Bacillus stearothermophilus NO2环糊精葡萄糖基转移酶Leu277突变提高α-环糊精产量
2023-08-15 13:46:02孔德民左方圆吴敬王蕾
食品与发酵工业 2023年15期
孔德民,左方圆,吴敬,王蕾*
1(食品科学与技术国家重点实验室(江南大学),江苏 无锡,214122)2(工业生物技术教育部重点实验室(江南大学),江苏 无锡,214122)
环糊精(cyclodextrin, CD)是一种由环糊精葡萄糖基转移酶(cyclodextrin glucosyltransferase, CGTase)以淀粉为底物合成的环状低聚糖[1],一般由6~8个吡喃葡萄糖单元构成,根据糖单元数量可分为α-环糊精(6)、β-环糊精(7)及γ-环糊精(8)。环糊精具有内部疏水及外部亲水的空腔结构,可以和客体分子形成包合物进而增加其溶解性或稳定性[2]。因此环糊精被广泛应用于制药、化妆品、食品、纺织、污水处理等领域[3]。在3种环糊精中,α-环糊精水溶性高且具有抗消化酶水解特性,因此还可以作为可溶性膳食纤维促进肠道蠕动、调节肠道菌群。然而制备α-环糊精时合成和降解同时存在限制了产量的进一步提升,甚至存在明显的产量下降趋势[4],这些问题导致α-环糊精价格昂贵。因此提高α-环糊精产量是一项极有意义的研究。
CGTase的分子改造是一种提高α-环糊精产量的方法[3]。CGTase属于α-淀粉酶家族[5],含有5个结构域[4]。催化中心由TIM(α/β)8结构组成,存在9个糖分子结合位点(-7~+2亚位点)用于结合供体分子及受体分子[4]。根据受体分子的不同,CGTase催化的反应被分为环化反应、歧化反应、偶联反应及水解反应[6]。环化反应是CGTase生产环糊精的关键反应。然而由于存在多个底物结合亚位点,CGTase的环化产物是3种环糊精的混合物[4, 7]。部分来源的CGTase在合成α-环糊精上占据一定的优势,如KlebsiellapneumoniaeM5a1 CGTase的环化产物中α-环糊精质量分数为73%,Thermococcussp.B1001 CGTase的环化产物中α-环糊精质量分数为51%[4],Paenibacillussp.602-1 CGTase的环化产物中α-环糊精质量分数为83%[8]。但是它们对底物的利用率均较低[4]。
此外,有研究报道指出了底物结合亚位点附近的氨基酸残基位点对CGTase催化反应的影响,如-7亚位点主要影响环化反应特异性[4,7,9],-3亚位点影响环化活性[10],+1/+2亚位点主要影响受体分子的结合[6,11]等。近些年,一些研究者展示了定向进化或定点突变改造CGTase,从而提高α-环糊精占比的相关报道[3]。如LI等[12]构建PaenibacillusmaceransJFB05-01 CGTase突变体D372K/Y89R的产物中α-环糊精占比提高23.2%,WANG等[13]构建PaenibacillusmaceransJFB05-01的突变体Y89D的产物中α-环糊精占比提高7.6%,CHAO等[14]构建Bacillussp.602-1 CGTase的突变体Y167H/V536A产物中α-环糊精占比提高5.4%,WANG等[9]构建PaenibacillusmaceransJFB05-01 CGTase突变体R146P/D147A的产物中α-环糊精占比提高12.9%。
本研究选择了表达水平较高[15]、总转化率较高且稳定性较好[4]的B.stearothermophilusNO2 CGTase进行研究。参照之前的研究成果[7,11],本研究在CGTase的-7亚位点构建突变体E142P以提高CGTase的α-环糊精合成特异性,+1/+2亚位点构建突变体L277M、L277F及N353A以提高转糖苷反应特异性。最终,通过检测它们的转化产物,E142P/L277M是最优的生产α-环糊精的突变体,其产量为10.15 g/L,较野生型提高1.7 g/L,并且α-环糊精占比较野生型提高14.3%。本研究为α-环糊精的工业化提供了新的思路,具有重要意义。
1 材料与方法
1.1 试剂、菌株及质粒
酵母粉、蛋白胨、琼脂粉、甲基橙、酚酞、三羟甲基氨基甲烷(Tris)、盐酸、咪唑、3,5-二硝基水杨酸(3,5-dinitrosalicylic acid,DNS)、α-环糊精、β-环糊精等,上海国药生物科技有限公司;2×Phanta Max Master Mix,南京诺唯赞生物科技股份有限公司;DpnI,大连TaKaRa生物科技有限公司。
EscherichiacoliJM109作为重组DNA操作的宿主菌。E.coliBL21作为CGTase表达的宿主菌。pET20b(+)/cgtase质粒参照TAO等[15]构建。
1.2 CGTase突变体质粒构建
使用pET20b(+)/cgtase作为模板质粒,使用表1中的引物1~4构建突变体L277M及L277F的质粒。

表1 定点突变引物
使用pET20b(+)/L277M及pET20b(-)/L277F作为模板质粒,使用表1中的引物5~8构建突变体E142P/L277M、E142P/L277F、L277M/N353A及L277F/N353A的质粒。同时,所有质粒在目的基因的C端均含有His-tag标签,可用于蛋白纯化。在PCR仪中进行扩增后,产物使用DpnI处理。最终转化到E.coliJM109,提取质粒并测序。测序正确的质粒转化到E.coliBL21(DE3)中用于后续目标蛋白的表达。
1.3 CGTase野生型及突变体的表达
将构建好的E.coliBL21(DE3)挑至LB(Luria-Bertani)液体培养基(含100 μg/mL的卡那抗生素)中,37 ℃、200 r/min条件下培养10 h作为种子液。按照体积分数5%的转接量将种子液转接至TB(Terrific-Broth)培养基(含100 μg/mL的卡那抗生素)中,37 ℃、200 r/min条件下培养2 h后调至25 ℃继续培养48 h后终止发酵培养。将获得的发酵液于8 000 r/min离心20 min后取上清液获得粗酶液。
1.4 CGTase的酶活力测定
1.4.1 CGTase环化反应活性检测
1.4.1.1 CGTase生成α-环糊精的酶活力检测条件
反应条件为50 ℃及25 mmol/L、pH 5.5的磷酸盐缓冲液。酶反应体系包括2 mL上述缓冲液配制的1%可溶性淀粉及0.1 mL的适量稀释的酶液。精确反应10 min后加入0.2 mL的3 mol/L HCl溶液终止反应,之后加入0.2 mL的0.44 mmol/L甲基橙溶液进行显色。生成的α-环糊精能够包埋甲基橙,使溶液在505 nm的吸光值降低,依照此制作标准曲线并计算α-环糊精的产量。α-环化反应酶活力定义为在上述反应条件下1 min内催化可溶性淀粉生成1 μmol α-环糊精所需要的酶量为1 U。
1.4.1.2 CGTase生成β-环糊精的酶活力检测条件
反应条件为50 ℃及25 mmol/L、pH 5.5的磷酸盐缓冲液。酶反应体系包括2 mL上述缓冲液配制的1%可溶性淀粉及0.1 mL的适量稀释的酶液。精确反应10 min后加入0.2 mL的0.6 mol/L HCl溶液终止反应,之后加入0.5 mL的0.6 mol/L的Na2CO3溶液并加入0.2 mL的1.2 mmol/L酚酞溶液显色。生成的β-环糊精能够包埋酚酞,使溶液在550 nm的吸光值降低,依照此制作标准曲线并计算β-环糊精的产量。β-环化反应酶活力定义为在上述反应条件下1 min内催化可溶性淀粉生成1 μmol β-环糊精所需的酶量为1 U。
1.4.2 CGTase水解反应活性检测
水解反应酶活力主要指CGTase降解可溶性淀粉生成小分子糖的能力。反应条件为50 ℃及50 mmol/L、pH 5.5的磷酸盐缓冲液。酶反应体系包括1 mL上述缓冲液配制的1%可溶性淀粉、0.9 mL上述缓冲液及0.1 mL适量稀释的酶液。精确反应10 min后加入3 mL DNS溶液终止反应,使用去离子水定容至15 mL。CGTase水解可溶性淀粉生成的小分子糖均为还原糖,能够与DNS反应并显色,依照此制作标准曲线并计算小分子糖的产量,最终检测540 nm的吸光值确定小分子糖的生成量。水解反应酶活力定义:在上述反应条件下1 min内催化可溶性淀粉生成1 μmol麦芽糖当量所需的酶量为1 U。
1.5 α-环糊精的制备
在50 ℃及50 mmol/L、pH 5.5的磷酸盐缓冲液条件下进行CGTase的催化反应制备α-环糊精。使用上述缓冲液配制5%可溶性淀粉作为底物,按照1 g底物5 U环化反应活性的加酶量添加CGTase,反应24 h并固定时间点取样。取样后煮沸灭活用于检测。
1.6 HPLC检测环糊精方法
使用HPLC检测环糊精产量,色谱柱使用Aps-2 Hypersil(4.6 mm×250 mm),柱温40 ℃,流动相为体积分数75%的乙腈,流速0.8 mL/min,检测器为2414型(示差折光检测器)。分别使用2、10、2 g/L的α-环糊精、β-环糊精及γ-环糊精作为标准品。按照1∶1的体积比添加乙腈至样品中沉淀1 h后于12 000 r/min、2 min离心去除大分子底物,用0.22 μm有机相微孔滤膜过滤后即可使用。
1.7 分子动力学(molecular dynamics, MD)结构模拟分析
以野生型B.stearothermophilusNO2 CGTase(PDB ID:1CYG)的三维结构作为模板,利用SWISS-MODEL(https://swissmodel.expasy.org/)[16]模拟突变体E142P/L277M及L277M/N353A的三维结构。利用Amber18进行MD模拟,分别采用ff14SB(蛋白残基)以及TIP3P(水分子)力场,在300 K的温度下模拟10 ns。得到的轨迹利用AMBER18的capptraj分析其均方根偏差(root mean square deviation,RMSD)、均方根波动(root mean square fluctuation,RMSF)、关键氨基酸残基Cα距离、回旋半径(radius of gyration,Rg)及溶剂可及表面积(solvent accessible surface area, SASA)。
2 结果与分析
2.1 突变体质粒的构建及异源表达
参照KONG等[11]对CGTase的Leu277位点转苷水解反应的研究结果,因其具有提高歧化反应并降低水解反应的作用,本文研究L277M及L277F突变体对α-环糊精产量的影响。同时,参照ZUO等[7]对α-环糊精产量的研究,在L277M及L277F突变体的基础上组合E142P及N353A突变体进行研究。
使用E.coliJM109作为宿主,构建突变体L277M、L277F、E142P/L277M、E142P/L277F、L277M/N353A及L277F/N353A的质粒。使用E.coliBL21(DE3)作为表达宿主表达上述突变体。野生型及相关突变体CGTase的十二烷基硫酸钠-聚丙烯酰氨凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)分析如图1所示,均在60 kDa附近有可溶性条带。环化反应活力检测如表2所示,所有突变体的环化反应活力均比野生型低。此外,突变体E142P/L277M及E142P/L277F的α/β值分别提高至野生型的2.52及2.07倍,说明这2个突变体的环化反应特异性向合成α-CD方向偏移。此外,由于Leu277及Gln353位点相关突变体对CGTase水解反应的抑制,所有突变体的水解反应活力均低于野生型,其中L277M/N353A及L277F/N353A组合突变体的水解活力不足野生型的50%。
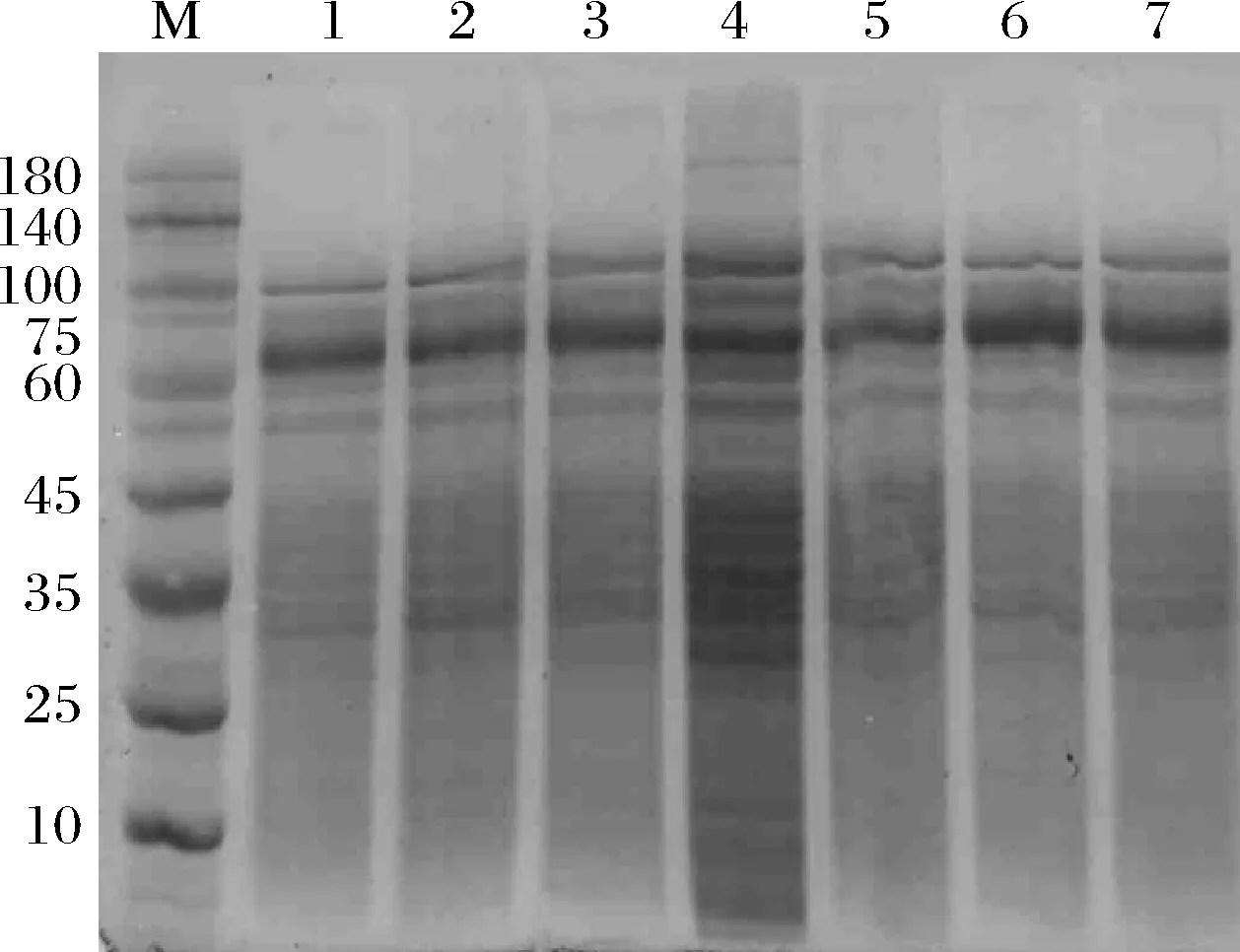
M-markers;1-野生型CGTase;2-L277M突变体;3-L277F突变体;4-E142P/L277M突变体;5-E142P/L277F突变体;6-L277M/N353A突变体;7-L277F/N353A突变体
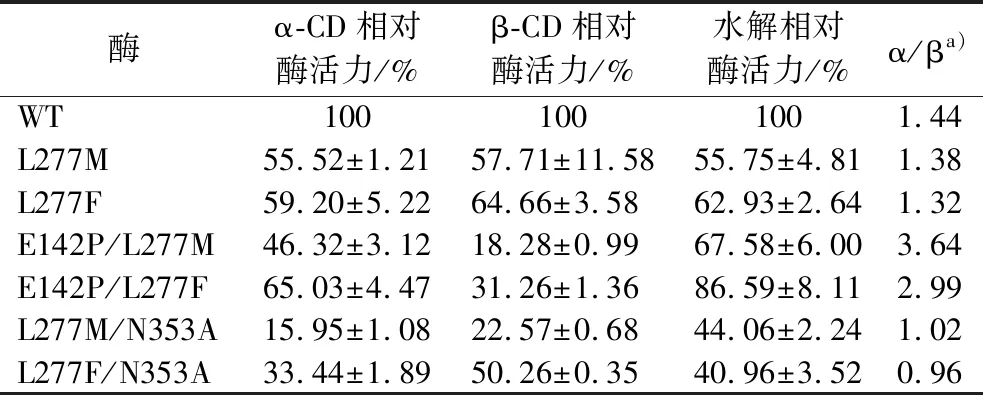
表2 野生型与L277相关突变体CGTase的环化及水解活性
2.2 L277M及L277F突变体制备α-环糊精的应用
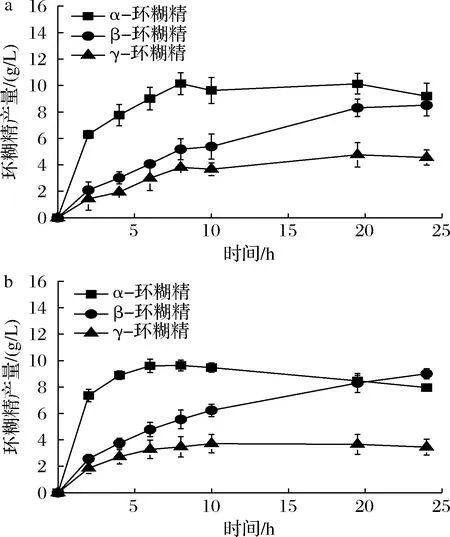
为了确定单突变体对环糊精产量的影响,使用5%可溶性淀粉作为底物进行酶转化反应。L277M、L277F催化生成α-环糊精的最高产量分别为8.97和8.39 g/L(图3),相较于野生型(8.45 g/L,图2)均无明显增加。同时,2种突变体在α-环糊精占比方面也未体现出明显的优势,均为35%左右,较野生型下降3%(表3)。参照KONG等[11]对Leu277相关突变体的分析,该位点影响受体亚位点附近的疏水性以及提高受体底物的亲和力。根据之前的CGTase突变体的相关研究[4]:影响环糊精比例的氨基酸残基位点位于供体位点,特别是—7亚位点。因此,L277M及L277F突变体可能并不能明显提高α-环糊精的产量及占比。但是,由于2个突变体均降低了水解反应(表2),这导致反应体系中小分子糖的减少,从而能够降低α-环糊精的降解。虽然单突变体L277M及L277F能够减弱α-环糊精的降解,但由于其并未从根本改变α-环糊精的产量,因此并不能展现其优势,进而导致突变体的α-环糊精产量及占比与野生型相当。
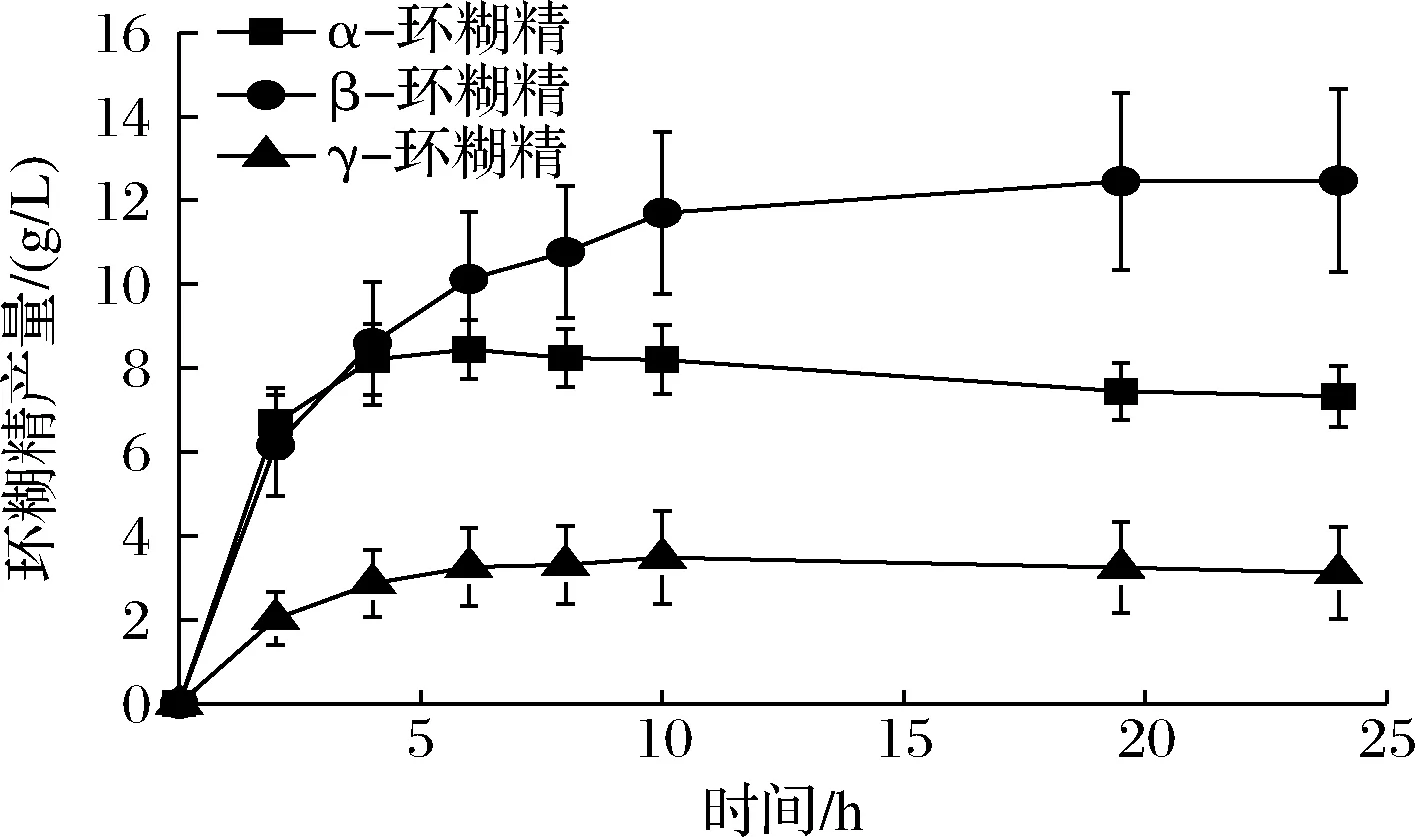
图2 24 h内野生型CGTase合成环糊精的产量变化

a-L277M合成环糊精的产量变化;b-L277F合成环糊精的产量变化
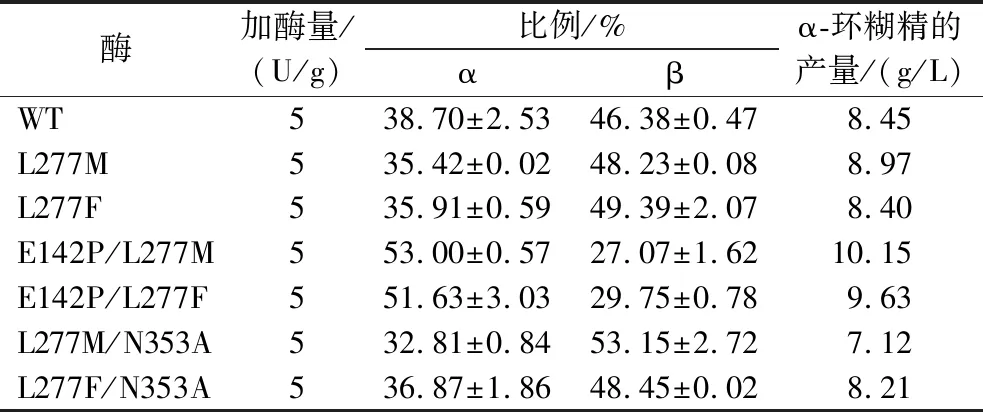
表3 野生型及突变体CGTase的环糊精产物比较
2.3 E142P/L277M及E142P/L277F制备α-环糊精的应用
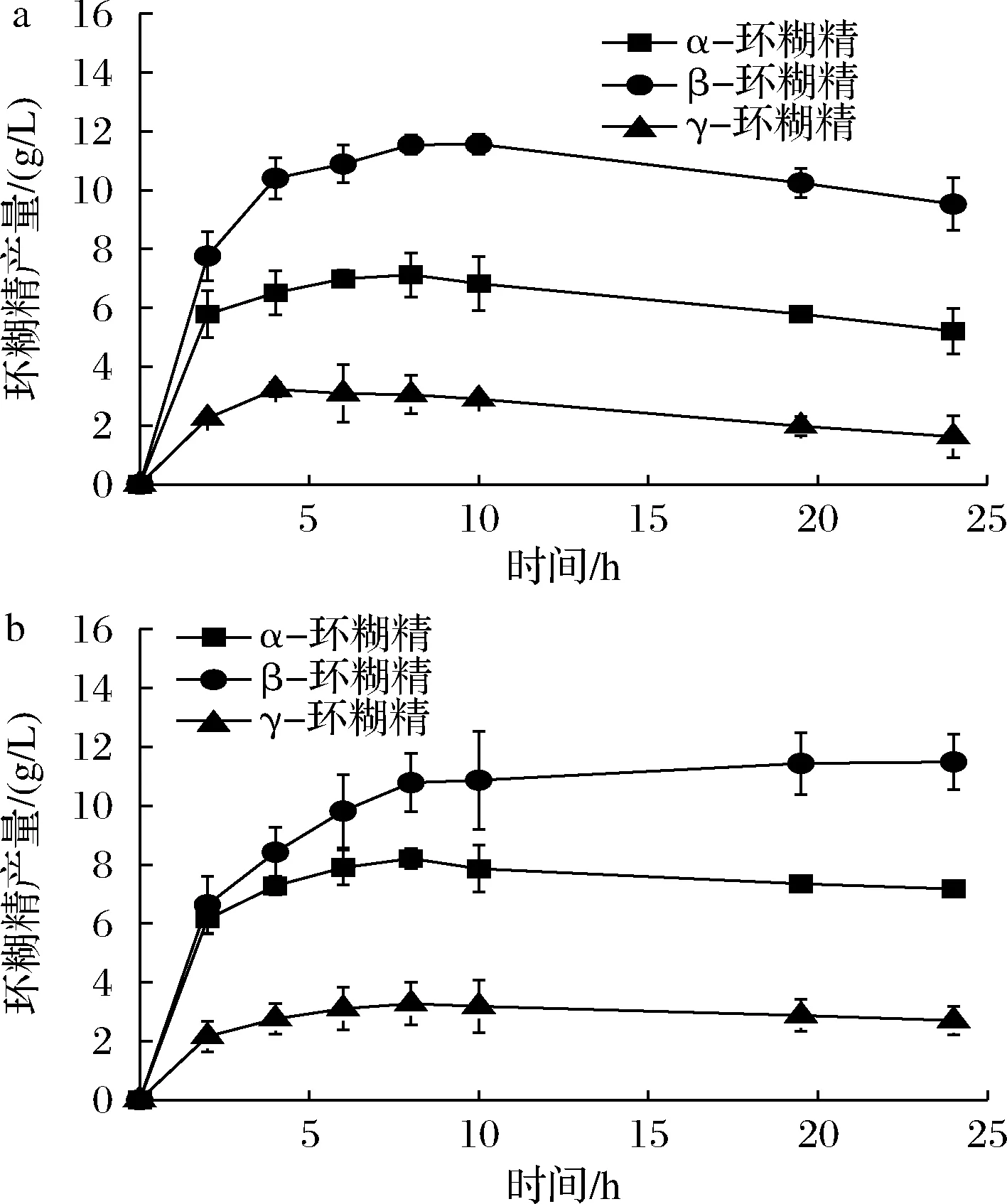
为了更好地利用L277M及L277F降低水解反应的优势,本研究引入了突变体E142P。该位点能够有效地提高产物中α-环糊精的占比(比野生型高9.65%[7]),但在反应后期存在严重的α-环糊精降解现象[4,7,9]。酶转化结果显示(图4),E142P/L277M最高催化产生了10.15 g/L的α-环糊精,而E142P/L277F最高催化产生了9.63 g/L的α-环糊精。相较于野生型,两者的产量均有提高。同时,对比ZUO等[7]报道的E142P最高催化产生约9.4 g/L的α-环糊精,该研究中的双突变体的α-环糊精产量出现了小幅的提升。同时,相较于E142P突变体,E142P/L277M及E142P/L277F在催化反应后期较为稳定,α-环糊精未出现显著的降解,这可能是由于Leu277突变体降低了水解的作用。
a-E142P/L277M合成环糊精的产量变化;b-E142P/L277F合成环糊精的产量变化
2.4 L277M/N353A及L277F/N353A制备α-环糊精的应用
为了进一步降低CGTase的水解反应,本研究引入了突变体N353A。该位点同样能够有效降低CGTase的水解反应,约为野生型的21.3%[7]。将其与L277M或L277F组合,尝试以组合突变体制备α-环糊精。结果如图5所示,L277M/N353A及L277F/N353A最高催化产生α-环糊精分别为7.12及8.21 g/L,较野生型更低。同时α-环糊精占比也比野生型更低(表3)。这可能是由于Asn353和Leu277在空间结构上距离较近,同时突变2个氨基酸残基位点时,该处存在的疏水簇结构[11]或电荷平衡[7]可能遭到破坏,导致CGTase的结构较野生型更不稳定。
a-L277M/N353A;b-L277F/N353A
2.5 MD结构模拟分析
为解析突变体E142P/L277M的优势所在以及L277M/N353A转化率降低的原因,本研究使用MD结构模拟分析了野生型及2个突变体在10 ns内的轨迹变化。RMSD结果如图6-a所示,3个结构在10 ns内均达到平衡状态,其中L277M/N353A较其他2种结构更晚达到平衡。RMSF结果如图6-b所示,L277M/N353A的RMSF在300位氨基酸残基附近波动较大。此外,图6-c显示2个突变体中L277与N353之间的Cα距离高于野生型。上述结果表明L277M/N353A结构波动更大。针对L277及N353所在疏水簇[11]分析SASA,结果如图6-d所示,L277M/N353A的SASA明显增大,这可能是因为双突变体破坏了疏水簇结构所致。此外,L277位点突变后自旋半径增大(图6-e),这与甲硫氨酸(M)侧链长度增大有关;N353位点突变后自旋半径降低,这与丙氨酸(A)侧链长度降低有关。MD结果说明E142P/L277M突变体由于突变位点相互影响并不大,因此对α-环糊精的转化产生了正向效应。与之相反,L277M/N353A突变体由于突变位点均位于疏水簇,对该区域影响较大,导致酶催化活力降低,因此对α-环糊精的转化产生了负向效应。
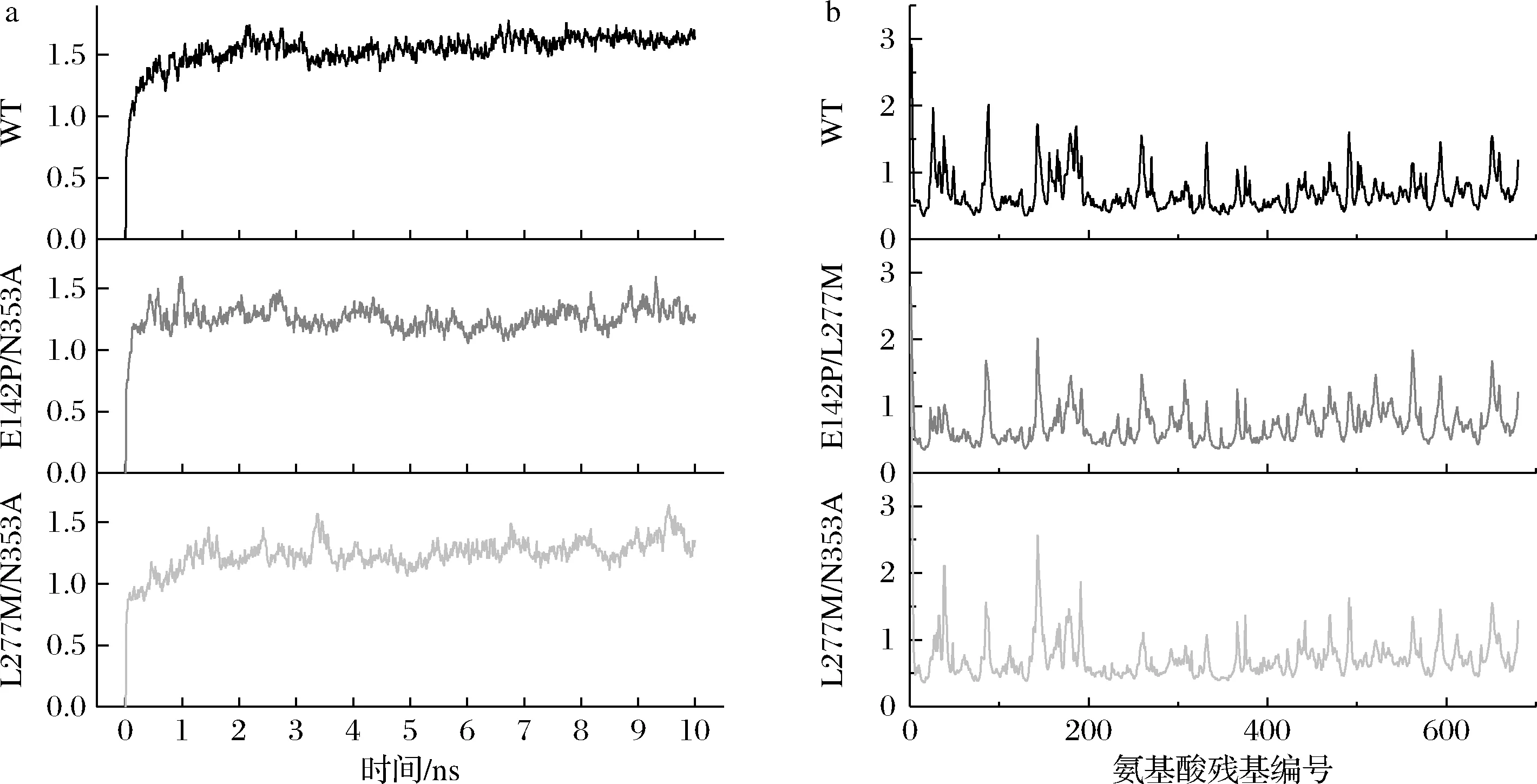
a-野生型及突变体RMSD的结果;b-野生型及突变体RMSF的结果;c-L277与N353A的Cα距离;d-突变体及野生型的SASA变化;e-L277位点的自旋半径;f-N353位点的自旋半径
3 结论
本研究从降低CGTase水解反应及提高环化反应活力对其进行分子改造。L277M、L277F及N353A突变体虽然降低了CGTase的水解反应活力,但其本身对α-环糊精的特异性生产无明显影响。L277M/N353A及L277F/N353A则导致CGTase的α/β值明显降低,因此α-环糊精转化率明显降低。最终的优势突变体为E142P/L277M,在24 h内最高催化生成了10.15 g/L的α-环糊精,较野生型α-环糊精产量提高1.7 g/L。此外突变体E142P/L277M生成的α-环糊精占比较野生型提高14.3%。整体而言,该研究为α-环糊精的生产应用提供了新的思路。
猜你喜欢
云南化工(2021年6期)2021-12-21 07:30:56
科学(2020年2期)2020-08-24 07:57:00
中成药(2018年8期)2018-08-29 01:28:08
中成药(2018年4期)2018-04-26 07:12:43
安徽医科大学学报(2016年12期)2017-01-15 14:21:44
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
生物技术通报(2015年1期)2015-04-10 16:15:19
山东医药(2015年40期)2015-02-28 14:28:45
食品科学(2013年23期)2013-03-11 18:30:11
食品科学(2013年13期)2013-03-11 18:24:19