
江苏省淮南地区主要小麦品种产量结构及Fhb1基因研究
2023-08-26 04:51李明星滕志英解小林王祝彩张晓慧陈春
安徽农业科学 2023年15期
李明星 滕志英 解小林 王祝彩 张晓慧 陈春
摘要 通过对江苏省淮南地区近年来审定的36个小麦品种进行试验示范,检测各品种是否携带Fhb1基因,旨在为优良品种推广及抗赤霉病基因Fhb1利用提供重要参考。结果表明,华麦12、华麦11、扬麦36、华麦8号、华麦13、宁麦资518、扬辐麦15等品种产量高,10个品种携带Fhb1基因,其中华麦8号和宁麦资518产量高、生育期短且携带Fhb1基因。
关键词 小麦;试验示范;赤霉病;Fhb1
中图分类号 S 512.1文獻标识码 A文章编号 0517-6611(2023)15-0025-04
doi:10.3969/j.issn.0517-6611.2023.15.007
Yield Structure and Fhb1 Resistance Gene of Main Wheat Varieties in Huainan Area of Jiangsu Province
LI Ming-xing,TENG Zhi-ying,XIE Xiao-lin et al
(Breeding Research Institute,Jiangsu Dahua Seed Enterprise Co.,Ltd.,Huaian,Jiangsu 223216)
Abstract A total of 36 wheat varieties were used for demonstrated experiment in Huainan Area of Jiangsu Province in recent years.Whether the varieties carried Fhb1 gene was tested,so as to provide important references for the promotion of excellent varieties and the utilization of Fhb1 gene for resistance to fusarium head blight.Results showed that Huamai 12,Huamai 11,Yangmai 36,Huamai 8,Huamai 13,Ningmaizi 518,Yangfumai 15 and other varieties had high yields,and 10 varieties carried the Fhb1 gene.Among them,Huamai 8,Ningmaizi 518 had high yield,short growth period and carried Fhb1 gene.
Key words Wheat;Experiment demonstration;Fusarium head blight;Fhb1
小麦(Triticum spp.L.)是世界上最重要的大田作物之一,是世界上很大一部分人口的主食。江苏省淮南地区属于长江中下游平原冬麦区,主要种植春性小麦品种,在江苏省粮食生产中扮演重要作用。高产、多抗的小麦品种便于在大田生产种植;小麦生产受到多种制约因素的影响,赤霉病是小麦生产的主要生物制约因素之一。小麦赤霉病(fusarium head blight,FHB)是由禾谷镰刀菌(Fusarium graminearum)等多种镰刀菌引起的一种全球性真菌病害,是造成我国小麦减产的罪魁祸首[1]。赤霉病不仅会造成籽粒萎缩,甚至成种失败,还会通过感染霉菌毒素使粮食品质恶化。脱氧雪腐镰刀菌烯醇(deoxynivalenol,DON)是由禾谷镰刀菌产生的霉菌毒素,同时也是一种致病因子,在非常低的浓度下对动物具有高度毒性,会导致恶心、呕吐、腹泻、腹痛、头痛、头晕、发烧等症状,如果接触足够多,可引起癌变,甚至会导致死亡[2]。在小麦的21条染色体上都发现了涉及赤霉病抗性的QTL,其中2个最强和最有效的是Fhb1和Qfhs.ifa-5A,均来源于“苏麦3号”[3]。Fhb1是小麦和其他小粒粮食作物中对赤霉病具有强II型抗性的主要效应QTL[4]。鉴于此,笔者通过对江苏省淮南地区近年来审定的36个小麦品种进行试验示范,检测各品种是否携带Fhb1基因,旨在为优良品种推广及抗赤霉病基因Fhb1利用提供重要参考。
1 材料与方法
1.1 试验地概况 试验在白马湖农场农科所(119.1°E,33.2°N)进行,各品种种植0.067 hm2,海拔7 m,苏北灌溉总渠旁;交通便利,地势平坦,排灌方便;田块为黏壤土,地力中等,土壤肥力均匀,前茬作物为水稻。
1.2 供试材料
江苏省淮南地区近年来审定的主要品种包括宁麦29、苏麦0588、苏麦288、隆麦39、扬辐麦10号、泰麦902、泰麦901、金丰麦1号、扬麦31、扬麦37、扬麦36、宁麦33、明麦133、扬麦33、明麦13、扬麦34、镇麦13、瑞华麦596、农麦99、农麦156、华麦8号、苏研麦017、扬辐麦15、宁麦资518、宁麦资166、华麦11号、华麦12号、江丰麦1号、宁麦资218、扬江麦586、扬辐麦9号、盐麦1号、镇麦18、中垦麦212、中垦麦616、华麦13号,抗赤霉病基因研究对照品种苏麦3号、扬麦158。
1.3 试验方法
播种前根据千粒重、发芽率等计算播量,11月3日用播种机等行条播,行距25 cm,10月29日机械普施复合肥600 kg/hm2,12月8日无人机施尿素150 kg/hm2,3月15日无人机施尿素225 kg/hm2,一生施纯氮26.5 kg/hm2。其他田间管理同本地区大田生产。
1.4 性状调查记载测定
小麦生长的全生育期内调查记载生育期、茎蘖动态、穗数、穗粒数等主要农艺性状;全区收获后及时晾晒风干,测量产量、千粒重等。
1.5 抗赤霉病基因Fhb1检测
1.5.1 小麦基因组DNA提取。
当小麦生长到分蘖拔节期时,取小麦的幼嫩叶片,放入-20 ℃储藏备用,用CTAB法提取小麦基因组DNA[5]。
1.5.2 赤霉病基因Fhb1扩增及标记检测。
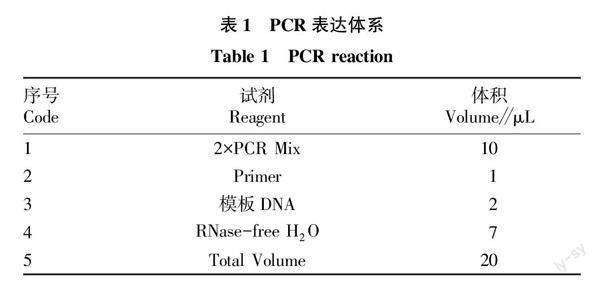
用与Fhb1基因连锁的标记引物His3B-4[6]进行赤霉病抗性检测。引物由上海生物工程技术公司合成。反应体系如表1所示,2×PCR Mix 包含0.1 U/μL Taq酶、500 μ mol/L dNTPs、20 mmol/L Tri-HCl、10 mmol/L KCl、3 mmol/L MgCl2。
PCR扩增程序:95 ℃ 预变性5 min;95 ℃ 变性5 s,58 ℃ 退火30 s,72 ℃ 延伸90 s,共35个循环,再72 ℃聚合延伸10 min,12 ℃保存。PCR反应结束后,得到扩增产物。扩增产物加入1.5%的琼脂糖凝胶电泳中检测,电泳完毕后放入EB中染色,然后用紫外凝胶成像仪拍照。
2 结果与分析
2.1 不同品种生育期及茎蘖动态比较
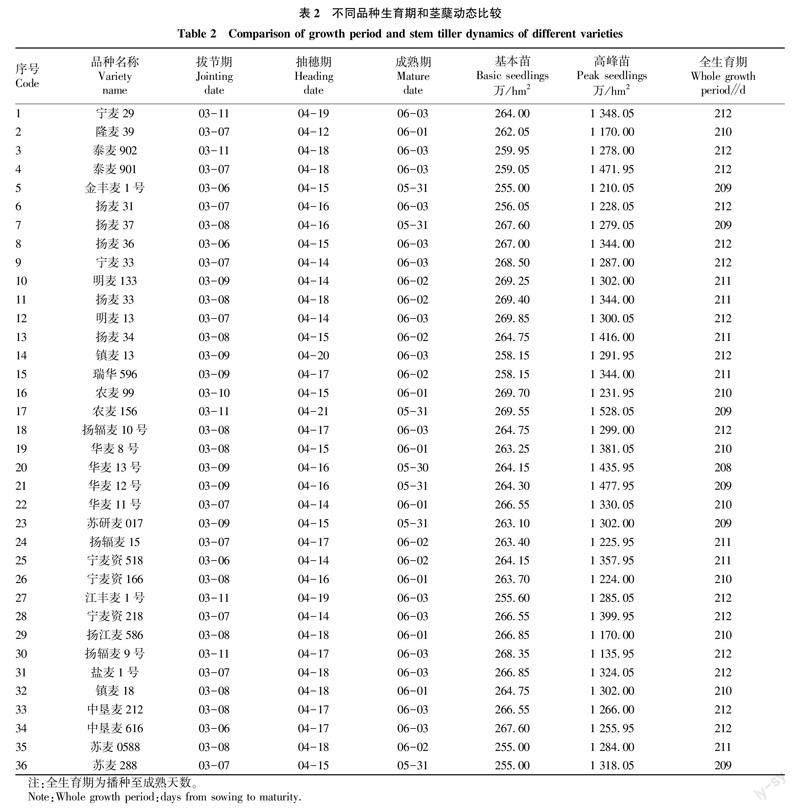
由表2可知,不同品种拔节期在3月6—11日,抽穗期在4月12—21日,成熟期在5月30日—6月3日,全生育期在208~212 d。基本苗在255.00万~269.85万/hm2,高峰苗在1 135.95万~1 528.05万/hm2。
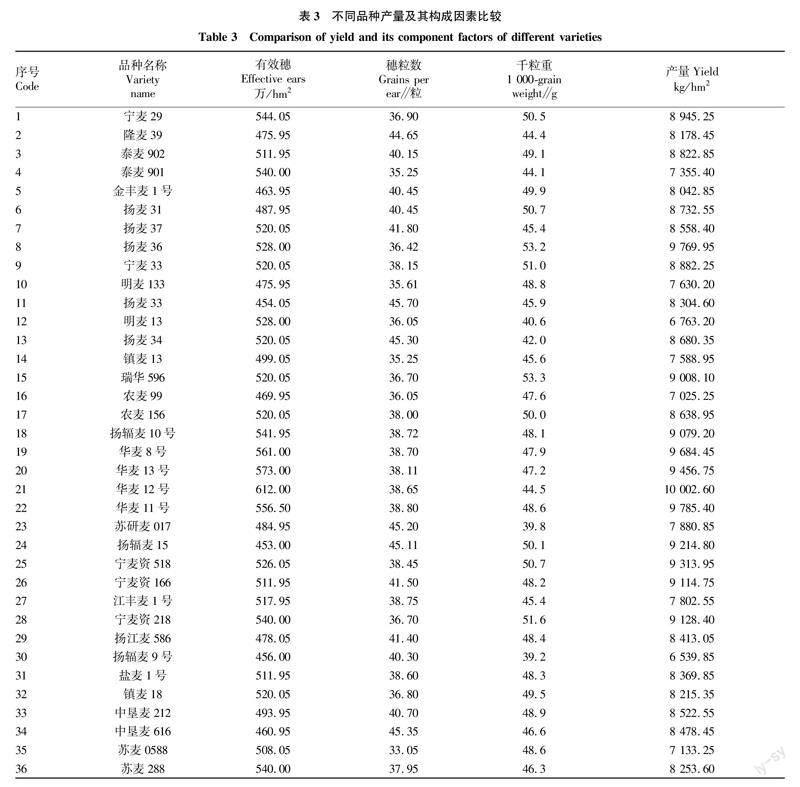
2.2 不同品种产量及其构成因素比较 由表3可知,不同品种有效穗在453.00万~612.00万穗/hm2,其中扬辐麦15最少,华麦12号最多;穗粒数在33.05~45.70粒/穗,其中苏麦0588最少,扬麦33最多;千粒重在39.2~53.3 g,其中扬辐麦9号最低,瑞华596最高;产量在6 539.85~10 002.60 kg/hm2,其中扬辐麦9号最低,华麦12号最高。
2.3 抗赤霉病基因Fhb1分子检测
利用His3B-4标记引物对各品种的Fhb1基因进行检测,如图1所示,检测出携带纯合Fhb1基因品种有华麦8号、扬麦33、镇麦13、泰麦901、苏麦0588、宁麦33、扬辐麦9号;携带杂合Fhb1基因品种有宁麦资166、泰麦902、扬麦28、宁麦资518;其他品种不含Fhb1基因。
3 结论与讨论
小麦作为我国重要的口粮作物,选育高产、多抗的优质品种,保障国家粮食安全具有重要作用[7],近年来江苏省淮南地区赤霉病发病趋势逐年过大,严重威胁小麦的产量品质等特性。现阶段赤霉病防治主要运用化学防治、农业防治和生物防治等方法。长年使用化学防治会使赤霉病菌产生抗性,且已发现病原菌对戊唑醇和苯丙咪唑类杀菌剂产生抗性[8]。农业防治主要是通过栽培措施和品种遗传改良来降低赤霉病为害程度,通过轮作、灭茬和排水减湿等栽培措施防治赤霉病效果十分有限[9]。由于赤霉病抗性是数量遗传,受环境的影响较大,抗性机理复杂,通过品种杂交改良的方法培育高抗品种目前难度极大[10]。改善小麦赤霉病遗传抗病性是培育抗赤霉病品种不可或缺的一部分。该研究通过对江苏省淮南地区近年来审定的主要小麦品种在2021—2022年进行大面积示范展示,筛选出华麦12号、华麦11号、扬麦36、华麦8号、华麦13号、宁麦资518、扬辐麦15产量超9 150.00 kg/hm2,同时通过分子标记检测到携带赤霉病抗性基因Fhb1的品种10个,其中华麦8号和宁麦资518产量高、生育期短且携带Fhb1基因,为高产优质品种大面积生产推广及Fhb1基因利用提供重要依据。
参考文献
[1] YIN Y N,LIU X,LI B,et al.Characterization of sterol demethylation inhibitor-resistant isolates of Fusarium asiaticum and F.gra minearum collected from wheat in China[J].Phytopathology,2009,99(5):487-497.
[2] MCMULLEN M,BERGSTROM G,DE WOLF E,et al.A unified effort to fight an enemy of wheat and barley: Fusarium head blight[J].Plant disease,2012,96(12):1712-1728.
[3] AGOSTINELLI A M,CLARK A J,BROWN-GUEDIRA G,et al.Optimizing phenotypic and genotypic selection for Fusarium head blight resistance in wheat[J].Euphytica,2012,186(1):115-126.
[4] STEINER B,BUERSTMAYR M,WAGNER C,et al.Fine-mapping of the Fusarium head blight resistance QTL Qfhs.ifa-5A identifies two resistance QTL associated with anther extrusion[J].Theoretical and applied genetics,2019,132(7):2039-2053.
[5] WANG M L,BARKLEY N A,YU J K,et al.Transfer of simple sequence repeat (SSR) markers from major cereal crops to minor grass species for germplasm characterization and evaluation[J].Plant genetic resources,2005,3(1):45-57.
[6] 朱展望,徐登安,程顺和,等.中国小麦品种抗赤霉病基因Fhb1的鉴定与溯源[J].作物学报,2018,44(4):473-482.
[7] 许娟,陈莉,丁克坚.安徽省小麦品种对赤霉病的抗性分析[J].安徽农业科学,2019,47(16):172-173,181.
[8] SPOLTI P,DEL PONTE E M,DONG Y H,et al.Triazole sensitivity in a contemporary population of Fusarium graminearum from New York wheat and competitiveness of a tebuconazole-resistant isolate[J].Plant disease,2014,98(5):607-613.
[9] 刘东军,宋维富,杨雪峰,等.小麦Fhb1基因定位、克隆及其在抗赤霉病育种中利用的研究进展[J].作物杂志,2020(4):16-20.
[10] BUERSTMAYR H,BAN T,ANDERSON J A.QTL mapping and marker-assisted selection for Fusarium head blight resistance in wheat:A review[J].Plant breeding,2009,128:1-26.
基金項目 江苏省农垦农业发展股份有限公司农业科技项目(NK202101)。
作者简介 李明星(1992—),男,江苏淮安人,助理农艺师,硕士,从事稻麦育种及农作物栽培研究。
收稿日期 2022-08-11
猜你喜欢
今日农业(2022年16期)2022-11-09
天津农林科技(2022年2期)2022-04-19
云南化工(2021年9期)2021-12-21
今日农业(2021年12期)2021-11-28
今日农业(2021年8期)2021-11-28
今日农业(2021年9期)2021-11-26
金桥(2021年10期)2021-11-05
今日农业(2021年13期)2021-08-14
今日农业(2021年9期)2021-07-28
作文小学中年级(2020年4期)2020-06-11
