
构树叶物候持续时间的纬度格局
2023-08-07 03:42王鑫洋杜彦君
热带生物学报 2023年4期
王鑫洋,王 媛,杨 华,张 泽,杜彦君
(海南大学 林学院,海口 570228)
国际生物计划(International Biological Programme,IBP)将物候学定义为研究周期性生物事件的时间、这一时间变化的生物和非生物驱动因素及相同或不同物种阶段之间相互关系的科学[1],如植物开花和展叶的开始时间、结束时间等,对生态系统功能的各个方面都有重要影响[2-3]。叶物候持续时间对生态系统功能和树木生产力也有很大影响[4]。对于落叶树种,春季展叶和秋季叶衰老的时间决定了树冠的持续时间,从而决定了其年际营养生长速率和繁殖成功率[5]。植物碳汇潜力和植被对气候系统的反馈也依赖于叶物候的持续时间,而叶物候持续时间的变化是森林气候变化的一项指标[6-7]。在过去的40 a里,树木的生长期平均每10 a延长了2~3 d[8-9]。由于森林在陆地碳储存方面发挥着重要作用,其叶物候持续时间的变化不但将对森林产品和生物多样性,而且对全球气候本身都有重要影响[10],因此,研究植物叶物候持续时间及其驱动机制对于了解气候变化对森林生态系统的影响具有重要意义[7],但以往的叶物候研究对叶物候持续时间关注较少[11]。植物叶物候在种间和种内存在显著差异,且沿着环境梯度呈现明显的变化。研究大多使用平均性状值来研究物种尺度的功能性状,隐藏了许多内部差异,包括种内不同种群间的差异[12],特别是广布种[13]。因此,为了预测广布树种的命运,往往需要评估物种对当地条件的适应机制及其应对环境变化的适应潜力。气候变化对植物的叶物候有着显著的影响,其中温度是控制植物叶物候最主要的因素[14]。纬度代表的是一个更加复杂的环境条件综合体,大量非生物环境因子存在明显的交互作用,形成了强烈的自然选择压力,驱动广布种的快速适应进化过程[15]。但是目前尚缺对广布种沿纬度梯度的叶物候变化的研究,阻碍了对其未来可能受气候变化影响的评估与分析。
构树(Broussonetiapapyrifera)是一种属于桑科、构属的乔木树种,分布于热带、亚热带地区以及我国绝大部分地区[16]。构树是一种先锋植物,分布广、适应性强、抗性强,是适合我国三北地区种植的防护林和山区大力推广种植的树种[17]。由于叶片本质上是获取能量的器官,因此季节性和非季节性环境中叶子在时间(叶物候)和空间(冠层结构)上的排列可被视为植物碳获取策略的核心要素[18]。构树不同种源的叶表现出了明显的地理变异趋势,其叶形态变异的主要环境因素可能是与采种点的经纬度相关的气候差异[19],如构树各种源叶片的叶面积、叶长、叶宽和绒毛密度与采种点纬度呈极显著的负相关,这表明低纬度种源比高纬度种源叶片更大[19]。目前关于几个地理上分布较远的种群的展叶物候、特定物种种群之间的物候差异及其驱动机制方面的研究较少[20]。这些问题不仅限制了评估气候变化对构树生产力的影响,也将影响预测构树植物的入侵风险和碳汇能力变化。本研究基于中国物候监测网多个城市站点记载的1963—2008年的物候数据拟研究以下3个问题:(1)构树叶物候持续时间是否有明显的纬度格局?(2)构树叶物候持续时间的延长是否是由展叶和落叶期共同引起的?(3)驱动构树叶物候性状纬度差异的关键环境因子是什么?以期从纬度地理差异角度研究广布种不同种群的物候分化,为评估物种未来的分布范围变化及灭绝风险提供参考。
1 材料与方法
1.1 研究区域与物候数据选取中国暖温带和亚热带地区,该区域包含了半湿润和半干旱2个气候区。多为大陆性季风气候,该区域四季分明,降雨集中在夏季,冬季干燥,水热条件在季节与空间上存在差异,植物物候也表现出显著的时空变化特征,对气候变化响应较为敏感[21],因此,适合作为研究植物物候变化的区域。
选用的物候观测资料来自中国物候观测网(Chinese Phenology Observation Network,CPON),由于人为观测和虫害等因素导致物候数据部分缺失[22],依据观测数据较丰富和连续性较好的原则,选择含观测期内(1963—1968年,1973—1978年,1983—1991年,2003—2007年)的展叶期、叶变色期和落叶期的叶物候观测记录[23],为了满足统计分析的最小样本量,排除了数据少于10 a的站点记录[23],共有九个物候观测站点有充足的构树叶物候数据符合筛选要求,分别是:北京、西安、洛阳、芜湖、镇江、长沙、南昌、贵阳和桂林,纬度范围25°14′—39°9′N,每个站点的具体纬度见表1。采用儒略日期(Julian day)换算法(1月1日为第1天,2月1日为第32天,依此类推),将展叶期、落叶期转换为距离当年1月1日的天数[24]。然后计算每一个站点内每一叶物候期累年平均时间,作为该站点叶物候数据。
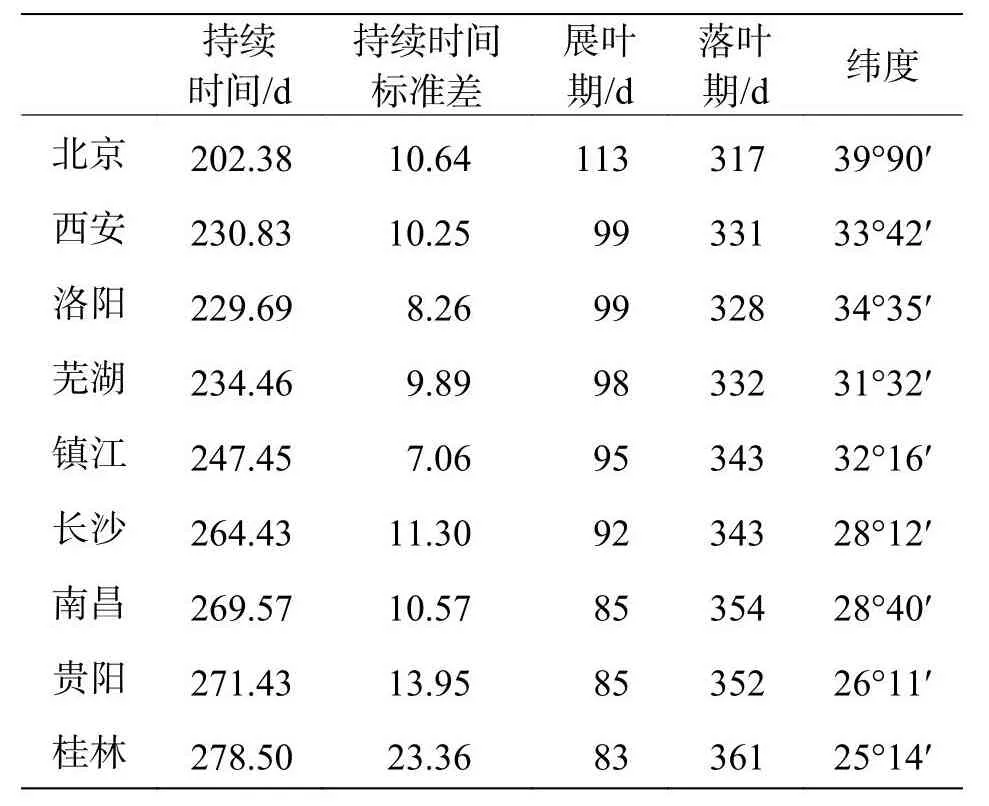
表1 9个站点构树叶物候数据和纬度坐标数据
1.2 气候数据气候数据来源于中国气象数据网(http://data.cma.cn/)[25],包括9个物候站点对应的气象站点的年平均气温、累年月平均气温、累年月平均降水量(1981—2010年)[25]。之前的研究结果揭示展叶季前1~2个月的平均气温影响展叶期的年度变化,2~3个月的平均气温影响落叶期的年度变化[21]。在此基础上,笔者根据9个研究站点构树平均物候数据(平均展叶期为4月5日),将2—3月平均气温定义为展叶季前温度,10—11月平均气温定义为落叶季前温度(平均落叶期为12月7日),并用12月至翌年2月的平均气温作为冬季平均气温;降水量选取时间段与此相同。在选取气象站数据点时依据物候观测站点选取同一位置的气象站点,其中除镇江站外,其余物候观测站点均有与之地理位置相同的气象站点,则直接使用这些站点的气象数据[26]。镇江无与之地理位置相同的气象站点,以地理位置最近的南京站替代[26]。
1.3 研究方法所有统计分析用R语言4.2.2版(R Core Team,2022)。分别对3个构树叶物候进行统计分析,分别为:展叶期、落叶期和持续时间,计算每个站点内构树叶物候儒略日的平均值。叶物候持续时间计算为落叶期与展叶期的差值。对于秋季物候,主要关注的是叶片脱落,而不是叶片颜色的变化,因为任何剩余的绿叶都能够进行大量碳同化,支持生物量增长[27]。为了评价纬度对构树叶物候的影响,采用线性模型(LM)。笔者将3个叶物候性状数据与以往研究认为与展叶落叶物候相关的6个环境因素进行随机森林回归(RF)来检测与叶持续时间最相关的变量。6个环境因子分别是:展叶季前温度、冬季温度、落叶季前温度、展叶季前降水量、冬季降水量和落叶季前降水量[28-29];选择此方法的原因是它不需要严格的数据假设,可以更好地处理多重共线性和非线性关系[30-31]。生成变量指数MSE(均方误差)以证明解释变量对响应变量的重要性[32];根据MSE指数的大小选择最关键的因素,MSE越大变量越重要。
2 结果与分析
2.1 物候格局1963—2007年各站点构树种群平均展叶期分布在3月25日(桂林)至4月24日(北京)之间,平均日期为4月5日。落叶期分布在11月14日(北京)至12月28日(桂林)之间,平均日期为12月7日。叶物候持续时间分布在202.38 d(北京)至278.5 d(桂林)之间,平均为247.63 d(表1)。
回归结果显示,构树的叶物候特征,无论是展叶期、落叶期和持续时间,都有明显的纬度梯度格局(展叶期:R2=0.93,P<0.001;落叶期:R2=0.86,P<0.001;叶持续时间:R2=0.94,P<0.001)(表2)。在种群水平,种群分布每降低1个纬度,构树展叶期平均提前1.97 d,落叶平均延迟2.88 d,持续时间增加5.32 d。展叶期有较落叶期更显著的纬度梯度格局,但是落叶期的斜率绝对值大于展叶(表2,图1)。

图1 构树叶物候纬度梯度格局
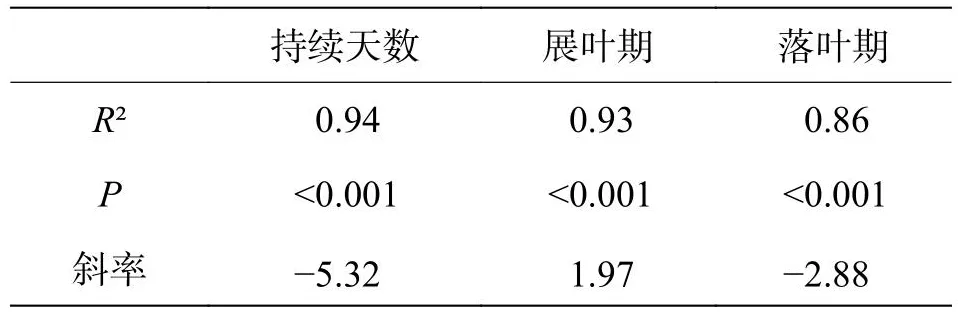
表2 构树展叶期、落叶期和叶物候持续时间与纬度回归结果
2.2 气候因子与叶物候持续时间的相关程度
如表3所示,对于纬度梯度上的构树种群叶物候,所选取的6个环境因子都有一定的相关性。造成构树3个叶物候性状纬度差异的环境因子较为一致,其最重要的前3个环境因子均是:展叶季前温度、落叶季前降水量和冬季温度。其余3个环境变量也对构树叶物候性状有影响,但相对不重要。同时,同一环境因子与不同叶物候特征的重要程度大小有一定差异,展叶季前温度是持续时间和展叶期最重要的环境因子(MSE值分别为:11.01、10.30),而落叶季前降水是落叶期最重要的环境因子(MSE值:9.51)。
3 讨 论
3.1 构树叶物候性状的纬度分布差异本研究结果表明,构树叶物候性状沿纬度梯度有显著的变化规律;展叶期与纬度呈正相关,高纬度地区种群明显有更晚展叶的趋势;落叶期与纬度呈负相关,高纬度地区种群有明显更早落叶的趋势;叶物候的持续时间与纬度呈负相关关系,高纬度地区种群有着更短的叶持续时间。因此来自单个位置种群的叶物候数据不能用于预测其他地方同一物种的叶物候变化[33]。木本植物的物候通常被认为是适应当地气候的,许多使用起源测试的研究强调了种群之间的物候差异[34]。对于同一树种不同纬度的种群来说,预计植物物候的可塑性将有利于种群的持续存在,因为物候事件的时间决定了竞争(由较长的生长季节促进)和避免霜冻损害(由生长期较短)[35]。但是,以前的相关纬度格局研究大多单独关注于展叶期或叶变色期,对叶物候持续时间的空间变异研究尚属首次[27,36]。
树木物候的种内变异性可能会严重限制研究人员预测物候对全球气候变化的一般反应的能力[36]。本研究结果表明了秋季落叶相比春季展叶对低纬度叶物候持续时间延长有更大的贡献,从而使低纬度地区种群获得更长的年际生长时间和更多的生物量增长。之前的研究也发现了展叶与落叶期对变暖响应的不一致,并认为落叶期主导植物的叶物候持续时间:对于平均叶持续时间延长的物种,落叶期延迟,而那些植被周期平均缩短的物种的落叶日期提前[37]。但是,并没有发现不同地理位置同一种群的秋季物候及叶持续时间差异。研究结果表明,同一物种的高纬度地区种群相较于低纬度地区种群具有较高的光饱和CO2同化率和较低的比叶面积[38-39],推测这是对较短生长季[40]的适应。此种情况下,尽管两者相关,但叶物候持续时间的差异所导致潜在生产力和碳固存能力的差异小于预期差异[41-42]。
3.2 构树叶物候的关键环境因子在对环境因子的分析中,展叶季前的温度是展叶期和叶物候持续时间最关键的环境因子。尽管大多数植物控制长叶的确切生理机制还不清楚,但可以明确的是,大多数物种的叶子发育对温度极其敏感。不同纬度种群长叶时间的转变是由于温度在植物生长发育中所起的主导作用,此结果验证了植物物候对气候变化高度敏感这一猜测,并成为气候变化的绝佳指标[43]。构树叶物候纬度差异可以用气候因子的变化及其与叶物候的相关性来解释,但不同叶物候性状的主要气候驱动因素有所差异。本研究结果中,落叶物候最相关的环境因子是落叶季前的降水量。这种正相关可能与水分胁迫对秋季植物生长的影响有关。有限的水势会抑制植物生长和光合作用活动[44],增加叶绿素退化和植物死亡的风险,并提前叶片衰老的时间[45]。因此,季前期间降水增加可以缓解水分胁迫并延长秋季物候。目前对于植物秋季物候调控机制的研究尚存许多争议,对于给定的纬度,落叶树种的抗寒性发生在生长停止之后和叶片衰老之前,并且生长停止、叶片衰老和年内及年际之间的抗寒性之间也存在很强的相关性[46]。这解释了秋季物候对冬季温度和展叶季前温度敏感的可能原因,也强调了更深入地研究秋叶衰老及其潜在过程作为叶持续时间和全球变化驱动因素的重要性。
不同纬度站点相同气候因子的变化导致当地树木的展叶和落叶物候都发生了变化,而秋季叶物候与主要环境因子的相关程度较小。这与之前的研究结果相一致,即与春季物候相比,秋季物候更多地受到遗传变异的影响[47]。全球温度升高可能会促进春季叶片提前展叶,并在秋季促进生长停止和叶片衰老[48-49]。已有研究结果表明,过去几十年的气温升高导致树木的春季物候发生了变化,而秋季叶物候的变化较小且不太一致[50-51]。因此笔者推断,构树可能通过适应性实现更早的叶片展叶和更长的叶物候持续时间。然而,后期生长停止和落叶期的发展可能将涉及种群差异和自然选择。未来的几十年里,世界上大部分温带地区的植物预计将继续提前落叶[52-53]。在较长一段时间内,一些树种可能会在局部范围内被消灭,并根据气候变化改变它们的活动范围[53]。因此对构树不同种群叶物候持续时间的环境调控机制和空间分布变化分析将有助于评估植物物种栖息地的变化[54],可能对生态系统动态产生重大影响,如碳和水的循环及植物-动物相互作用[55-56],以及对依赖树种的行业,如苹果、桃子和构树生产可能的经济影响。
4 结 论
本研究发现了构树种群叶物候持续时间沿纬度增加逐渐缩短的地理格局,并且发现这一格局是由低纬度种群展叶期提前和落叶期延迟共同导致。此外,本研究还发现构树展叶期和叶物候持续时间的主要调控因子是展叶季前温度。相比展叶期,构树的落叶期受季前降水的影响程度大于季前温度,这为进一步理解构树秋季物候的调控机制提供了参考。但是要清楚理解这一机制及不同纬度种群物候差异原因还需要其他相关工作,如系统发育差异的研究以及基因调控机制的研究等。
猜你喜欢
热带作物学报(2022年6期)2022-07-08
绿色中国(2019年16期)2019-11-26
中国林业产业(2019年3期)2019-03-25
中国扶贫(2017年16期)2017-08-16
齐鲁周刊(2017年29期)2017-08-08
西南农业学报(2017年5期)2017-06-23
西南学林(2016年2期)2016-11-08
河北林业科技(2016年5期)2016-11-08
西南农业学报(2016年5期)2016-05-17
技术经济与管理研究(2014年11期)2014-03-11
