
糜子矮秆突变体海5农艺性状及对GA3的敏感性鉴定
2023-08-07 01:04:16郭英杰刘洋刘晓婕魏玮王瑶张帅王振山闫留延朱学海贾小平
作物杂志 2023年3期
郭英杰 刘洋 刘晓婕 魏玮 王瑶 张帅 王振山 闫留延 朱学海 贾小平
(1 张家口市农业科学院,075000,河北张家口;2 河南科技大学农学院,471000,河南洛阳;3 张家口市杂交黍子技术创新中心,075000,河北张家口)
糜子(Panicum miliaceumL.)起源于我国,是一年生禾本科植物,有近万年的栽培历史[1]。其营养丰富,富含蛋白质、膳食纤维和维生素等,兼具食用和药用价值[2],同时可作为饲料[3]和着色剂[4]等,具有较高的经济价值。糜子抗旱性强,耐贫瘠,生育周期短,在干旱半干旱地区有独特的优势,是该区域理想的主粮替代物。近年来,随着环境恶化、水资源缺失和粮食问题突显,糜子越来越受到人们的关注。但是由于糜子存在倒伏现象普遍和产量不高等问题[5],导致其优势未能完全发挥出来。
在临床扫描诊断中,一共出现13例良性包块和9例恶性包块、8例转移淋巴结(其中有6例为CT扫描诊断、2例为MRI扫描诊断)。关于良性包块、恶性包块以及转移淋巴结的诊断来源和影像学特征等均需要综合临床资料进行统计和分析。
株高与抗倒伏、收获指数、光合作用效率密切相关,控制株高能够有效增强植株的抗倒伏性,提高产量[6]。目前,对植株矮化的研究主要集中在小麦、水稻和玉米等作物上。小麦目前正式命名的矮秆基因有25 个(RHT 1-25),其中有9 个通过EMS 诱变得到,赤霉素(GA3)不敏感矮秆基因有5 个,分别为RHT1、RHT2、RHT3、RHT10和RHT21,GA3敏感矮秆基因有20 个[7],实际生产中利用较多的有5 个,分别为RHT1、RHT2、RHT8、RHT9和RHT10。研究[8]发现,RHT1能够使小麦株高降低24%,促进小穗发育,使分蘖数和穗粒数增加。水稻株高矮化主要与一些激素的合成及信号转导途径有关,如目前发现的与GA3合成或转导相关的矮秆突变体sd1、d18、d35、slr1、gid1和gid2等[9],与油菜素内酯合成或转导相关的矮秆基因D2、D11、BRD1、OSDWARF4等[10]。矮化玉米有抗倒伏和产量高的优势,且一般株型紧凑,适合密植,综合性状良好[11]。目前,已发现200 多个控制玉米株高的QTL[12]。控制玉米株高的基因中隐性基因占比较多,如BR1、BR2、D1、D2、D3和D5等[13-15],其中D1、D2、D3和D5为GA3敏感型,显性基因占比较少,如D8[16]等。研究[17-18]发现,BR2基因可使玉米茎节间距变短,茎粗增大,抗倒伏性增强;突变体与矮化植株配合的F1代植株株高可进一步降低。
但是,目前糜子的研究基本聚焦在栽培[19]、抗逆[20]、资源收集及多样性分析[21]和品质分析[22]等方面,关于矮化研究很少,导致糜子育种理论基础相对薄弱。本研究通过EMS 诱变野生型糜子得到性状稳定遗传的糜子矮秆突变体海5(dm5),对其农艺性状及喷施GA3后表型变化进行研究,探索其矮化的可能生理机制,为糜子矮秆育种及其产能优势发挥提供一定参考。
1 材料与方法
1.1 试验地概况
试验于河北省张家口市农业科学院沙岭子基地进行。沙岭子位于华北平原北,海拔643m,属温带大陆性季风气候,年气温在-10.0℃~23.4℃,年均日照时数2800h,年均降水400mm,昼夜温差大,光资源丰富,无霜期长。试验地为水地,土壤为黏土,地势平坦,肥力良好。
1.2 试验设计
以野生型糜子wt260 及糜子矮秆dm5为研究材料。wt260 为石湖千斤糜,由中国农业科学院陆平研究员提供。dm5为由wt260 经甲基磺酸乙酯(EMS)诱导(诱变浓度为0.4%,时间为20h)后在突变体库中获得的能够稳定遗传的突变体材料。
2.1.2 农艺性状变化 由表2 可得,dm5的株高为60.72cm,相比wt260,降低了44.00cm;分蘖数为6.40,增加了4.90;穗柄长为6.42cm,减小了3.69cm;穗长为13.52cm,减小了2.60cm。这些性状差异均达到极显著水平(P<0.01)。茎粗、茎节数、倒3 叶长度、倒3 叶宽度、倒3 叶面积与wt260 相比均无显著差异(P>0.05)。
鉴于此,以京津冀城市群为载体,以土地综合承载力为基础,扩大生态环境容量,不断调整优化区域经济发展结构和空间结构,确保区域经济健康增长,推进京津冀城市群土地综合承载力约束下区域经济发展的持续增效策略,促进京津冀城市群土地综合承载力与区域经济发展由良好协调向优质协调发展转变。
1.3 测定项目与方法
2.1.1 叶片颜色变化 由图1 和表1 可得,野生型wt260 植株苗期及抽穗期叶片呈绿色,成熟期逐渐变黄。dm5苗期叶片呈黄色,其SPAD 值为19.96,与wt260 相比差异达到极显著水平(P<0.01),抽穗期植株叶片变绿,其SPAD 值为31.16,与wt260 叶片相比无明显差异;成熟期叶片呈绿色,其SPAD 值与wt260 相比有极显著差异(P<0.01)。说明糜子矮秆dm5幼苗为黄苗,随着植株生长,叶色变绿,且能够维持至植株成熟,推测其衰老期较wt260 晚。
1.3.2 茎节细胞 在糜子灌浆期,选取喷施及未喷施GA3的wt260 和dm5主茎茎节进行切片。观察茎秆纵切面同一视野薄壁细胞的大小及数量。
1.4 数据处理
利用Excel软件和SPSS 21进行数据统计分析。
2 结果与分析
2.1 dm5 主要农艺性状
1.3.1 表型数据 在试验田随机选取长势均一的糜子dm5及其对照材料wt260 各10 株,在糜子苗期、抽穗期和成熟期检测剑叶的相对叶绿素含量(SPAD 值);在糜子成熟期,测量株高、茎粗、茎节数、分蘖数、节间长、穗柄长、穗长、倒3叶长和倒3 叶宽,计算倒3 叶面积。在糜子拔节期分别用水和50mg/L GA3溶液喷施wt260 及dm5各5 株,共喷3 次,每次间隔7d,喷施处理液需均匀分布在植株表面。在糜子成熟期,分别记录wt260 及dm5试验组和对照组的株高、茎节数、穗柄长、穗长和节间长度。

表1 不同生长时期wt260 和dm5 的SPAD 对比Table 1 Comparison of chlorophyll contents(SPAD value)of wt260 and dm5 at different stages

图1 wt260、dm5 苗期及成熟期植株Fig.1 Plants of wt260 and dm5 at seedling and mature stages
试验于2017 年6 月进行,每个小区4 行,行长3m,行距33cm,株距10cm,5 次重复,田间管理与当地大田生产管理方式相同。
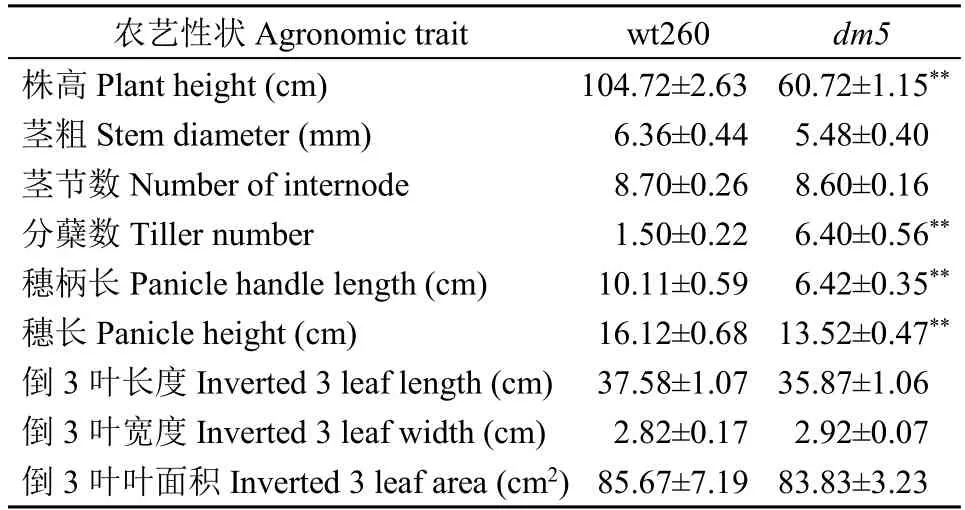
表2 wt260 及dm5 农艺性状Table 2 Agronomic traits of wt260 and dm5
2.2.2 各节间长度变化 由图3 可知,喷施GA3后,相比对照组,wt260 第5~8 节间长度均极显著增大(P<0.01),第7 节间长度增大最多,为58.71%,第5、6、8 节间长度分别增大32.45%、47.57%、40.78%,第1~4 节间长度无明显变化。dm5第1~4 节间长度均极显著增大(P<0.01),第1 节间长度增大最多,达到171.54%,第2、3、4 节间长度分别增大97.74%、78.95%、49.33%,第5~8 节间长度无明显变化。对于wt260 而言,植株的节间序数越大,喷施GA3后的节间长度变化率越大,推测wt260 植株顶部茎节对GA3更敏感;对于dm5而言,植株的节间序数越小,喷施GA3后的节间长度变化率越大,推测dm5植株基部茎节对GA3更敏感;整体而言,dm5节间长度变化率大部分在70%以上,wt260 节间长度变化率均在60%以下,说明dm5对GA3更敏感。

图2 wt260 及dm5 节间长度变化Fig.2 Internode length changes of wt260 and dm5
2.2 喷施GA3 后dm5 表型变化
由图4 可得,植株wt260 及dm5茎秆纵切细胞均排列规则,整齐。通过观察发现喷施GA3后,wt260 及dm5的茎秆纵切细胞长度均增大。前述研究发现wt260 及dm5喷施GA3后株高均显著增大,茎节数均没有明显差异,说明喷施GA3后,茎秆纵向细胞伸长,使wt260 及dm5植株增高。
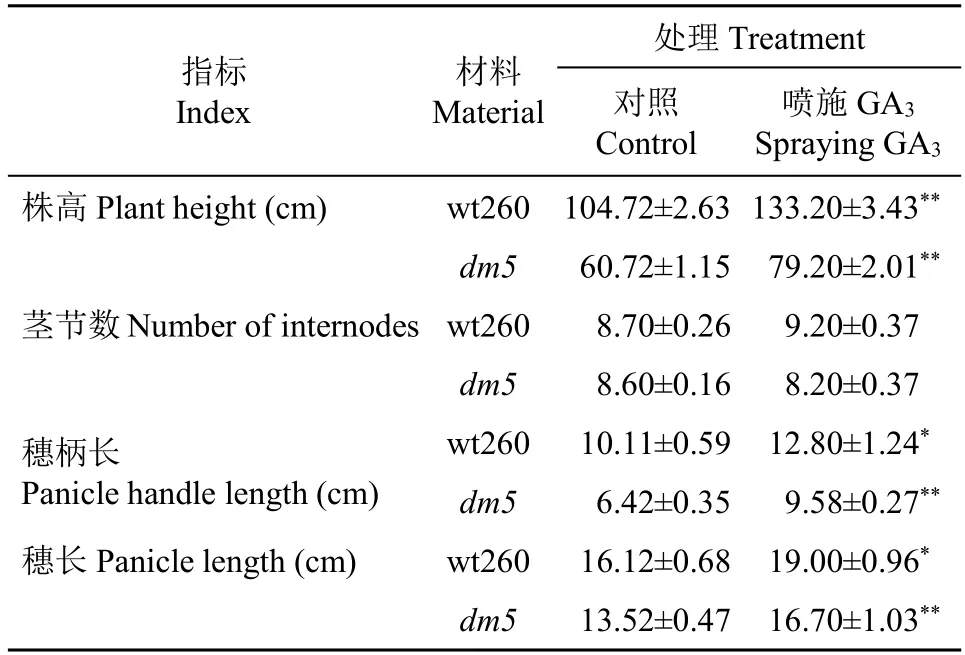
表3 喷施GA3 前后wt260 及dm5 主要表型变化Table 3 Main phenotypic changes of wt260 and dm5 before and after spraying GA3
2.1.3 各节间长度变化 由图2 可得,对wt260及dm5节间长度进行分析发现其趋势一致,均随着节间序数的增加先增大后减小。wt260 的第4 节间长度最大,第1 节间长度最小,dm5的第5 节间长度最大,第1 节间长度最小。与wt260 相比,dm5的第1~8 个节间长度均明显减小,差异均极显著(P<0.01)。dm5各节间长度的变化率为-65.25%~-45.31%,其中第1 节间的减小幅度最大,其余减小幅度则在50%左右。整体对比wt260及dm5发现,节间序数越小,节间长度减小率越大,节间序数越大,节间长度减小率越小。

图3 喷施GA3 前后wt260(a)及dm5(b)节间长度变化Fig.3 Internode length change of wt260(a)and dm5(b)before and after spraying GA3
2.3 dm5 主茎剖面细胞形态学变化
2.2.1 主要表型变化 由表3 可得,喷施GA3后,野生型wt260 的株高为133.20cm,增加27.20%,穗柄长为12.80cm,增加26.61%,穗长为19.00cm,增加17.87%,以上性状喷施和不喷施差异显著或极显著。dm5的株高为79.20cm,喷施GA3后增加30.43%;穗柄长为9.58cm,喷施GA 3 后增加49.22%;穗长为16.70cm,喷施GA 3 后增加23.52%,且喷施和不喷施差异均极显著(P<0.01),说明GA3的喷施能够使糜子野生型及其突变体的株高、穗柄长和穗长明显增大,但对糜子的茎节数没有显著影响(P>0.05)。

图4 喷施GA3 前后wt260 及dm5 主茎纵切细胞Fig.4 Main stem longitudinal observation of wt260 and dm5 before and after spraying GA3
3 讨论
另外由于生鲜产品在质量方面存在很大的不确定性,商品容易出现问题,售后纠纷很多屡见不鲜。根据调查,有消费者反应称收到每日优鲜水果腐败变质甚至生虫,也有罐头生锈等情况,但在跟售后反应此情况以后,每日优鲜客服却表示不予退还,只能退货款的30%,而且还存在客服不及时处理的情况。
本研究通过EMS 诱变得到dm5,其株高及各茎节长度均显著缩短,茎节数无明显变化。其茎秆纵切细胞排列规则,相比于野生型wt260,dm5细胞长度减小,推测造成糜子突变体dm5矮化的原因可能是茎秆细胞长度的减小。郝小花等[23]对EMS 诱变得到的超矮秆突变株水稻ag1进行研究,发现倒2 节间的细胞长度显著变短。马宁[24]对不同株高甘蓝型油菜进行研究发现,矮秆主要与其茎秆纵切细胞较小有关,这些结果均与本研究结果一致。此外造,成植株矮化的原因还有茎节间数目减少[18]、茎节细胞数目减少[23]或茎节细胞数目减少与细胞长度缩短共同作用[25]等。
植株高度的降低能够显著提高抗倒伏能力,直接影响作物的高产和稳产[26-27]。对比dm5及wt260,dm5第1~4 茎节长度变化显著,喷施GA3后,矮秆突变体dm5第1~4 茎节增长最多,这一结果从正反两面说明了糜子矮秆突变体dm5基部茎节长度对株高的影响更大,这与前人研究结果一致。崔淑佳等[28]对矮秆、中秆、高秆共20 个小麦品种(系)的株高及各节间长进行研究,发现矮秆小麦主要是通过缩短不同节间长度来降低株高,特别是基部节间。魏国才[29]发现,造成矮秆玉米植株较矮的主要原因是节间长度缩短,特别是基部节间缩短最多,而节间数无变化。此外,基部茎节也与植株的抗倒性密切相关。苏亚蕊等[30]对不同小麦材料不同生育时期抗倒性的研究发现,较长的基部茎节长度对植物抗倒伏能力有消极作用。在玉米上,基部茎节的表现也决定了其抗倒伏能力。在华北平原,30%~60%的玉米倒伏发生在茎秆,且茎折均发生在基部第3~5 茎节[31-32]。这就直接或间接地说明基部茎节在植株矮化及抗倒伏中起到重要作用。
1.1.1 CKD的诊断标准[7](1)肾损害(肾脏结构或功能异常)≥3月,伴或不伴肾小球滤过率(GFR)下降。可出现以下表现之一:病理学检查出现异常;或肾损害指标显示阳性:如血、尿成分异常或影像学检查异常;(2)GFR<60 mL/(min·1.73 m2)≥3月,有或无肾脏损伤的依据。
喷施GA3后,dm5的株高显著增大,推测dm5可能为GA3敏感型矮秆突变体,由于内源GA3合成途径受阻,使植株体内缺乏内源GA3,导致矮秆。其株高及茎节增长率均高于野生型,说明相比于wt260,dm5对GA3更敏感,这可能与二者含有不同内源生物活性的GA 水平有关[6]。但dm5的株高并未恢复至wt260 株高,且二者差异显著(P<0.01),这可能与GA3喷施的时间、浓度和次数等有关。
(4)通过实行支农惠农政策减缓贫困。从2002年开始,政府陆续出台了一系列增加农民收入、减少农民支出的支农惠农强农政策,直接增加农民的收入,对减少农村贫困产生了重要的作用。2002—2014年“四项补贴”(良种补贴、种粮补贴、农机购置补贴和农资综合补贴),从146亿元增加到1683亿元[注]2014年数据来自财政部:http://yss.mof.gov.cn/2014czys/2015071t201507091629855.html;2002年数据来自《中国财政年鉴2003》。。研究[1]表明这些以“多予少取”为取向的支农惠农强农政策,具有较好的益贫效果,对农村减贫发挥了积极作用。
作为一种新的糜子矮秆资源,dm5矮化的分子机制还有待进一步研究,挖掘其可能存在的矮秆突变位点,筛选糜子矮秆基因分子标记,可为糜子育种的发展提供新思路。
通过网络教学平台来组织教学是一种新的教学模式,为职业学校数学教学提供了丰富的教学手段。它通过网络技术将数学教学内容以文字、图形、图像、动画等多种形式表现出来,使得数学内容在展示上更加灵活、生动、形象。
4 结论
经EMS 诱变得到的糜子矮秆突变体dm5株高明显降低,造成这一现象的原因可能为茎节纵向细胞长度减小、基部茎节长度显著下降。通过喷施GA3初步推测突变体dm5为GA3合成缺陷型矮秆突变体。因此,突变体dm5可为糜子矮化育种提供研究材料。
猜你喜欢
中国粮油学报(2022年6期)2022-07-23 10:16:08
今日农业(2022年8期)2022-07-05 00:05:26
作物学报(2022年3期)2022-01-22 07:11:18
河北果树(2021年4期)2021-12-02 01:14:52
麦类作物学报(2021年5期)2021-09-23 09:19:00
河北果树(2020年1期)2020-02-09 12:31:18
现代农村科技(2019年12期)2019-01-06 04:54:21
种子(2018年10期)2018-11-14 08:54:00
麦类作物学报(2018年9期)2018-10-09 02:33:56
老友(2017年7期)2017-08-22 02:36:36
