献率分别为17.4%和14.4%,且位点高度重叠[7];高世斌等运用复合区间作图法检测N87-1与9625配制的含183个家系的F3分离群体,在干旱和正常2个环境下共检测到雄穗主轴长和分枝数的11个QTL,其中有2个QTL在2个环境下均能被检测到[8];代资举等以郑58和昌7-2为亲本组建RIL群体,在2年重复检测到5个雄穗分枝数QTL,其中,qTBN5的表型贡献率为19.92%,并将其精细定位到13.2 Mb区间之内[9];张媛等利用雄穗分支数差异明显的甜玉米T14与T4配制F2、F2:3群体,检测到分别位于第2、4号染色体上的3个表现稳定的雄穗分枝数QTLs[10]。除了以上QTL定位结果以外,Guan等利用B73和CML247构成的RIL群体检测到4个雄穗分枝数QTL位点,并克隆到负调节雄穗分枝数的基因ZmPAT7,当ZmPAT7基因启动子发生变异,表达量下降以后,玉米的雄穗分枝数得到显著提高[11]。尽管国内外开展了很多定位和克隆玉米雄穗主轴长和分枝数相关基因的工作,但由于研究中使用的群体种类、遗传背景、环境条件等各因素的差异,能有效控制雄穗主轴长和分枝数的基因被发掘的不多,因此进一步对雄穗主轴长和分枝数的QTL定位和基因发掘工作是十分必要的。
本研究以雄穗主轴长和分枝数有显著差异的Y915和郑58为亲本,经过多年连续自交得到了包含171个家系的RIL。通过构建的高密度遗传图谱对雄穗主轴长和分枝数性状进行QTL分析,以期能够在多个环境下找到稳定遗传的QTL位点,为玉米雄穗性狀的改良以及选育理想雄穗株型提供参考。
1 材料与方法
1.1 试验材料
利用笔者所在实验室玉米自交系Y915和郑58在2017年杂交得到F1,在扬州和海南两地连续自交6代后得到包含171个家系的重组自交系群体。
1.2 田间试验设计
171个重组自交系群体以及亲本在2020年和2021年分别种植于3个环境中(2020年春江苏省扬州市扬州大学试验田、2021年春江苏省扬州市扬州大学试验田、2021年夏江苏省镇江市世业洲试验田,分别简称为20春扬州、21春扬州、21夏镇江)。田间试验采用随机区组试验设计,设置2个重复:行长2.5 m,短行行距30 cm,长行行距90 cm,每行种植10株,株距25 cm,田间管理按照当地正常栽培措施进行管理。
1.3 田间性状调查
表型数据以行为单位进行调查。每行植株中,除边上2株外,随机选取3株对表型值进行调查,以3株玉米测定值的平均值作为此行的表型值。本研究于植株散粉期后20 d共调查了RIL群体中的雄穗相关性状:雄穗主轴长(TL)和雄穗分枝数(TBN)。雄穗主轴长为雄穗穗颈节到雄穗顶部的长度(cm)[12];雄穗分枝数为雄穗主轴及其分枝数目[13]。
1.4 数据处理
利用SPSS 21.0软件对测定的各表型数据进行描述性统计和相关性分析,利用Microsoft Excel 2010处理分析基因型数据。
1.5 连锁图谱构建与QTL分析
将亲本以及重组自交系送样至北京中玉金标记生物技术股份有限公司,进行maize10K基因型检测。参考陈俊宇等的方法[14]构建连锁遗传图谱。
QTL 定位使用WinQTL Cartographer 2.5软件进行,方法采用复合区间作图法,使用 kosambi函数转化图距,作图步长为1.00 cM (centimorgan,简称cM)。QTL的阈值设为2.5,QTL的置信区间为从LOD曲线的最高点下降1.5个LOD值的标记区间[15-16]。
本研究中 QTL命名参考 McCouch等的方法[17],规则如下:雄穗相关性状的QTL以qTBN1-1为例,q表示QTL,TBN为性状英文字母缩写,1-1表示在1号染色体检测到的第1个QTL。
2 结果与分析
2.1 遗传图谱的构建
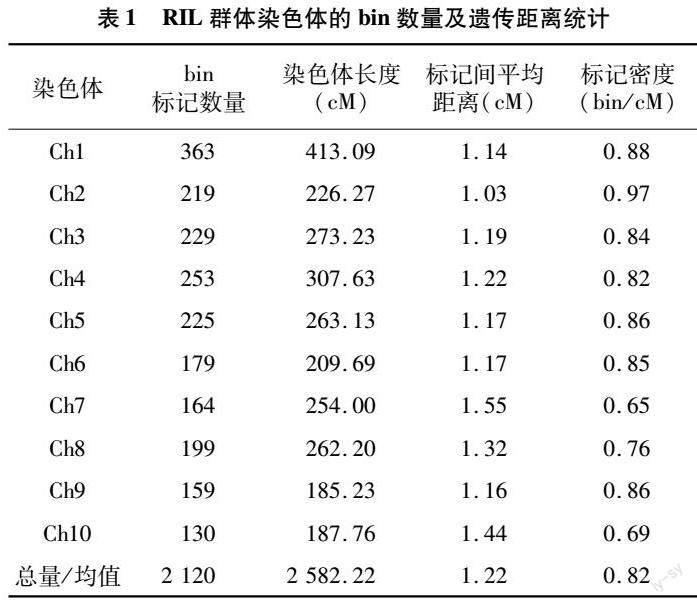
利用10K的SNP标记检测RIL群体的基因型,将其中连续多个标记基因型结果相同的SNP作为一个bin,筛选后共得到2 120个bin(表1)。构建bin map遗传图谱,全长为2 582.22 cM,标记间平均距离1.22 cM,标记密度平均为0.82 bin/cM。bin标记的分布在各条染色体中并不均匀:1号染色体的遗传距离最长(413.09 cM),9号染色体的遗传距离最短(185.23 cM);标记密度最大在2号染色体,为0.97 bin/cM,7号染色体标记密度最小,为 0.65 bin/cM(图1-A)。以bin在各染色体上的位置信息为根据,计算bin两两之间的重组率(图1-B)。
2.2 RIL群体雄穗分枝数和雄穗主轴长的表型和相关性分析
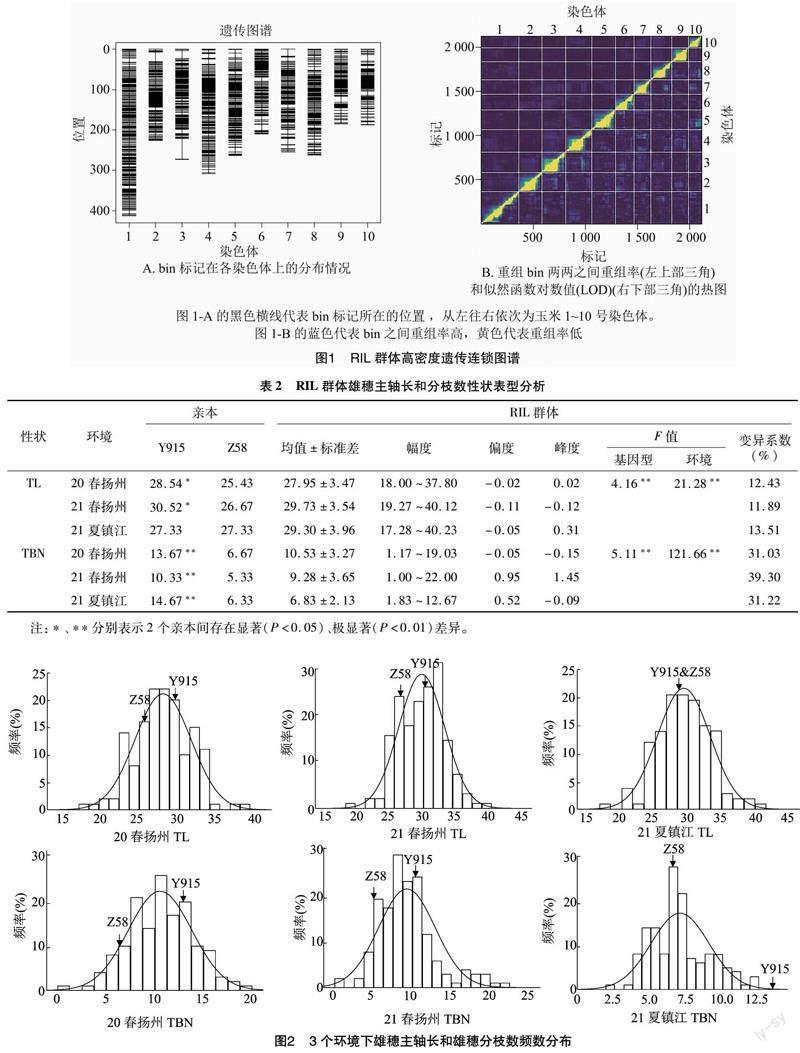
由表2可知,在3个环境中,2个亲本的雄穗主轴长和分枝数都有显著(P<0.05)或极显著(P<0.01)差异(21夏镇江的雄穗主轴长除外)。RIL群体除了分枝数在21春扬州峰度大于1外,群体的雄穗主轴长和分枝数在其他的环境中峰度和偏度的绝对值都小于1,近似正态分布(图2)。雄穗主轴长在3个环境下的变异系数分别是12.43%、11.89%和13.51%。雄穗分枝数在3个环境下的变异系数分别是31.03%、39.30%和31.22%。由基因型和环境的F值可知,2个性状在遗传差异和环境差异都表现为极显著水平。
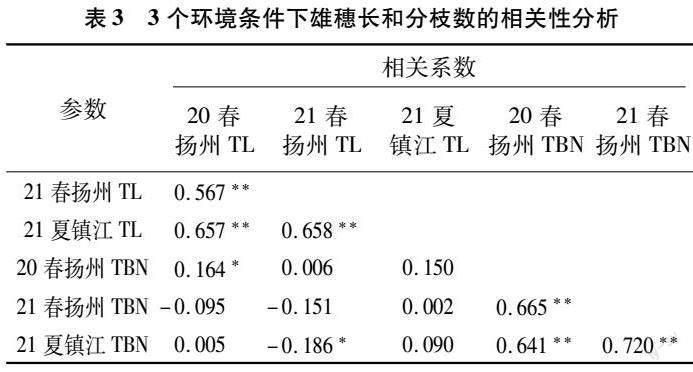
利用SPSS 21.0整合分析重组自交系群体3个环境下玉米RIL群体的雄穗主轴长和分枝数的表型值,结果(表3)表明,在3个环境下,雄穗主轴长之间呈极显著正相关关系,相关系数为0.567~0.658;分枝数两两之间也呈极显著正相关关系,相关系数为0.641~0.720。只有21春扬州雄穗主轴长和21夏镇江雄穗分枝数以及20春扬州雄穗主轴长和20春扬州雄穗分枝数之间存在显著相关关系,相关系数分别为-0.186和0.164,其余均无显著相关性。
2.3 玉米雄穗相关性状QTL定位
在3个环境下对雄穗主轴长和雄穗分枝数进行QTL定位,结果如表4和图3所示。4个雄穗分枝数QTLs分别位于1、4、6号染色体上,LOD值的范围为2.92~4.01,表型变异贡献率为6.34%~10.56%,其中qTBN1-1表型贡献率最大,达到10.56%,加性效应为1.34。雄穗主轴长的QTL位点共有6个,分别位于1、8、10号染色体,LOD值最大为5.21,最小为2.59,表型变异贡献率为5.10%~14.63%。经过分析发现,在21夏镇江环境下定位到的qTL1-3提供了最大的表型贡献率,达到14.63%,加性效应为-1.56。
在20春扬州和21春扬州定位到了同一个控制雄穗分枝数的位点qTBN1-1,2个环境下可解释的表型变异分别是10.56%和8.32%。并且此区间内在21春扬州检测到1个控制雄穗主轴长的位点qTL1-1,解释了8.73%的表型变异。
3 讨论
玉米育种目标之一是选育合适的玉米雄穗表型的品种,即保证在有足够粉量的同时尽量减小雄穗大小[18]。前人研究发现,雄穗分枝数与产量负相关,与雄穗大小正相关,是衡量雄穗大小的重要指标之一[19],但雄穗主轴长与分枝数的相关性却罕见报道。本研究利用Y915和郑58构成的RIL群体计算了雄穗主轴长和分枝数的相关性,大多数环境下主轴长和分枝数之间并无显著相关(表3),说明雄穗主轴长和分枝数可能是2个独立的影响雄穗大小的指标。
本研究在3个环境中共检测到10個雄穗主轴长和分枝数相关的QTL位点,分布在1、4、6、8、10号染色体上。检测到的QTL位点部分与前人研究有重复性,其中玉米雄穗主轴长QTL有4个与高晶等的研究结果[20]一致。位于1号染色体上的qTL1-1和qTL1-3均在高晶等基于代换系CL108发现的qTL1-2区间重叠;8号染色体上的qTL8-1在高晶等基于代换系CL113发现的qTL8区间重叠;10号染色体上的qTL10-1则与高晶等基于代换系CL136发现的qTL10区间重叠。雄穗分枝数QTL定位结果有3个与前人研究结果一致。位于4号染色体上的qTBN4-1与代资举等发现的qTBN4位点[9-10]在同一个区段内;位于6号染色体上的qTBN6-1与白成银构建的齐319X黄早四群体于标记umc1656-umc1796内定位到雄穗分枝数区间[21]重叠;6号染色体上定位到的另外一个QTL位点qTBN6-2则缩小了高晶等在代换系CL95中定位到的位于标记Y6q120-phi299852之间的qTBN6-2位点[20]。与前人研究有相似的位点说明了本研究QTL检测结果是真实可靠的。玉米雄穗主轴长和分枝数在多个环境下很少有重叠位点,这可能是由于在不同的环境条件下雄穗发育状况不同,从而对位点的特异性产生了影响。
本研究中还存在qTBN1-1、qTL1-2和qTL8-2这3个前人未发现的QTL,其中qTBN1-1在2个环境中被检测到,且表型贡献率分别达到8.32%和10.56%,此QTL位点可能具有较高的育种利用价值。另外2个QTL位点qTL1-2和qTL8-2只在单环境下检测到,这2个QTL则需要在新的不同群体中来验证其有效性。
4 结论
本研究对雄穗主轴长和分枝数2个性状进行分析,共检测到10个QTLs,其中只有qTBN1-1被重复检测到,且置信区间有重叠。同时这个区段还定位到1个控制雄穗主轴长的位点qTL1-1,表明这里可能存在控制雄穗主轴长和雄穗分枝数的一因多效或者紧密连锁的基因,可能具有较高的育种利用价值。
参考文献:
[1]Nikolic A,Andjelkovic V,Dodig D,et al. Quantitative trait loci for yield and morphological traits in maize under drought stress[J]. ABI Genetika,2011,43(2):263-276.
[2]李建波. 玉米雄穗分枝相关主效基因ub4的精细定位和候选基因鉴定[D]. 沈阳:沈阳农业大学,2019.
[3] Lambert R J,Johnson R R. Leaf angle,tassel morphology,and the performance of maize Hybrids1[J]. Crop Science,1978,18(3):499-502.
[4]贾 波,谢庆春,蒋学祥,等. 玉米雄穗主要性状研究进展[J]. 安徽农业科学,2016,44(32):24-26.
[5]Upadyayula N,da Silva H S,Bohn M O,et al. Genetic and QTL analysis of maize tassel and ear inflorescence architecture[J]. Theoretical and Applied Genetics,2006,112(4):592-606.
[6]汤 华,严建兵,黄益勤,等. 玉米5个农艺性状的QTL定位[J]. 遗传学报,2005,32(2):203-209.
[7]杨钊钊,李永祥,劉 成,等. 基于多个相关群体的玉米雄穗相关性状QTL分析[J]. 作物学报,2012,38(8):1435-1442.
[8]高世斌,赵茂俊,兰 海,等. 玉米雄穗分枝数与主轴长的QTL鉴定[J]. 遗传,2007,29(8):1013-1017.
[9]代资举,王新涛,杨 青,等. 玉米雄穗分枝数主效QTL定位及qTBN5近等基因系构建[J]. 作物学报,2018,44(8):1127-1135.
[10]张 媛,蒋 锋,刘鹏飞,等. 甜玉米雄穗分枝数的QTL定位[J]. 湖北农业科学,2013,52(15):3492-3495.
[11]Guan H H,Chen X J,Wang K L,et al. Genetic variation in ZmPAT7 contributes to tassel branch number in maize[J]. International Journal of Molecular Sciences,2022,23(5):2586.
[12]王铁固,马 娟,张怀胜,等. 玉米雄穗主轴长度和分枝数的主基因+多基因遗传分析[J]. 核农学报,2012,26(2):280-287.
[13] Mock J J,Schuetz S H. Inheritance of tassel branch number in Maize1[J]. Crop Science,1974,14(6):885-888.
[14]陈俊宇,卢 峰,陈婉莹,等. 玉米生育期相关性状QTL定位[J]. 江苏农业科学,2022,50(8):63-68.
[15]刘欢欢,王 艳,陈婉莹,等. 玉米穗位叶快速叶绿素荧光参数及产量相关性状QTL定位[J]. 分子植物育种,2022,20(6):1872-1882.
[16]Wang D F,Portis A R. Increased sensitivity of oxidized large isoform of ribulose-1,5-bisphosphate carboxylase/oxygenase (rubisco) activase to ADP inhibition is due to an interaction between its carboxyl extension and nucleotide-binding pocket[J]. Journal of Biological Chemistry,2006,281(35):25241-25249.
[17]McCouch S R,Cho Y G,Yano M,et al. Report on QTL nomenclature[J]. Rice Genetics Newsletter,1997,14:11-13
[18]刘军霞,何小娟,刘婷婷. 不同环境下玉米雄穗分枝数和主轴长的QTL分析[J]. 分子植物育种,2018,16(10):3219-3226.
[19]陈正杰. 玉米雄穗分枝数目数量遗传分析及QTL定位[D]. 雅安:四川农业大学,2014.
[20]高 晶,李海灵,田 然,等. 玉米雄穗相关性状QTL分析[J]. 玉米科学,2022,30(2):35-41,50.
[21]白成银. 玉米雄穗分枝数的QTL定位[D]. 雅安:四川农业大学,2013.
猜你喜欢
儿童时代·快乐苗苗(2022年11期)2022-12-12
少儿科学周刊·儿童版(2021年21期)2021-12-11
今日农业(2021年10期)2021-11-27
今日农业(2021年6期)2021-11-27
今日农业(2021年14期)2021-11-25
今日农业(2021年14期)2021-10-14
汉字汉语研究(2021年4期)2021-03-09
幽默大师(2020年5期)2020-06-22
小天使·二年级语数英综合(2019年10期)2019-11-08
红领巾·萌芽(2015年3期)2015-04-16
