
广西君武森林公园海南繁殖生态和巢址选择
2023-08-16 13:02谭丽凤曹和琴俸海刀李斌强蒋桂娟杨再玺粟海军
野生动物学报 2023年3期
谭丽凤,曹和琴,俸海刀,李斌强,蒋桂娟,杨再玺,粟海军
(1.广西生态工程职业技术学院,柳州,545004;2.贵州大学林学院,贵阳,550025;3.柳州市动物园管理处,柳州,545005)
动物行为是动物个体或群体适应内外环境变化时做出的反应,是对周围生物和非生物环境的动态适应[1]。在长期的自然选择中,鸟类繁殖行为表现出与环境相适应的繁殖策略。繁殖是鸟类生活史中极其关键的环节,影响着种群规模和物种延续,而繁殖成功与否则与巢址质量密切相关[2]。高质量的巢址能降低风险,为动物最大限度地提供食物和微栖息环境,提高繁殖成功率[3]。鸟类的巢址选择,是鸟类重要的繁殖行为对策[4]。因此,了解物种的繁殖生态学习性,以及它们如何选择适宜的巢址,对保护珍稀濒危物种至关重要[5]。
2013 年3 月—2022 年7 月,项目团队在君武森林公园对海南进行了连续近10 年的繁殖观察和记录,并调查繁殖生态和巢址特征,分析影响巢址选择的主要因素,以期为研究和保护海南提供更翔实的基础资料,对有效保护该物种有重要意义。
1 研究区概况
君武森林公园(24°27′21.59″—24°29′49.19″ N,109°20′16.8″—109°24′46.7″ E)位于广西柳州市北郊,面积338.8 hm2(图1)。公园地处南亚热带季风区,全年热量充足,夏热冬暖,年平均气温20.7 ℃;降水量充沛,年平均降水量1 535.6 mm。公园内地势平坦,植被覆盖率高达85.7%,树种以人工种植的马尾松(Pinus massoniana)、大叶栎(Quercus griffithii)和石栎(Lithocarpus glaber)为主,灌木层主要有大青(Clerodendrum cyrtophyllum)、杜茎山(Maesa japonica)、三桠苦(Melicope pteleifolia)和牛耳枫(Daphniphyllum calycinum)等,草本层主要有淡竹叶(Lophatherum gracile)、五节芒(Miscanthus floridulus)和扇叶铁线蕨(Adiantum flabellulatum)等(图2A、B)。
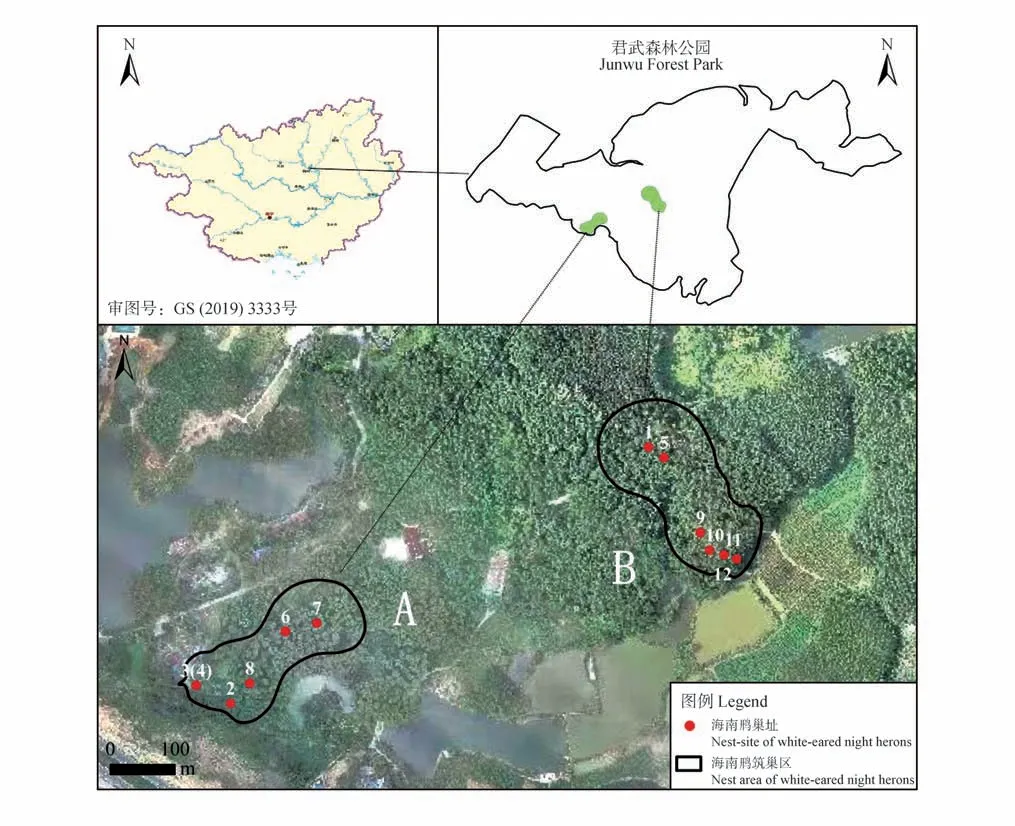
图1 研究区域及巢分布Fig.1 Study area and nest distribution

图2 君武森林公园海南繁殖记录Fig.2 The breeding record of white-eared night herons in Junwu Forest Park
2 研究方法
2013—2022 年,于每年2 月下旬开始,在君武森林公园内使用望远镜(西光航海家8×42、兴和BDⅡ8×42 等)或观察粪便痕迹等方法地毯式摸排寻找海南。发现繁殖巢后,使用GPS(GARMIN MAP60CSx 或集思宝G130BD)定位。采用年份 加顺序号命名,如2013 年发现的第1 个巢,命名为201301 号巢,以此类推。根据海南昼伏夜出的习性,采用定点观察法,于06:00—08:00、18:00—22:00 使用双筒望远镜和夜视仪(博士能260501)进行观察,并不定期做全天观察,直到最后一只幼鸟离开巢树。记录海南的个体数目、巢树树种、窝卵数、幼鸟数及繁殖各阶段的行为活动。
在发现繁殖巢时,以巢树为中心,设置1 个 10 m×10 m的样方,采用年份加顺序号命名,如2013年的第1 个巢样方,命名为201301 号样方。海南离巢后在保证不对其造成影响的情况下,快速测量样方内巢和巢址的相关数据(表1)。

表1 巢址选择的调查参数和测量方法Tab.1 The parameters of nest-site selection and sampling methods
3 数据处理
用SPSS 20.0 对巢和繁殖成效数据进行描述性特征分析,均值用平均值±标准差(mean±SD)表示。计算孵化率、幼鸟成活率和繁殖成功率。孵化率=孵化数/窝卵数×100%;幼鸟成活率=幼鸟成活数/孵化数×100%;繁殖成功率=幼鸟飞出巢数/总巢数×100%。
参考丁长青等[2]通过主成分分析法(PCA)对反映巢址主要特征的变量进行分析。选择特征值大于1的主成分进行巢址选择影响因素分析[14]。
4 结果
4.1 繁殖基本情况
在2013—2022 年,除2017 年和2020 年未记录到海南繁殖,其余年份共记录到12 个巢的繁殖情况。其中,2013—2016年观察到8个巢的窝卵数均为3 枚,共计24 枚,孵化出20 只雏鸟,孵化率83.33%;幼鸟成活18只,幼鸟成活率90.00%;有2个巢弃巢,6个巢有幼鸟成功飞出,繁殖成功率75.00%。2018—2022 年有4 个巢未观察到窝卵数,仅观察到每窝 出雏均为2 只,共8 只,幼鸟成活8 只,幼鸟成活率100.00%;4 个巢的幼鸟全部成功飞出,繁殖成功率100%。
整体而言,12 个巢共计出雏32 只,幼鸟成活 28 只,幼鸟成活率87.50%;2 个巢弃巢,无幼鸟飞出,其余10 个巢均有幼鸟成功飞出,总繁殖成功率为83.33%。
4.2 繁殖行为
4.2.1 筑巢与产卵、孵卵和出雏
拾到的卵壳中有2 枚带新鲜血丝。第1 枚于2014 年3 月31 日在201401 号巢下拾到,第2 枚于2014 年5 月5 日在201402 号巢下拾到,推测雏鸟刚出壳(表2)。海南孵化期为25~27 d[17,19],应用回溯方法来推算,201401号巢产卵时间为3月4—6日;201402号巢产卵时间为4月8—10日。
表2 2013—2022年君武森林公园海南育雏及幼鸟行为记录Tab.2 Records of brooding and behavior of young birds of white-eared night herons in Junwu Forest Park from 2013 to 2022
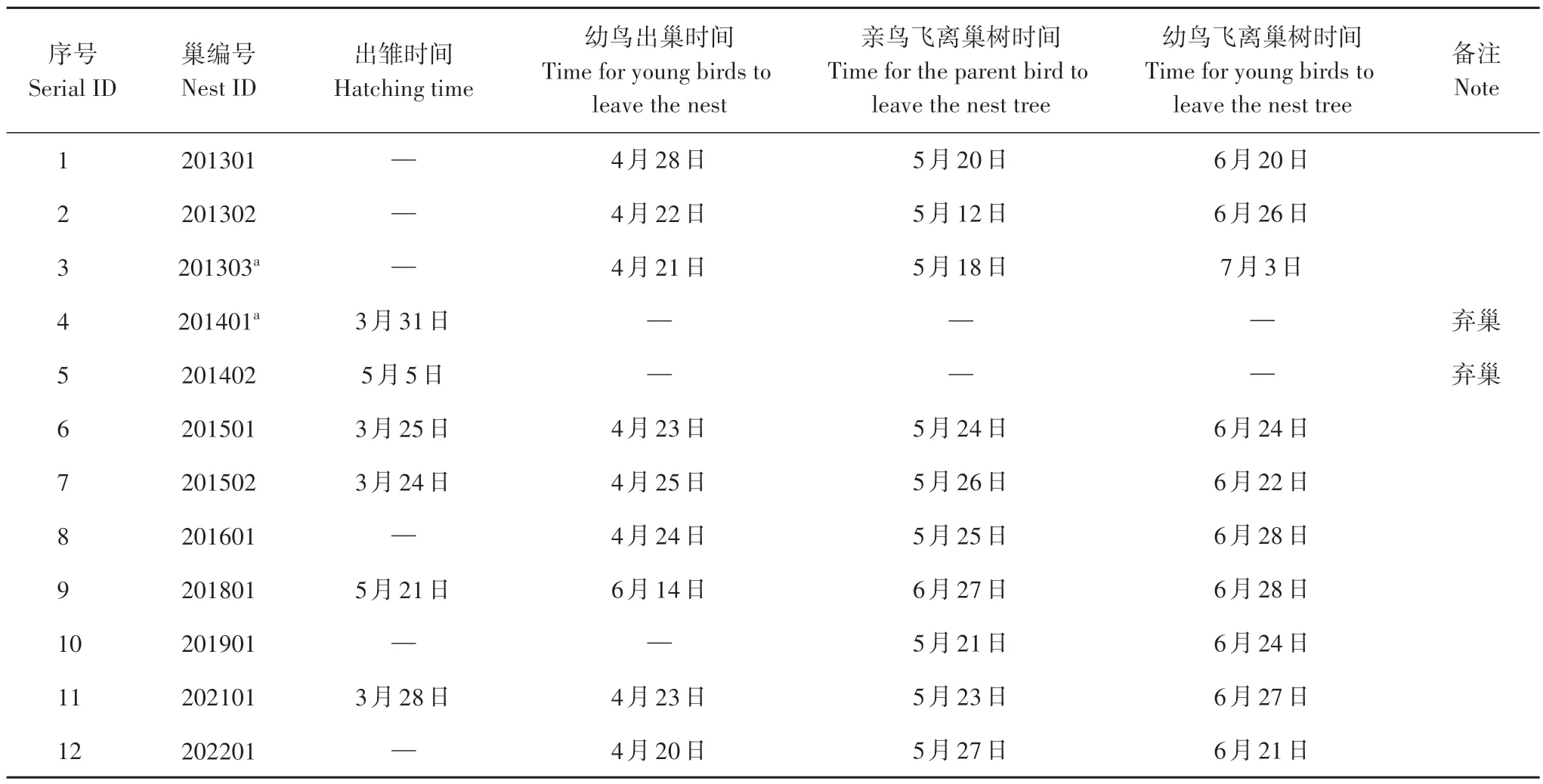
表2 2013—2022年君武森林公园海南育雏及幼鸟行为记录Tab.2 Records of brooding and behavior of young birds of white-eared night herons in Junwu Forest Park from 2013 to 2022
注:a.不同年份使用同一棵树,不同树枝Note:a,Using different branches of the same tree in different years
观察到亲鸟轮流孵卵。一只亲鸟在巢内孵卵时,另一只亲鸟站在巢旁的树枝上(图2D)。白天静栖不动,夜间轮流外出觅食。
4.2.2 育雏及幼鸟行为
雏鸟出壳后,仍趴在亲鸟身下取暖(图2E),亲鸟轮流外出觅食。若亲鸟相继飞出觅食,飞出的方向均不相同,可能是为了避免觅食区域重叠从而提高觅食效率。外出觅食开始时间为19:30 左右,记录的觅食最短时间为21 min。亲鸟飞回后,站在巢边等待2~5 min后才开始喂食。到育雏后期,观察到亲鸟10:30 仍外出觅食,可能与幼鸟快速生长需要更多能量有关。
共记录到9个巢的幼鸟出巢时间,其中8个巢的幼鸟出巢时间在4 月下旬,此时幼鸟的白色绒羽基本褪完,出巢后站到巢所在的树枝上(图2F)。白天基本站立不动,天色渐黑后开始练习拍打翅膀、跳跃等动作。随着成长,幼鸟逐渐练习飞行,离开巢树再返回的时长不等。直至6 月末—7 月初,未再观察到幼鸟回到巢树。另外,观察到的9个巢中,有8个巢的亲鸟于5月中下旬后在巢树上未再观察到(表2)。亲鸟离开巢树后是否在附近区域抚育幼鸟,幼鸟离巢后如何觅食及生存质量如何等还有待进一步研究。
4.3 巢和巢址特征
在2013—2022年,测量了12个巢样方的相关参数。巢距地面高(27.0±2.2)m(n=12)。由于巢距离地面过高,未测量到巢的大小。目测巢呈浅盘状,以马尾松的枯枝简单搭建。
巢树11 株,其中1 株连续2 年有巢(201303 号巢和201401 号巢)。巢树树种均为马尾松,树高(32.6±2.0)m(n=12),胸径(55.0±2.2)cm(n=12),距水边距离(111±60)m(n=12),距林缘距离(127±63)m(n=12),距道路距离(12±8)m(n=12)。
表3 2013—2022年君武森林公园海南繁殖巢址特征Tab.3 Nest characteristics of white-eared night herons in Junwu Forest Park from 2013 to 2022
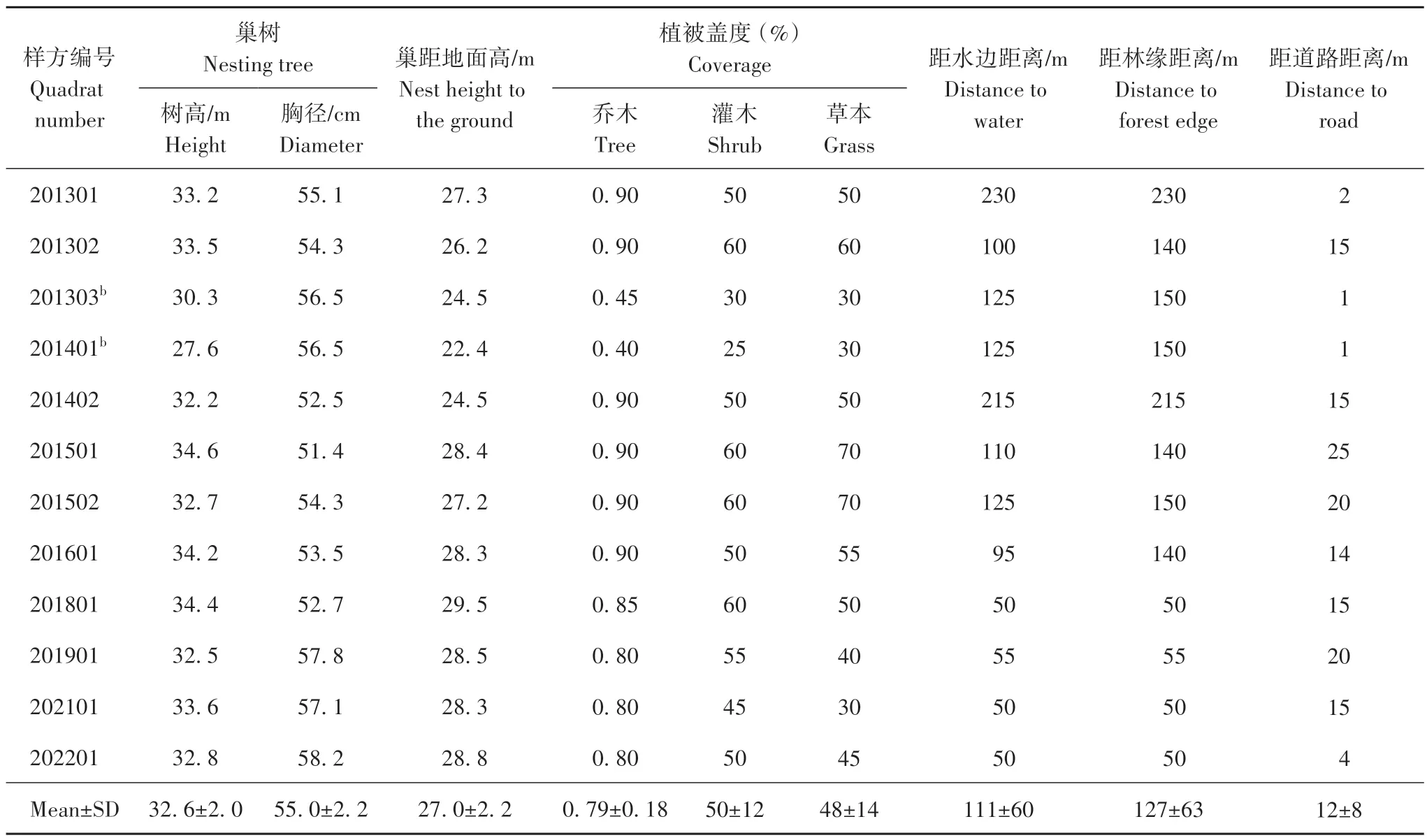
表3 2013—2022年君武森林公园海南繁殖巢址特征Tab.3 Nest characteristics of white-eared night herons in Junwu Forest Park from 2013 to 2022
注:b.为同一地理位置不同年份的巢样方Note:b,The nest sample square of the same geographical location in different years
通过对12 个巢的生境因子进行主成分分析,得到9个因子的相关矩阵和特征值(表4)。结果显示,前2个主成分的特征值大于1,其方差贡献率分别是55.00%和29.00%,累计方差贡献率达84.00%,可以较好地反映海南的巢址生境特征。因此提取前2个特征值,计算特征向量。
表4 海南巢址主成分分析结果Tab.4 The principal component analysis on nest-site characteristics of white-eared night herons
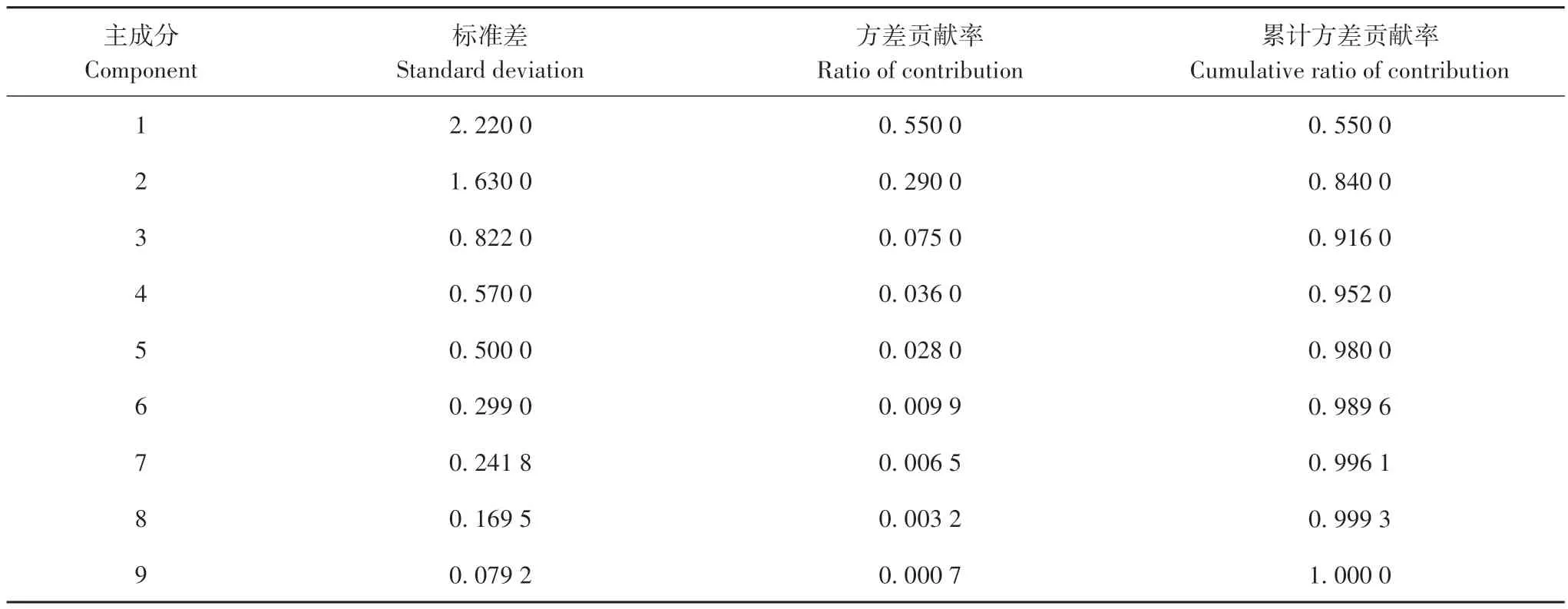
表4 海南巢址主成分分析结果Tab.4 The principal component analysis on nest-site characteristics of white-eared night herons
由表5 可知,第一主成分贡献率为55.00%,其中灌木盖度、巢树高和乔木郁闭度的因子载荷绝对值>0.4,这3 项与巢的隐蔽性有关,说明第一主成分是安全因素,且第一主成分与这3 个因子呈正相关。第二主成分贡献率为29.00%,其中,距林缘距离、距水边距离的因子载荷绝对值>0.5,在本研究中,林缘紧挨水塘,这2 项因子均与海南觅食的距离有关,因此归纳为食物因素,且第二主成分与这2 个因子呈负相关。
表5 海南巢址选择的主成分Tab.5 The principal component index matrix on nest-site characteristics of the white-eared night herons
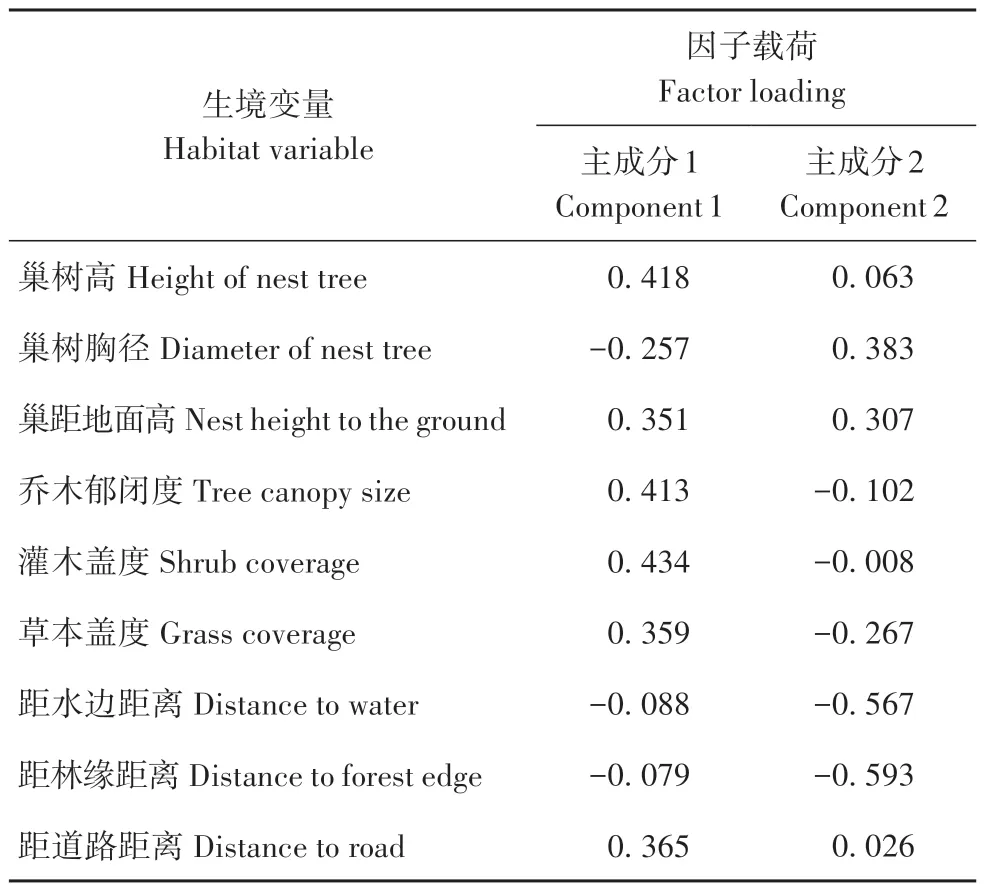
表5 海南巢址选择的主成分Tab.5 The principal component index matrix on nest-site characteristics of the white-eared night herons
5 讨论
5.1 繁殖行为
5.2 巢址选择
巢址选择是鸟类进化过程中形成的有利于种群繁衍的行为策略。在这个选择的过程中,鸟类更倾向于选择风险小、繁殖成效大的营巢生境[25]。在海南繁殖栖息地研究中,余丽江[16]发现栖息地质量(海拔、坡度、巢树高和巢距地面高)和隐蔽性(乔木盖度、巢树胸径和距水边距离)是影响海南巢址选择的主要因素;李必成[17]的研究结果显示,松树盖度、灌木层平均高度和距离浅滩距离是影响海南巢址选择的关键因子;周放等[5]指出,距水边距离、坡度和植被覆盖度是海南巢址选择的重要指标。而本研究结果显示,灌木盖度、巢树高和乔木郁闭度对海南巢址选择影响最大,其次是距林缘距离和距水边距离。尽管各研究结果受研究地和生境因子选择的不同而不完全相同,但均反映出植被盖度、巢树高和距水边距离等几项是最主要的影响因子。植被盖度、巢树高与营巢隐蔽性有关,可归纳为安全因素。茂盛的植被可作为海南良好的遮蔽物,减少被空中和地面天敌发现的风险。高大的巢树,可提供更高的筑巢位置,远离地面各种干扰,保障繁殖巢的安全。距水边距离与觅食有关,可归纳为食物因素。巢距水边越远,对海南觅食产生的直接负面影响越大[17]。海南到觅食场所的路程越近,越能节约时间成本,提高觅食效率。可见,安全因素是海南巢址选择的首要因素,其次是食物因素。
另外,许多鹭科鸟类有利用旧巢址或旧巢树的习惯[26-29]。本研究中,海南在君武森林公园中连续多年在A 区和B 区繁殖,且2013、2014 年连续2 年在同一棵树上营巢。研究团队也监测到柳州市柳江区的一棵马尾松树上连续4 年有海南筑巢。此现象说明旧巢址和旧巢树是海南后续繁殖的重要资源,故对其进行保护尤显重要。
6 存在问题与保护建议
致谢:在君武森林公园海南繁殖监测的过程中,得到刘志元、孙家杰、徐勇及柳州市观鸟协会的大力支持,在此一并表示感谢!
猜你喜欢
故事作文·低年级(2019年8期)2019-08-20
科学24小时(2019年12期)2019-07-24
广东第二课堂·小学(2019年5期)2019-06-14
森林与人类(2018年6期)2018-12-03
乡村地理(2018年1期)2018-07-06
现代园艺(2018年3期)2018-02-10
浙江林业(2016年6期)2016-12-03
浙江林业(2016年5期)2016-06-14
山东畜牧兽医(2011年5期)2011-10-26
山西林业(2010年1期)2010-08-15
