
长期施用餐厨堆肥对水稻生育期杂草群落的影响
2023-08-27 07:58:02宋鹏曹盼刘欣宇方昭林永锋田光明邵小明
江苏农业科学 2023年14期
宋鹏 曹盼 刘欣宇 方昭 林永锋 田光明 邵小明
摘要:为探究长期施用餐厨堆肥后稻田杂草群落的变化规律,评估杂草群落结构对水稻产量的影响。连续4年设置餐厨堆肥(KF)、鸡粪有机肥(CM)、化肥(CF)及对照(CK)4个处理,在2021年对水稻在不同生育期的杂草群落进行监测和取样,对杂草的种类、密度、生物量、多样性以及杂草群落与水稻产量间的关系进行分析。结果表明,长期施用餐厨堆肥、鸡粪有机肥、化肥均减少了水稻莎草科和阔叶类杂草的密度和生物量,施肥处理的灌浆期杂草总生物量从高到低依次为CM(24.45 g/m2)、KF(21.98 g/m2)和CF(21.79 g/m2),水稻产量从高到低为KF(8 626.50 kg/hm2)、CF(8 071.50 kg/hm2)、CM(7 083.00 kg/hm2);改变了杂草群落多样性,CF处理杂草群落的 Shannon-Wiener 指数和Pielou均匀度指数较KF和CM处理显著降低,Simpson优势度显著提高;杂草群落结构的NMDS分析结果表明,KF和CM处理的杂草群落与CK有更好的相似性,与化肥相比,施用有机肥对杂草群落结构的改变具有一定的缓冲作用。综合来看,长期施用餐厨堆肥有利于减少稻田杂草总量并维持较高的杂草群落多样性和均匀度,减少杂草造成的水稻产量损失,有利于杂草群落的长期稳定。
关键词:餐厨堆肥;稻田;杂草群落;生物多样性;水稻产量
中图分类号:S511.06文献标志码:A
文章编号:1002-1302(2023)14-0146-07
杂草通过与作物竞争光照、水分和土壤养分等影响作物产量[1],而杂草多样性在维持生态系统功能方面发挥着重要作用。杂草多样性不仅可以为农业害虫的天敌提供栖息环境,降低害虫的种群数量,还能保持水土流失和促进土壤养分循环[2]。多样化的杂草群落还有助于维持杂草间的竞争关系,降低杂草与作物的竞争和限制杂草形成优势物种,以减少杂草对作物产量的负面影响[3]。影响杂草多样性的因子很多,包括外源肥料投入、种植制度、田间管理等[4-5],一直是杂草管理中关注的热点。其中,不同施肥条件下土壤中的N、P以及有机质含量都会影响杂草对养分的吸收和杂草生物量,进而影响杂草群落多样性构成[6],如土壤高氮条件能明显增加杂草丰富度[7],增加土壤磷含量会提高杂草生物量、莎草科以及苋属杂草的优势度[8-9]。因此合理平衡施用N、P、K肥可以减少优势种数量,维持较高的杂草多样性[10]。
餐厨堆肥作为一种新型肥料,是居民、餐饮业等食品生产、加工、消费各环节中产生的废料和食物残余,經过堆肥化处理得到的有机肥料[11]。已有研究表明,餐厨堆肥可以在改善土壤养分的同时,保障水稻产量[12]。也有研究发现,施用餐厨垃圾作为原料制备的新型有机肥对稻田禾本科和阔叶杂草有抑制作用,平均除草率可以达到80%,显著增加水稻产量[13]。鉴于餐厨堆肥在废弃物资源化利用方面的重要作用,却少有施用餐厨堆肥后对稻田杂草群落变化的研究。因此,本研究通过对连续施用不同类型肥料的稻田杂草群落开展调查,对水稻主要生育期的杂草种类、密度、生物量以及杂草多样性进行比较,探究施用餐厨堆肥后稻田杂草的演替规律,并通过与水稻产量的联系,综合评价餐厨堆肥对稻田杂草群落的影响。
1 材料与方法
1.1 试验地概况
试验地位于江苏省苏州市临湖镇中国农业大学水稻生态农场临湖基地(31°12′N, 120°45′E)。该地处于北亚热带湿润性季风气候区,年均气温为 16 ℃,降水量为1 160 mm,是水稻适宜生长区。试验地土壤为长三角地区典型渗育型水稻土,土壤基本理化性质:pH值为6.23,电导率为0.118 mS/cm,土壤速效氮含量为153.42 mg/kg,速效磷含量为 28.19 mg/kg,速效钾含量为171.67 mg/kg。
1.2 试验设计
本试验自2018年开始,试验所用肥料为餐厨堆肥、鸡粪有机肥和化肥。其中,餐厨堆肥为餐厨垃圾生化一体机处理,经过条垛式好氧发酵充分腐熟的堆肥产品(氮磷钾养分质量比为129 ∶24 ∶20);鸡粪有机肥为购买的商品有机肥(氮磷钾养分质量比为160 ∶117 ∶214);化肥为当地农业部门测土配施的化肥(氮磷钾养分质量比为16 ∶12 ∶17)。试验为随机区组设计,设置3个施肥处理:餐厨堆肥(KF)、鸡粪有机肥(CM)和化肥(CF),以不施肥为对照(CK)。各肥料按照等氮量在种植前作基肥施用,每个处理3个重复,随机区组排列,小区面积为10 m×10 m。各小区用田埂隔开,埂宽为1 m,埂两边采用薄膜隔离小区,并设置单独进出水道防止窜水窜肥。试验小区在每年的6月到11月种植水稻,水稻品种为南粳46(Oryza sativa Nanjing 46),行间距为15 cm。该试验地只种植一季水稻,其他时间不种植其他作物。为探究长期施肥后杂草群落原始结构的差异,在维持基本田间管理的基础上不采取额外除草措施。
1.3 调查方法
2021年,在水稻的拔节期(8月25日)、扬花期(9月16日)、灌浆期(9月26日)和成熟期(10月26日)在每个小区采用5点取样法,收集1 m×1 m样方中的全部杂草,带回实验室记录样方内杂草的种类及数量,并将杂草按种类分装,剪除根系后在 105 ℃ 条件下杀青1 h,随后80 ℃条件下烘干24 h后称其质量,杂草的鉴定主要参考《中国植物志》[14]和“iplant植物智”网站。在水稻成熟期每小区选择5个1 m×1 m样方,记录样方中水稻穴数,收割后用于脱粒考种计算水稻产量。在上述4个采样时期用土钻采集每个小区的土壤表层(0~15 cm)样品5个,混匀、风干后过筛用于常规养分检测。土壤养分含量测定参考鲍士旦的方法[15],测定pH值、全碳、全氮、碱解氮、有效磷、速效钾含量。
1.4 数据处理与分析
杂草多样性指数使用Past3软件计算[16],包括物种丰富度(S)、Shannon Wiener多样性指数(H)、Simpson优势度指数(D)和Pielou均匀度(J)指数。土壤碳氮比用全碳含量除以全氮含量得到。利用R 4.0.5软件对杂草密度、杂草生物量以及杂草多样性指数进行单因素方差分析,用 “Vegan”包基于Bray-Curtis距离对不同施肥处理下的杂草群落进行非度量多维尺度分析(NMDS),用“adonis2”函数进行置换多元方差分析(PREMNOVA),检验不同施肥條件下的杂草群落结构差异显著性,并利用“ggplot2”包进行绘图。
2 结果与分析
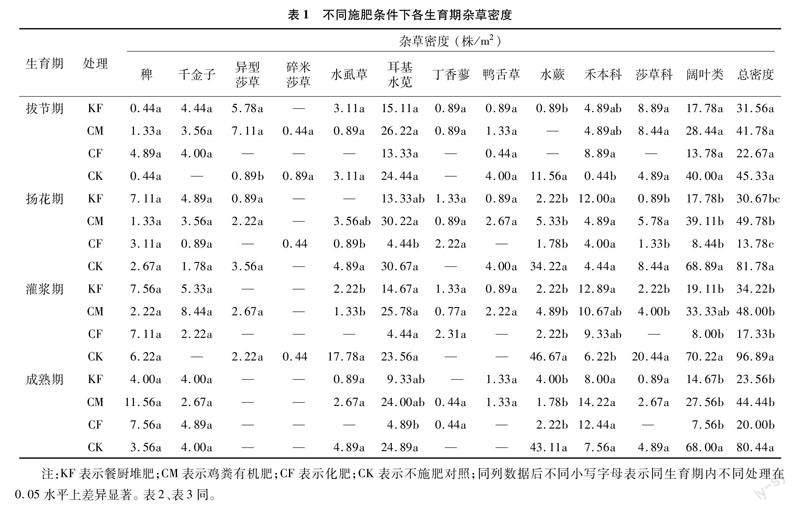
2.1 杂草密度和生物量
本试验在不同施肥处理的水稻4个生育期共发现9种杂草,分为禾本科、莎草科和阔叶类杂草三大类。与CK相比,施肥显著降低了除拔节期外的杂草总密度(表1),在水稻拔节期,各施肥处理的杂草总密度差异不显著,仅禾本科杂草密度在CK处理中显著少于CF处理,杂草总密度从大到小分别为CK(45.33株/m2)>CM(41.78株/m2)>KF(31.56株/m2)>CF(22.67株/m2);在水稻扬花期,与CK相比施肥显著降低了杂草总密度、阔叶类杂草的密度,其中CF处理的杂草总密度最低,为13.78株/m2,CK处理的杂草密度最高,为 81.78株/m2,与拔节期相比,CK处理的杂草总密度增加了80.4%;在水稻灌浆期,施肥增加了禾本科杂草密度,降低了杂草的总密度以及莎草科和阔叶类杂草密度,与CK相比KF处理显著增加了禾本科杂草尤其是千金子的密度,CF处理中无莎草科杂草被发现,与扬花期相比,CK处理的杂草总密度增加了18.48%;在水稻成熟期,与CK相比,施肥仅对阔叶类杂草密度影响显著,进而显著降低了杂草总密度。
从物种来看,与CK相比,施肥主要减少了水虱草(Fimbristylis littoralis)、耳基水苋(Ammannia arenaria)和水蕨(Ceratopteris thalictroides)的密度,从而使莎草科、阔叶类杂草减少,导致总杂草密度显著减少。随着水稻生育期的推进,密度上的差距逐渐扩大。各处理间,施肥处理的杂草总密度在各生育期较稳定,而CK处理的杂草总密度随生育期的推进而呈现整体增加趋势,在水稻成熟期有所下降。在施肥的3个处理中,与施用化肥(CF)相比,施用有机肥(KF、CM)增加了田间出现的杂草种类数,同时也增加了阔叶类杂草密度和杂草总密度,主要增加了耳基水苋的密度。在施用有机肥的处理中,与CM相比,KF降低了耳基水苋的密度,使杂草总密度减少。
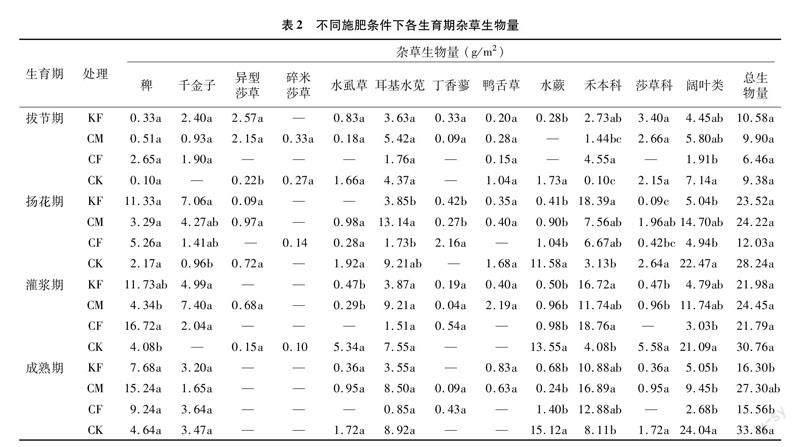
施肥整体明显增加了水稻各生育期禾本科杂草生物量、减少了莎草科和阔叶类杂草生物量,在杂草总生物量方面也有所减少(表2)。在水稻拔节期,禾本科杂草生物量从大到小为CF(4.55 g/m2)>KF(2.73 g/m2)>CM(1.44 g/m2)>CK(0.10 g/m2)。在水稻扬花期,施肥处理的禾本科杂草生物量,与CK相比,CF处理的杂草总生物量减少了57.40%,KF和CM分别减少了16.71%和14.24%;在水稻灌浆期,与CK相比,KF、CM和CF处理的莎草科杂草生物量减少了91.58%、82.80%和100%,杂草总生物量CF和KF分别减少了29.16%和28.54%;在水稻成熟期,与对照相比,施肥显著减少了阔叶类杂草生物量,同时增加了禾本科杂草生物量,CF和KF处理的杂草总生物量较CK显著减少,分别减少了54.05%和51.86%,CM处理减少了19.37%。
具体从物种水平看,与对照相比,施肥减少了水虱草和水蕨的生物量,增加了稗草生物量。与施用有机肥(KF、CM)相比,施用化肥(CF)使莎草科和阔叶类杂草生物量减少,在物种水平上主要是减少了耳基水苋的生物量。在施用有机肥的2个处理中,施用餐厨堆肥(KF)通过减少耳基水苋的生物量减少了杂草总生物量。
2.2 杂草多样性
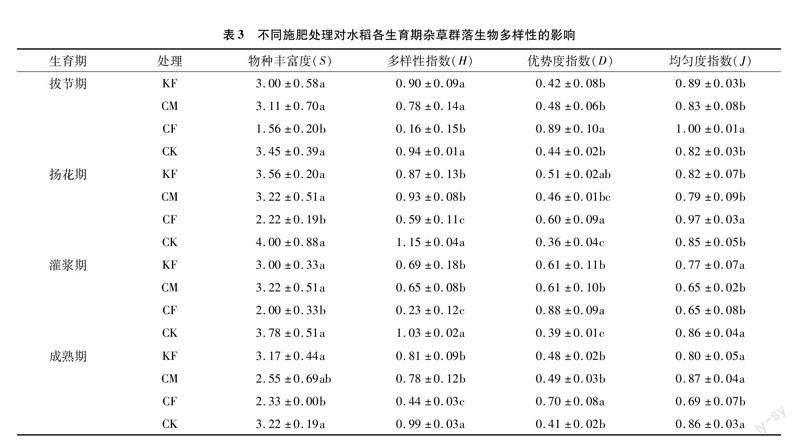
杂草密度和生物量在不同施肥条件下的变化促使各处理间杂草多样性产生差异。与CK相比,KF、CM和CF处理均减少了杂草群落的物种丰富度和Shannon-Wiener指数,其中CF处理降低效果显著,说明相较对照,施肥会减少杂草群落中出现的物种数目(表3)。同时,CF处理由于物种数目和多样性的降低使杂草群落的Simpson优势度提高,单一物种优势变大,表明施用化肥会影响杂草间的竞争能力,抑制部分杂草的生长。杂草群落Pielou均匀度指数方面,在水稻生长前期的拔节期和扬花期CF处理的均匀度指数显著高于其他处理,而随着生育期的推进而逐渐降低。CK处理的Pielou均匀度指数随着生育期推进而提高,说明CK处理中每种杂草的生物量较均匀,即每种杂草在与水稻竞争中都具有相同的竞争能力。
在物种水平上,造成CF处理与其他处理间群落多样性差异的原因是CF处理通过抑制碎米莎草、异型莎草和水虱草的生长使杂草群落的Shannon-Wiener指数下降,并且通过削弱耳基水苋和水蕨的竞争能力提高了群落Simpson优势度,促使杂草群落均匀度降低。而施用有机肥(KF、CM)时,杂草群落各物种组成均匀,多样性指数与CK相比差异不大。
2.3 杂草群落结构相似性及对环境因子的响应
NMDS图中处理间的距离表示各处理杂草群落结构的相似程度,通过对杂草群落的NMDS分析(图1),表明施肥改变了水稻各生育期的杂草群落结构。在水稻拔节期,施肥显著影响了杂草群落结构(PREMNOVA test P=0.015),CF处理与CK处理的距离最远,表明二者在杂草群落的组成和数量方面相差较大,KF和CM处理的原始距离较为接近,说明杂草群落的物种组成和数量较为相似。并且KF和CM与CF相比,前者与CK杂草群落的距离接近,表明相较于CF,KF和CM处理的杂草群落在物种组成和数量上与CK更为相似。在水稻扬花期,不同处理的杂草群落结构呈现明显分离(PREMNOVA test P=0.012),并且从距离上看,KF与CF的杂草群落相似度较高,CM和CK的相似度较高。水稻灌浆期展示了不同种类的杂草生物量和土壤养分对杂草群落结构的影响(图1-C),CF、KF和CM处理的杂草群落较为相似,而CK处理的杂草群落与其他3个处理沿莎草科和阔叶类杂草生物量增加方向明显分离,表明莎草科和阔叶类杂草生物量的差异是促使CK处理杂草群落与施肥处理不同的直接因素。由图1-C可知,表征土壤养分的碱解氮、有效磷和速效钾含量与莎草科和阔叶类杂草生物量之间呈负相关关系,表明施肥增加了土壤养分含量,导致杂草群落中禾本科、莎草科和阔叶类杂草组成和生物量的变化,间接引起杂草群落组成的差异。在水稻成熟期,各处理的杂草群落没有明显分离,表明在该时期,各处理的杂草群落相似度比较高,不同处理间杂草群落差异不显著(PREMNOVA test P=0.077)。
因此,与施用有机肥相比,施用化肥对杂草群落组成和数量的改变较大,与CF相比,KF和CM的杂草群落与CK有着更好的相似性,说明相比CF,KF和CM处理对杂草群落结构的改变具有缓冲作用,纵观整个生育期,长期施肥会显著改变拔节期、扬花期和灌浆期的杂草群落结构,而到了水稻成熟期,各处理杂草群落各组分的生物量又趋于相似。
2.4 杂草生物量和水稻产量的关系
不同施肥条件下水稻产量如图2所示,施用餐厨堆肥和化肥后水稻产量显著高于对照,施用餐厨堆肥、雞粪有机肥、化肥后水稻增产分别达53.23%、25.81%、43.37%,其中施用KF产量最高,为8 626.50 kg/hm2,较CM和CF分别增产21.79%、6.88%,但未达到显著水平。说明长期施用餐厨堆肥可以显著提升水稻产量,且效果优于鸡粪有机肥和化肥。
为探究不同杂草类群对水稻产量的响应,对各生育期的杂草类群生物量与水稻产量进行回归分析(表4)。结果表明,在水稻拔节期,禾本科杂草生物量与水稻产量具有显著的正相关关系,而莎草科和阔叶类杂草与水稻产量表现为不显著的负相关效应;水稻产量与扬花期莎草科和阔叶类杂草生物量表现为极显著的负相关关系,与禾本科杂草生物量为不显著的正相关关系;在水稻灌浆期,禾本科杂草生物量与水稻产量表现为极显著正相关,与莎草科和阔叶类杂草生物量表现为显著负相关关系;水稻产量与成熟期的禾本科和莎草科杂草生物量分别呈正相关和负相关,相关性不显著,与阔叶类杂草生物量呈显著负相关关系。这说明水稻的产量潜力越高,对于莎草科和阔叶类杂草的生长抑制作用就越强。
3 结论与讨论
农田生态系统中,施肥通过促进作物生长和使杂草适应土壤养分的规律性变化,改变杂草和作物的竞争关系,使杂草群落发生演替[1,9]。不同施肥方式对田间杂草总密度和生物量以及不同种类杂草的影响存在差异。有研究表明,长期施用猪粪有机肥和有机无机配施会增加稻田中禾本科杂草的密度,使杂草总密度和生物量增加[17-19];另有研究表明,长期施肥配合秸秆还田显著降低了莎草科和阔叶类杂草的密度和生物量,而对禾本科杂草密度仅有降低趋势[20]。在本研究中,施用的3种肥料均抑制了水稻各生育期莎草科和阔叶类杂草的生长,显著降低了莎草科以及阔叶类杂草的密度,同时减少二者以及总体的生物量,但不同肥料间仍存在差异。相比有机肥,施用化肥对莎草科和阔叶类杂草的抑制效果更好。与施用鸡粪有机肥相比,餐厨堆肥处理的杂草群落通过降低耳基水苋的发生数量,使阔叶类杂草的密度在水稻4个生育期分别降低了37.48%、54.54%、42.66%、46.77%,总杂草密度也均降低;在禾本科杂草方面,施肥明显提升了禾本科杂草生物量;而在密度方面,除水稻扬花期外,其他各生育期施肥均增加了禾本科杂草的密度。造成莎草科、阔叶类和禾本科杂草不同变化的原因可能是由于养分释放速率和不同种类杂草对光照的敏感度不同[10]。施肥促进了水稻快速生长,同时化肥的养分释放快,易使水稻迅速形成遮阴环境[21],抑制生长高度较矮的莎草科和阔叶类杂草的光合作用。而在对照中,由于水稻缺乏养分生长缓慢,不利于形成遮阴环境,使莎草科和阔叶类杂草大量生长,反过来抑制了同为禾本科的杂草和水稻的生长。
农田杂草多样性对于养分循环、保护天敌和控制害虫方面有重要作用,在农业生态系统中发挥着重要的生态功能[2]。杂草物种丰富度、多样性、优势度和均匀度指数可以从不同方面衡量杂草多样性,多样性指数反映了杂草群落物种数量、主要杂草的危害情况以及群落中每种杂草是否均匀,杂草种类越丰富,每种杂草越均匀,杂草群落的稳定性也就越高。施用不同肥料可以通过影响杂草群落中各种类型杂草的数量使杂草多样性指数发生改变。本研究结果表明,化肥处理的物种丰富度和多样性指数均较低,均匀度也随着生育期的推进逐渐降低,群落优势度较高,易使单一优势杂草演化为恶性杂草。施用有机肥处理维持了较高的物种丰富度和群落多样性指数,较低的群落优势度并在水稻生育后期维持较高的群落均匀度。说明施用有机肥有利于调节杂草的种间竞争,降低优势杂草演化为恶性杂草的可能,利于维持稳定的农田生态系统[18,22-23]。
降低杂草总量同时提高杂草多样性和均匀度对水稻增产有积极作用。本试验中,与对照相比,施用3种肥料通过提升土壤养分含量增加水稻养分吸收,显著提升了水稻产量,并通过增强水稻对杂草的竞争能力,降低了莎草科、阔叶类杂草以及总杂草生物量。本试验中施肥处理禾本科杂草生物量的增加,是出现禾本科杂草生物量与水稻产量呈正相关的原因,不代表禾本科杂草的增加会促使水稻增产,这可能是由于施肥处理中莎草科和阔叶类杂草减少,使其对禾本科杂草的竞争作用减弱,使禾本科杂草生物量增加。在水稻产量方面,施用餐厨堆肥略高于化肥处理,而杂草总生物量方面化肥处理杂草生物量小于餐厨堆肥,一般来说,杂草生物量越高对水稻产量的影响越大,本试验中可能是由于餐厨堆肥处理的杂草群落的多样性、均匀度提高,通过杂草性状的多样性限制了杂草与作物间的生态位重叠,减少了杂草与水稻的竞争,降低了杂草总量对水稻产量的负面影响[3,24]。综合来看,施用餐厨堆肥提升水稻产量的机制是通过减少群落中的杂草总量,改变禾本科、莎草科和阔叶类杂草三者的相对含量,调节群落结构,降低杂草与水稻间的竞争能力,减少杂草对水稻产量的负面效果。相关研究也认为减少杂草生物量同时增加杂草群落均匀度可以减轻作物的产量损失[3],这说明多样化的杂草群落限制了竞争性和优势物种对作物生产力的负面影响,同时可能促进杂草多样化提供的生态系统服务。
长期施用餐厨堆肥可以减少稻田中莎草科和阔叶类杂草的密度和生物量,使杂草总密度和生物量降低。其中,在水稻灌浆期杂草总密度和生物量相比对照分别减少了64.68%、28.54%。施肥显著改变了水稻各生育期的杂草群落结构,餐厨堆肥可以在降低杂草总密度和生物量的同时增加杂草群落多样性和均匀度,维持杂草群落的多样性和稳定性。与施用化肥相比,施用餐厨堆肥的杂草群落多样性和均匀度显著提高。长期施用餐厨堆肥可以通过改变水稻和杂草的竞争关系,降低莎草科和阔叶类杂草生物量,平衡不同种类杂草间的竞争关系,减少杂草导致的水稻产量损失。与化肥相比,餐厨堆肥处理中禾本科、莎草科和阔叶类杂草三者占比更均匀,杂草群落组成更稳定,并且在水稻产量方面相比化肥提高了6.88%,有较好的生态效益和经济效益。
参考文献:
[1]Tshewang S,Sindel B M,Ghimiray M,et al. Weed management challenges in rice (Oryza sativa L.) for food security in Bhutan:a review[J]. Crop Protection,2016,90:117-124.
[2]陈 欣,王兆骞,唐建军. 农业生态系统杂草多样性保持的生态学功能[J]. 生态学杂志,2000,19(4):50-52.
[3]Adeux G,Vieren E,Carlesi S,et al. Mitigating crop yield losses through weed diversity[J]. Nature Sustainability,2019,2(11):1018-1026.
[4]李照全,尹力初,周卫军,等. 农田管理措施对红壤稻田系统杂草种群结构的影响[J]. 农业现代化研究,2008,29(2):239-241,245.
[5]赵玉信,杨惠敏. 作物格局、土壤耕作和水肥管理对农田杂草发生的影响及其调控机制[J]. 草业学报,2015,24(8):199-210.
[6]Blackshaw R E ,Molnar L J ,Larney F J . Fertilizer, manure and compost effects on weed growth and competition with winter wheat in western Canada[J]. Crop Protection,2005,24(11):971-980.
[7]Ryan M R,Smith R G,Mirsky S B,et al. Management filters and species traits:weed community assembly in long-term organic and conventional systems[J]. Weed Science,2010,58(3):265-277.
[8]趙 锋,董文军,芮雯奕,等. 不同施肥模式对南方红壤稻田冬春杂草群落特征的影响[J]. 杂草科学,2009,27(1):7-12.
[9]蒋 敏,黄年生,张小祥,等. 施肥对农田杂草生理生态及群落形成的影响综述[J]. 杂草学报,2016,34(4):1-6.
[10]潘俊峰,万开元,李祖章,等. 施肥模式对晚稻田杂草群落的影响[J]. 植物营养与肥料学报,2015,21(1):200-210.
[11]Ren Y Y,Yu M,Wu C F,et al. A comprehensive review on food waste anaerobic digestion:research updates and tendencies[J]. Bioresource Technology,2018,247:1069-1076.
[12]满吉勇,林永锋,李丛林,等. 餐厨垃圾堆肥对水稻生长、产量及土壤养分含量的影响[J]. 中国农业大学学报,2021,26(11):165-179.
[13]Li Z R. The weeds control of a novel bioorganic fertilizer and its effects on agronomic traits of rice[J]. International Journal of Agriculture and Biology,2018,20(3):507-512.
[14]中国科学院中国植物志编辑委员会. 中国植物志:第十八卷[M]. 北京:科学出版社,1999.
[15]鲍士旦. 土壤农化分析[M]. 3版.北京:中国农业出版社,2000:40-114.
[16]Hammer-Muntz O,Harper D,Ryan P. PAST:paleontological statistics software package for education and data analysis[J]. Palaeontologia Electronica,2001,4(1):1-9.
[17]孙金秋,任相亮,胡红岩,等. 农田杂草群落演替的影响因素综述[J]. 杂草学报,2019,37(2):1-9.
[18]李昌新,赵 锋,芮雯奕,等. 长期秸秆还田和有机肥施用对双季稻田冬春季杂草群落的影响[J]. 草业学报,2009,18(3):142-147.
[19]董春华,曾希柏,文石林,等. 长期施肥对红壤双季稻冬闲田春季杂草群落的影响[J]. 中国生态农业学报,2015,23(9):1150-1157.
[20]张海艳,朱荣松,孙国俊,等. 不同施肥方式对稻麦轮作制稻田杂草群落的影响[J]. 杂草学报,2016,34(1):50-55.
[21]Yin L C,Cai Z C,Zhong W H. Changes in weed community diversity of maize crops due to long-term fertilization[J]. Crop Protection,2006,25(9):910-914.
[22]李儒海,强 胜,邱多生,等. 长期不同施肥方式对稻油轮作制水稻田杂草群落的影响[J]. 生态学报,2008,28(7):3236-3243.
[23]汤雷雷,万开元,陈 防. 养分管理与农田杂草生物多样性和遗传进化的关系研究进展[J]. 生态环境学报,2010,19(7):1744-1749.
[24]Navas M L. Trait-based approaches to unravelling the assembly of weed communities and their impact on agro-ecosystem functioning[J]. Weed Research,2012,52(6):479-488.
收稿日期:2022-09-30
基金项目:国家重点研发计划(编号:2016YFD0800605);江苏省苏州市农业科技创新项目(编号:SNG2020041、SNG2021012)。
作者简介:宋 鹏(1998—),男,山东日照人,硕士研究生,从事稻田杂草多样性和废弃物资源化研究。E-mail:songp@cau.edu.cn。
通信作者:邵小明,博士,教授,研究方向为植物分类和生态学。E-mail:shaoxm@cau.edu.cn。
猜你喜欢
趣味(作文与阅读)(2021年5期)2021-08-19 02:48:54
今日农业(2020年21期)2020-12-19 13:52:28
作文大王·低年级(2019年2期)2019-01-23 11:35:18
创新作文(小学版)(2018年19期)2018-11-30 01:56:08
江苏农业科学(2016年8期)2017-02-15 19:25:09
江苏农业科学(2016年8期)2017-02-15 19:24:57
环球人文地理·评论版(2016年5期)2017-01-03 03:19:00
农家科技下旬刊(2016年9期)2016-12-15 13:55:30
农业与技术(2016年20期)2016-12-08 21:26:55
农业与技术(2016年15期)2016-11-09 16:57:47
