
不同草本植物对铜镍复合胁迫的生理耐受性
2023-08-04 13:01李景峰郭全恩梁鹏飞何海鹏南丽丽
干旱地区农业研究 2023年4期
李景峰,郭全恩,梁鹏飞,何海鹏,南丽丽
(1.甘肃农业大学草业学院/草业生态系统教育部重点实验室 甘肃 兰州 730070;2.甘肃省农业科学院土壤肥料与节水农业研究所,甘肃 兰州 730070)
随着人类经济社会的迅速发展,矿山开采、化肥农药施用、生活垃圾和交通运输等过程中产生的大量重金属元素被排放到土壤、水体中,导致土壤、生物和水资源受到不同程度的重金属污染,造成土壤地力下降、农作物被污染,生态环境遭到破坏[1-2]。铜(Cu)和镍(Ni)是工农业生产中较为常见的2种重金属元素,可被植物吸收积累在可食用部位,并通过食物链的传递进入人及牲畜体内,危害人类健康[3]。因此,探索出科学、高效、合理的重金属修复措施有利于人类健康和环境的可持续发展,而植物修复以其环保、高效、成本低等优势,被广泛用于重金属污染土壤的修复[4]。近年来,我国在Cu、Ni耐性及超累积植物研究方面取得了一定的进展[5-6],但实际生产中需要修复的土壤多是2种或2种以上重金属复合污染,复合污染的毒害作用比单一元素的污染复杂很多[7],大大增加了重金属污染土壤的植物修复难度。因此,筛选耐复合铜镍胁迫植物及与耐复合铜镍性能相关的指标已迫在眉睫。
Cu、Ni对植物产生的毒害首先会引起植物生理指标变化,其可能与蛋白质结合,抑制酶活性,也可能通过破坏质膜的通透性阻碍植物的正常代谢,引起植物萎黄、根畸形和生长迟缓[6,8-9]。大量研究发现Cu、Ni胁迫对植物光合作用、渗透调节物质、酶活性以及根系生长会产生影响,李芹梅等[10]针对Cu2+对滇水金凤(Impatiensuliginosa)花色变化生理研究结果显示,随着Cu2+含量增加,植物花瓣可溶性糖和可溶性蛋白含量下降,而类胡萝卜素和脯氨酸含量变化不明显;刘丽杰等[11]研究表明,铜镍复合胁迫造成车前草(Plantagoasiatica)株高和生物量显著降低,叶绿素含量先升后降,MDA含量、POD、SOD活性和CAT活性显著升高;朱成豪等[12]通过研究铜、锌、镉复合胁迫对麻疯树(Jatrophacurcas)幼苗生长的影响发现,麻风树幼苗可溶性糖含量和根系活力随复合胁迫浓度增加呈先升后降趋势。由此可见,重金属胁迫对植物生理指标影响较为明显,筛选与耐铜镍复合胁迫密切相关的生理指标较为重要,可为后期筛选更多的耐铜镍复合胁迫植物提供捷径。前人已研究发现了一些耐铜或耐镍植物,但这些植物大多生物量小,修复土壤所需时间较长,修复效率较低[13]。因此本研究选用生物量大(清水苜蓿和鹰嘴紫云英)和可以多次刈割(黑麦草和早熟禾)的草本植物进行铜镍复合污染修复试验,以筛选出对铜镍复合胁迫耐受性较好的植物及其耐铜镍指标,为土壤重金属污染修复及寻找耐重金属污染植物提供理论基础。
1 材料和方法
1.1 试验材料
供试材料为苗期筛选的耐铜镍植物[14]‘卡特’多年生黑麦草(Loliumperenne‘Cutter’),‘午夜草地’早熟禾(Poapratensis‘Midnight’),‘清水’紫花苜蓿(Medicagosativa‘Qingshui’),‘卢塔纳’鹰嘴紫云英(Astragalussinicus‘Lutana’),种子由甘肃农业大学草业学院提供;供试土壤取自甘肃省金昌市金川矿区农田土壤(Ni、Cu背景值含量分别为258.2 mg·kg-1和261.1 mg·kg-1),采集土样经自然风干后去除杂物、研磨、过100目筛,待用。
1.2 试验设计
试验在甘肃省农业科学院温室进行,将22.5 kg过筛土壤装入口径、底径、高为34.5、23.0、44.5 cm的花盆中,于2021年4月23日播种经10% H2O2溶液消毒后的供试植物种子,其中早熟禾和黑麦草播量为40 kg·hm-2,播深2~3 cm;鹰嘴紫云英和紫花苜蓿播量为20 kg·hm-2,播深1~2 cm。出苗后间苗,每盆保留生长一致、分布均匀的幼苗30株,待生长至开花初期(10%开花),设置6个铜镍复合胁迫浓度梯度,各复合胁迫中铜和镍浓度均为0(CK)、100、200、400、800、1 600 mg·kg-1,用CuCl2·2H2O和NiCl2·6H2O转化成质量,配成1 L溶液,每盆每5 d浇铜镍复合液500 mL,待生长20 d后采样测定其相关生理指标,每个处理重复4次。
1.3 测定指标及方法
株高差:处理前后定株,用卷尺测10个植株株高取平均值,株高差=处理后株高-处理前株高。
地上、地下生物量及根冠比:每盆取10个植株,地上、地下部位分离后烘干,分别称重,即为地上和地下生物量;根冠比=地下生物量/地上生物量。
生理指标参考邹琦[15]的方法进行,其中可溶性糖(Soluble sugar)含量采取蒽酮比色法测定,可溶性蛋白(Soluble protein)含量采用考马斯亮蓝G-250染色法测定,丙二醛(Malondialdehyde,MDA)含量采用硫代巴比妥酸法测定,脯氨酸(Proline)含量采用酸性茚三酮比色法测定,过氧化物酶(Peroxidase,POD)活性采用愈创木酚法测定,超氧化物歧化酶(Superoxide dismutase,SOD)活性采用氮蓝四唑(NBT)光化还原法测定,过氧化氢酶(Catalase,CAT)活性采用高锰酸钾滴定法测定,根系活力(Root activity)采用氯化三苯基四氮唑(TTC)法测定,叶片相对含水量(Relative water content)采用饱和称重法测定,叶绿素a(Chlorophyll a,Chl a)、叶绿素b(Chlorophyll b,Chl b)、类胡萝卜素(Carotenoid)、叶绿体色素(Chloroplast pigment, Chl)含量采用分光光度法测定;植物叶、茎、根中的铜(Cu2+)、镍(Ni2+)含量采用灰化消解处理及原子吸收分光光度计法[16]测定。
1.4 数据统计分析
采用Excel 2010进行图表制作,利用SPSS 20.0进行方差、回归分析。各指标耐铜镍系数=铜镍胁迫下的测定值/CK测定值[17]。采用隶属函数对供试植物进行耐铜镍综合评价,其中与铜镍胁迫正相关的指标采用公式Fij=(Xij-Xjmin)/(Xjmax-Xjmin)计算,与铜镍胁迫负相关的指标采用公式Fij=1-(Xij-Xjmin)/(Xjmax-Xjmin)计算,式中,Fij为i品种的j性状测定的具体隶属值;Xij为i品种j性状测定值;Xjmin为j性状中测定的最小值;Xjmax为j性状中测定的最大值。
权重采用客观赋权法进行计算:正相关指标计算公式为Ij=Cj/Sj,负相关指标计算公式为Ij=Sj/Cj,式中,Ij是一个无量纲数,反映该评价指标CK与铜镍处理下测定值之比;Cj为第j个指标的CK测定值;Sj为第j个指标在胁迫处理下的测定值。最后通过归一化,计算出每个评价指标的权重Wj=Ij/∑Ij,综合评价值D=∑(Fij×Wj)[18]。
2 结果与分析
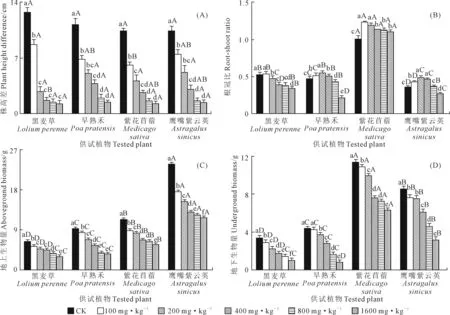
2.1 铜镍胁迫对供试植物生长的影响
由图1A可知,供试植物株高差均为正值,表明株高在铜镍胁迫后较胁迫前均有所增加,但株高差随铜镍胁迫程度加剧呈降低趋势,表明铜镍胁迫浓度越大,株高受抑制作用越明显。当铜镍浓度为1 600 mg·kg-1时,相比CK,株高降幅以黑麦草最大(90.54%),早熟禾最小(86.83%)。
铜镍胁迫下,供试植物地上和地下生物量均随铜镍浓度增加呈减小趋势,在浓度为1 600 mg·kg-1时,相比CK,黑麦草、早熟禾、紫花苜蓿和鹰嘴紫云英的地上和地下生物量分别降低53.60%和69.90%、61.50%和82.25、49.40%和44.44%、50.67%和62.94%,所有胁迫浓度下,鹰嘴紫云英的地上生物量和紫花苜蓿的地下生物量显著(P<0.05)高于同一胁迫浓度下的其他植物(图1C、1D)。
由图1B可知,铜镍胁迫下,除紫花苜蓿根冠比高于CK外,其余植物的根冠比呈先增后降趋势,其中黑麦草和紫花苜蓿的根冠比在铜镍浓度为100 mg·kg-1时达到最大,早熟禾和鹰嘴紫云英的根冠比在铜镍浓度为200 mg·kg-1时达到峰值,且紫花苜蓿的根冠比在铜镍胁迫下均大于1,其余植物的根冠比则均小于1。
2.2 铜镍胁迫对供试植物叶绿素含量的影响
图2显示,随着铜镍复合胁迫浓度增加,4种植物的叶绿素a(Chl a)、叶绿素b(Chl b)和叶绿体色素(Chl)含量均降低,类胡萝卜素含量均逐渐增加,其中早熟禾的Chl含量呈波动降低趋势。与相应CK相比,黑麦草、早熟禾、紫花苜蓿的Chl a含量,黑麦草、早熟禾、鹰嘴紫云英的Chl b含量以及4种植物的Chl含量均在胁迫浓度为100 mg·kg-1时开始显著降低(P<0.05),鹰嘴紫云英的Chl a含量及紫花苜蓿的Chl b含量均在胁迫浓度为200 mg·kg-1时开始显著降低(P<0.05);黑麦草、早熟禾、鹰嘴紫云英的类胡萝卜素含量在胁迫浓度为100 mg·kg-1时开始显著增加(P<0.05),紫花苜蓿在胁迫浓度为200 mg·kg-1时开始显著增加(P<0.05)。当胁迫浓度≤200 mg·kg-1时,鹰嘴紫云英的Chl a含量显著高于其他植物(P<0.05);所有胁迫浓度下,紫花苜蓿的Chl b含量显著高于其他植物(P<0.05);当胁迫浓度≤800 mg·kg-1时,紫花苜蓿的Chl含量显著高于其他植物(P<0.05);当胁迫浓度≥400 mg·kg-1时,早熟禾的类胡萝卜素含量显著高于其他植物(P<0.05)。当胁迫浓度为1 600 mg·kg-1时,与CK相比,Chl a、Chl b和Chl含量分别以紫花苜蓿(44.26%)、鹰嘴紫云英(70.22%和36.59%)降幅最大,以黑麦草(24.47%)、紫花苜蓿(45.91%)和黑麦草(25.04%)降幅最小;类胡萝卜素含量以紫花苜蓿增加最多,是CK的13.83倍,黑麦草增加最少,是CK的6.76倍。
注:不同小写字母表示同一植物不同胁迫浓度之间差异显著(P<0.05);不同大写字母表示同一浓度下不同植物间差异显著(P<0.05)。下同。
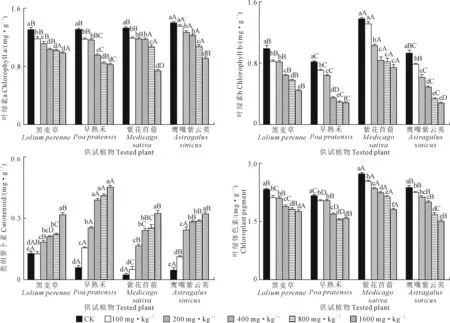
图2 铜镍胁迫对供试植物叶绿素含量的影响
2.3 铜镍胁迫对供试植物叶片相对含水量和根系活力的影响
由图3可知,4种植物叶片相对含水量和根系活力均随铜镍复合胁迫浓度的升高而降低。与相应CK相比,黑麦草的叶片相对含水量在所有胁迫浓度下均显著降低(P<0.05),而其余植物在胁迫浓度为400 mg·kg-1时才开始显著下降(P<0.05)。不同植物根系活力均在胁迫浓度为100 mg·kg-1时开始较CK显著降低(P<0.05);且在不同胁迫浓度下,早熟禾的根系活力均高于其他植物。当胁迫浓度为1 600 mg·kg-1时,与CK相比,叶片含水量降幅为紫花苜蓿最大(20.98%),黑麦草最小(17.59%);根系活力降幅则为鹰嘴紫云英最大(69.32%),黑麦草最小(14.71%)。
2.4 铜镍胁迫对供试植物渗透调节物质及MDA含量的影响
图4显示,随铜镍胁迫浓度的升高,供试植物的可溶性糖和可溶性蛋白含量呈先逐渐增大后减小的趋势。与相应CK相比,黑麦草和早熟禾的可溶性糖含量在胁迫浓度为100 mg·kg-1时开始显著增加(P<0.05),紫花苜蓿和鹰嘴紫云英的可溶性糖含量在胁迫浓度为400 mg·kg-1时显著升高(P<0.05);4种植物在胁迫浓度为800 mg·kg-1时可溶性糖含量均显著升高至最大值(P<0.05),黑麦草较CK增加最多(8.44倍),紫花苜蓿增加最少(3.65倍)(图4A)。与相应CK相比,黑麦草和鹰嘴紫云英的可溶性蛋白含量均在胁迫浓度为200 mg·kg-1时开始显著增加(P<0.05),早熟禾和紫花苜蓿则在胁迫浓度为100 mg·kg-1时开始显著升高(P<0.05),4种植物均在胁迫浓度为800 mg·kg-1时达到峰值,与CK相比,早熟禾上升最多(327.81%),黑麦草上升最少(65.93%)(图4B)。
供试植物的脯氨酸和MDA含量均随铜镍胁迫浓度的升高持续上升,且均在浓度为1 600 mg·kg-1时升至最高,此浓度下与CK相比,脯氨酸和MDA含量分别以早熟禾(11.60倍)和黑麦草(62.90%)增加最多,以紫花苜蓿(3.35倍和40.62%)增加最少。与相应CK相比,黑麦草和鹰嘴紫云英的脯氨酸含量在胁迫浓度为200 mg·kg-1时开始显著增加(P<0.05),早熟禾和紫花苜蓿则在胁迫浓度为100 mg·kg-1时即开始显著上升(P<0.05);相同胁迫浓度下,紫花苜蓿的脯氨酸含量均显著高于其他植物(P<0.05)(图4C)。与CK相比,4种植物的MDA含量均在胁迫浓度为100 mg·kg-1时开始显著升高(P<0.05);所有胁迫浓度下,紫花苜蓿的MDA含量均高于相同浓度处理的其他植物(图4D)。
2.5 铜镍胁迫对供试植物抗氧化酶活性的影响
由图5可知,供试植物的SOD、POD和CAT活性均随铜镍胁迫浓度的增加呈先升后降趋势,且均在胁迫浓度为100 mg·kg-1时开始显著上升(P<0.05)。黑麦草的SOD活性在胁迫浓度为800 mg·kg-1时最大,早熟禾、紫花苜蓿、鹰嘴紫云英的SOD活性在胁迫浓度为400 mg·kg-1时升至最高;与CK相比,各植物SOD活性最大值增幅为黑麦草(107.55%)最大,早熟禾(30.20%)最小(图5A)。4种植物的POD和CAT活性均在胁迫浓度为800 mg·kg-1时达到峰值,之后随胁迫浓度增加显著下降(P<0.05),其峰值与CK相比,POD和CAT活性增幅分别以鹰嘴紫云英(176.93%)和紫花苜蓿(85.87%)最大,以黑麦草(131.65%)和鹰嘴紫云英(72.42%)最小(图5B、5C)。
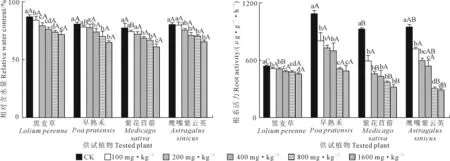
图3 铜镍胁迫对供试植物叶片相对含水量及根系活力的影响
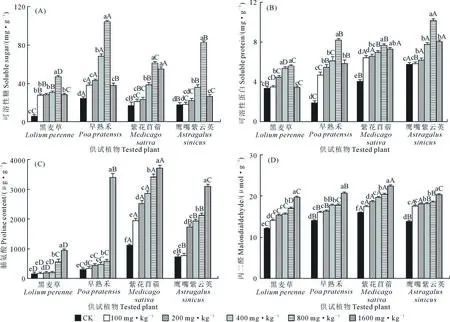
图4 铜镍胁迫对供试植物渗透调节物质及MDA含量的影响

图5 铜镍胁迫对供试植物抗氧化酶活性的影响
2.6 各供试植物根、茎、叶中Cu2+和Ni2+分布规律
由图6、图7可知,供试植物根、茎、叶中Cu2+和Ni2+含量均随着铜镍胁迫浓度的升高而升高。黑麦草和早熟禾植株体内的Cu2+含量表现为根>茎>叶,紫花苜蓿体内的Cu2+含量表现为叶>茎>根;当铜镍胁迫浓度≤100 mg·kg-1时,鹰嘴紫云英体内的Cu2+含量根略大于茎,其他胁迫浓度下其体内的Cu2+含量表现为叶>茎>根(图6)。各供试植物根、茎、叶中Ni2+含量分布不同,其中黑麦草在<800 mg·kg-1胁迫下各器官Ni2+含量表现为茎>叶>根,而在高浓度(≥800 mg·kg-1)胁迫下,各器官Ni2+含量表现为茎>根>叶;早熟禾的各器官Ni2+分布表现为根>茎>叶,紫花苜蓿和鹰嘴紫云英的各器官Ni2+分布表现为叶>茎>根(图7)。

图6 铜镍胁迫下各供试植物体内Cu2+含量分布规律
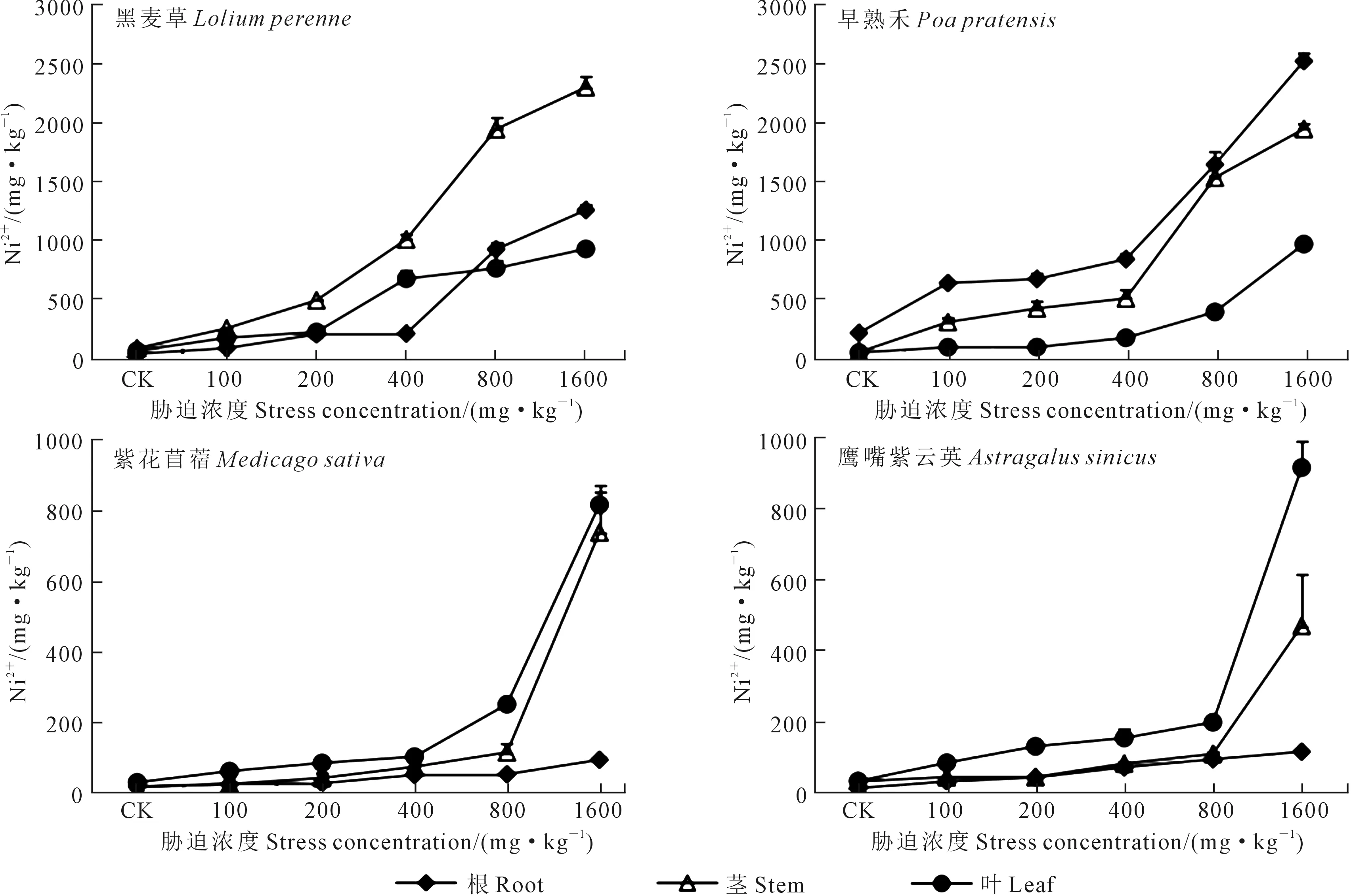
图7 铜镍胁迫下各供试植物体内Ni2+含量分布规律
2.7 供试植物耐铜镍性能评价及指标筛选
利用19项生理指标各胁迫浓度下的值进行隶属函数加权平均,对4种供试植物进行耐铜镍胁迫能力综合评价(表1),其强弱表现为:紫花苜蓿(0.565)>鹰嘴紫云英(0.538)>早熟禾(0.449)>黑麦草(0.448)。
为分析各生理指标与植物耐铜镍性之间的关系,探讨植物耐铜镍鉴定指标,需建立可靠的耐铜镍性评价模型。因此,对19个生理指标进行逐步线性回归分析,以各胁迫浓度下耐铜镍性评价值为因变量,各单项指标耐铜镍系数为自变量,进行耐铜镍指标预测,并建立最优回归方程[19]:Y=0.225X1-0.015X2-0.015X3-0.019X4+0.078X5-0.011X6+0.355(R2=0.989),其中,X1、X2、X3、X4、X5和X6分别代表地下生物量、可溶性糖、可溶性蛋白、脯氨酸、SOD和Cu2+含量。可见,以上6个指标对4种供试植物耐铜镍性筛选具有指导作用,可作为其他耐铜镍植物筛选的关键指标。
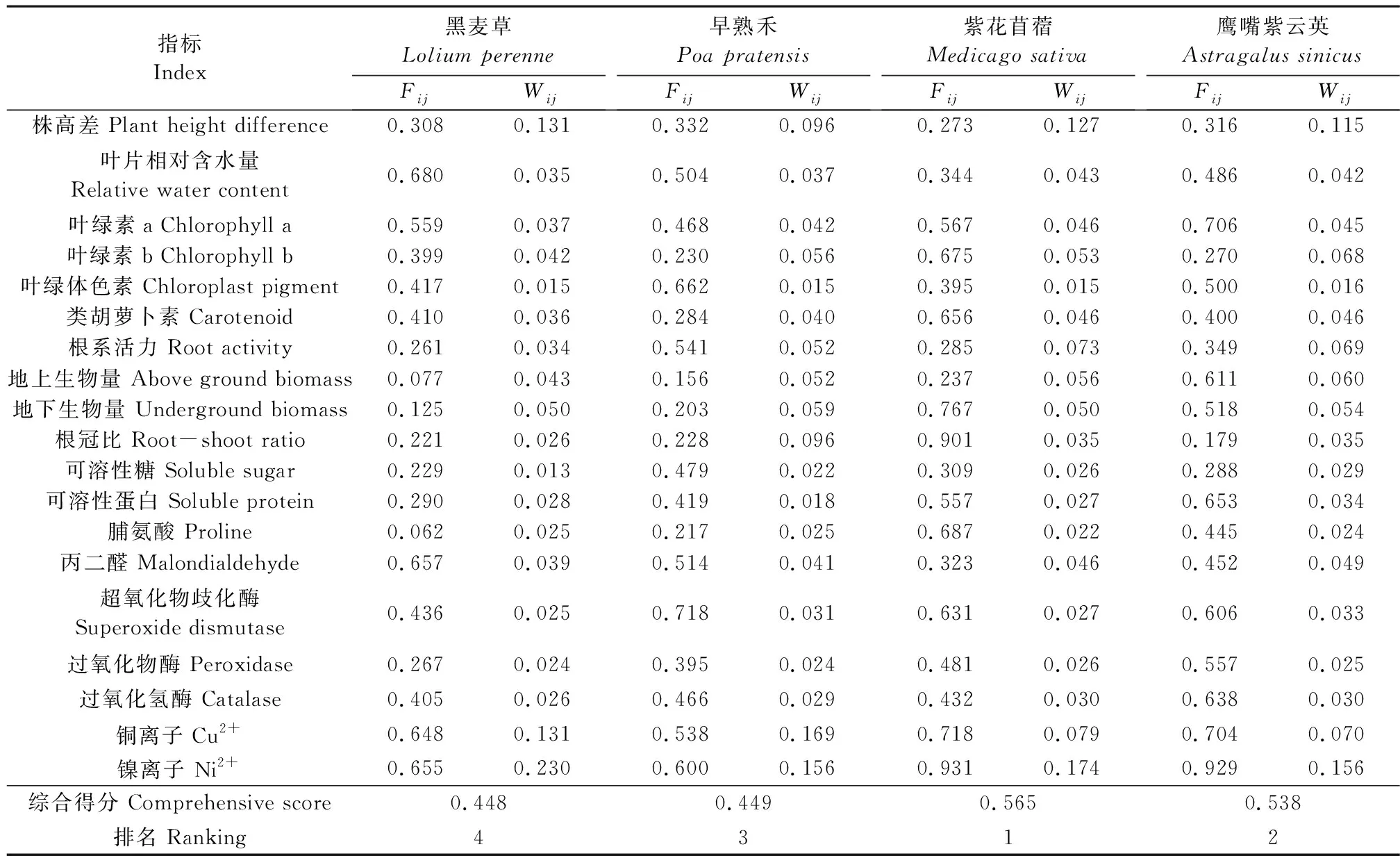
表1 铜镍胁迫下各指标隶属值及综合评价
3 讨 论
重金属胁迫会阻碍植物的正常生长和生理生化反应,引起植株根系活力下降、株高和生物量降低、组织脱水、叶绿素减少、呼吸光合作用减弱、代谢紊乱等变化,这些变化均可导致植物体蛋白质性能和抗氧化系统酶功能的改变[13]。
根系作为直接接触土壤的植物器官,会最先受到重金属胁迫毒害,造成根系主根长度变短、一级侧根和根毛数量减少、根系活力下降等,最终表现为地下生物量降低[20],影响根系对土壤养分和水分的吸收,进而导致植物株高、叶片相对含水量、地上生物量等降低[13]。因此根系形态可作为植物受重金属胁迫的主要特征指标。赵娜等[21]研究显示,玉米根系受镍胁迫后生长变缓、根须变短、根毛变少、根系发达程度降低,最终表现为地下鲜质量下降。本研究表明,铜镍胁迫下4种供试植物均表现为根系活力下降,地下生物量、株高、地上生物量、叶片相对含水量降低,根冠比也发生相应变化;此外,回归分析得出地下生物量对植物耐铜镍性能贡献较大。因此,地下生物量可作为评价植物耐铜镍性能强弱指标之一。
Cu是植物叶绿体色素的重要组成成分,可参与植物光合电子的传递,同时也是叶绿素合成过程中部分酶的催化剂,过量的Cu会抑制叶绿素合成和破坏光合器官,从而影响光合速率;而Ni可以延缓植物叶片衰老,使叶片保持较高的叶绿素水平[22]。因此,Cu2+含量可表征植物受重金属污染时的光合特性[13]。本研究显示,随着铜镍胁迫浓度的增加,4种植物叶片的叶绿素a和叶绿素b含量逐渐下降,类胡萝卜素含量逐渐增加,叶绿体色素含量均降低。究其原因可能是植物体内高浓度的Cu破坏了叶片组织中的叶绿体结构,使叶绿素合成受阻,从而导致叶绿素a和叶绿素b含量降低;叶黄素是一种类胡萝卜素,也是优异的抗氧化剂[23],其在铜镍胁迫下会为阻止细胞氧化而不断增加,使类胡萝卜素含量上升,但这不足以抵抗细胞衰老和氧化,叶绿素的降解大于抗氧化色素的合成,进而导致叶绿体色素含量下降。因此,植物体内Cu2+含量也可作为植物耐铜镍性能的评价指标。
重金属胁迫时,植物会通过积累渗透调节物质(可溶性糖、可溶性蛋白、脯氨酸)维持细胞渗透压以减少伤害,其中可溶性糖可提高细胞保水和吸水能力,增加植物原生质的弹性;可溶性蛋白具有维持细胞渗透压、调节代谢和提高抗性的作用[24]。张诗雯等[25]研究显示,千屈菜(LythrumsalicariaLinnaeus)在重金属铜和铬胁迫下,可溶性糖和可溶性蛋白表现出一定的渗透调节抗性。本试验中,4种供试植物可溶性糖和可溶性蛋白含量均随铜镍浓度增加呈先升后降趋势,且均高于CK,说明植物为保护自身不受伤害而积累了可溶性糖和可溶性蛋白;此外,紫花苜蓿和鹰嘴紫云英表现出更好的耐铜镍特性,其体内可溶性糖含量显著增加时的胁迫浓度高于早熟禾和黑麦草,其可溶性蛋白总含量高于早熟禾和黑麦草。脯氨酸含量增加可增强植物对逆境的抵御作用,4种植物的脯氨酸含量随铜镍胁迫浓度增加而增加,且在高浓度时显著增加,可能是可溶性糖和可溶性蛋白合成量减小所致,通过三者含量平衡消除重金属对细胞的氧化损伤或维持胞内外渗透势,表明脯氨酸有效抵御了铜镍胁迫对植物的伤害[26]。结合线性回归分析结果可知,可溶性糖、可溶性蛋白和脯氨酸含量可作为植物耐铜镍性能强弱的评价指标。
SOD是维持植物细胞氧化还原稳定的关键酶,其催化细胞内强氧化离子发生歧化反应生成氧化性较弱的H2O2,H2O2又在POD和CAT的作用下形成H2O,以减少氧化胁迫对植物的毒害作用,因此SOD起着抵御重金属胁迫的先导作用[27]。赵会君等[28]研究显示,Cu、Zn、Mn胁迫处理显著增加了SOD同工酶条带数量和SOD活性,SOD可作为植物耐受重金属污染的重要生理指标。本研究表明,4种供试植物的SOD、POD和CAT活性均随铜镍胁迫浓度增加呈先升后降趋势,其中POD和CAT活性峰值变化相同;而SOD活性峰值有所不同,表现为早熟禾、紫花苜蓿和鹰嘴紫云英SOD活性先于黑麦草达到峰值,说明4种植物的抗氧化系统产生适应性诱导反应的时间节点不同,有较好耐铜镍胁迫能力植物的SOD活性增加时间早于耐受能力差的植物[29]。
土壤重金属污染的植物修复是指富累积植物将金属污染物吸收于自身体内,通过对其不断地刈割和集中处理,将重金属带出土壤的过程[30]。重金属胁迫下,植物各器官对重金属元素的吸收能力有所不同,周婷等[31]研究表明,芹菜对镍的富集能力表现为根系>地上部植株,李媛等[32]研究表明,Cu在小麦各器官中的质量分数表现为根须>麦叶>麦壳>籽粒>茎杆,Ni表现为根须>麦叶>麦壳>茎杆>籽粒。本研究表明,Cu2+含量在黑麦草、早熟禾体内表现为根>茎>叶,在紫花苜蓿、鹰嘴紫云英体内表现为叶>茎>根;Ni2+含量在黑麦草、早熟禾体内分别表现为茎>叶>根、茎>根>叶,在紫花苜蓿、鹰嘴紫云英体内表现为叶>茎>根,可见紫花苜蓿、鹰嘴紫云英对铜镍具有较好的转移能力。
4 结 论
植物耐铜镍性与其形态特征和生理指标密切相关。回归分析表明,地下生物量、可溶性糖、可溶性蛋白、脯氨酸、超氧化物歧化酶和Cu2+对4种供试植物耐铜镍性影响较大;隶属函数综合评价发现,4种植物的耐铜镍性强弱表现为:紫花苜蓿>鹰嘴紫云英>草地早熟禾>多年生黑麦草。本结果可为耐铜镍或超富集植物的筛选及铜镍污染土壤的生物修复提供材料并进行推广和应用。
猜你喜欢
实用手外科杂志(2022年2期)2022-08-31
中国民间疗法(2021年13期)2021-08-30
中学时代(2018年6期)2018-11-20
海峡姐妹(2018年7期)2018-07-27
电子测试(2017年12期)2017-12-18
自然资源情报(2017年5期)2017-11-26
作文与考试·小学高年级版(2016年14期)2016-09-10
中华骨与关节外科杂志(2016年5期)2016-05-17
实用手外科杂志(2015年4期)2015-08-27
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06
